
Download
ORIGINAL ARTICLE
Biotransformation of sesaminol triglycoside by intestinal microflora of swine supplemented with probiotic or antibiotic diet
Kuo-Ching Jan1*, Tzu-Yi Wang2, Lucy Sun Hwang2, Mohsen Gavahian1*
1Department of Food Science, National Pingtung University of Science and Technology, Neipu, Pingtung 91201, Taiwan, Republic of China;
2Graduate Institute of Food Science and Technology, National Taiwan University, Taipei, Taiwan, Republic of China
Abstract
Lignans, found throughout the plant kingdom, are complex diphenolic chemicals that act as phytoestrogens. The main lignan component in sesame flour is sesaminol triglucoside (STG). In vivo, STG is converted into antioxidant sesaminol, mammalian lignans enterodiol, and enterolactone. Thus, use of antibiotics may impact the conversion of lignans to enterolactone. The present study investigated the metabolism of STG by microorganisms in swine-fed diets supplemented with either probiotics or antibiotics. It was observed that microorganisms from swine-fed probiotics helped with hydrolyzing the glucose unit of STG and generated metabolite 2-hydroxymethyl sesaminol- tetrahydrofuran (ST-2) as compared to antibiotic diet. It was also observed that bacteria that converted sesaminol to ST-2 were oxygen-sensitive and it was hypothesized that these were anaerobes. While Lactobacillus and Bifidobacterium were more prevalent in probiotic-fed swine feces, both groups of swine had a healthy mix of helpful and detrimental bacteria. All selective investigated microorganisms (Lactobacillus, Bifidobacterium, and Clostridium perfringens) could hydrolyze the glucose unit of STG; however, lactic acid bacteria contributed least. In conclusion, the bacteria found in the feces of swine-fed probiotics had a greater ability to digest STG. It can be concluded that the biotransformation of STG could be attributable to anaerobes, but Lactobacillus and Bifidobacterium could balance intestinal flora.
Key words: sesame flour, sesaminol triglucoside, biotransformation, gut microbiota, probiotics
*Corresponding Authors: Kuo-Ching Jan and Mohsen Gavahian, National Pingtung University of Science and Technology, Pingtung 912, Taiwan, Republic of China. Emails: [email protected] and [email protected]
Received: 7 December 2021; Accepted: 25 February 2022; Published: 10 June 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Sesame (Sesamum indicum L.), a traditional health food in Asian nations, has been recognized for long as a significant oilseed across the world. Sesame lignans, such as sesamin, sesaminol, and sesamolin, with bioactive properties are significant components of sesame oil and share a methylenedioxyphenyl group as well as various in vivo functions. (Kamal-Eldin et al., 2011). Although sesame oil meal contains a high concentration of linoleic acid, both selenium and vitamin E protect unsaturated fatty acids from oxidation in animal milk fatty acids (Mitsiopoulou et al., 2021). The major lignans (sesamin and sesamolin) are benzodioxol–substituted furofurans. Transformation products found in processed goods include sesamol, sesaminol, its epimers, and episesamin (Andargie et al., 2021). Lignans are important plants formed by the coupling of two methylenedioxyphenyl with extraordinary biological properties that help in plant defense and provide health advantages to humans, including modulation of fatty acid metabolism, inhibition of cholesterol absorption, and biosynthesis, antioxidant, and vitamin E-sparing effects, hypotensive effect, improvement of liver function in relation to alcohol metabolism, and anti-aging effect (Chhillar et al., 2021). However, the maximum content found in sesame is that of sesaminol triglucoside (STG), which is transformed into enterolignans with higher physiological activity after metabolism by microorganisms in the intestines of organisms.
Many investigations have revealed that these phytoestrogens can be converted by gut bacteria into compounds with altered bioactivity via new enzymatic processes. Recent research has also suggested that these metabolites may influence the host gut ecology, gene expression, metabolism, and immune system. Isolating gut microbes capable of biotransforming phytoestrogens and characterizing the novel enzymatic reactions involved are thus essential for understanding the mechanisms of beneficial effects brought by gut microbiota and their metabolism on phytoestrogens as well as providing theoretical knowledge for the development of functional probiotics (Hameed et al., 2020). Some lignans, known as ‘phytoestrogen’ lignans, are transformed by the human gut microbiota into the mammalian lignans (enterodiol and enterolactone) upon intake. These mammalian lignans having chemo-preventive effects against different malignancies (such as breast, colon, and prostate cancers) and/or cardiovascular diseases have been widely characterized and studied (Hano et al., 2021; Rietjens et al., 2017).
Bacteria in the gut perform essential metabolic and physiological functions and have a significant influence on the nutritional and health status of the host. The metabolic activity of the intestinal microflora aids in digestion and absorption of food components as well as transformation of various microorganisms (Gavahian et al., 2019). Composition of the gut microflora population is influenced by both endogenous and external causes. Food is believed to be the most significant driver of gut bacterial diversity among these factors, which may impact the host’s functional relationships with the bacteria (Ley et al., 2008). Dietary components are also important in basic nutrition, resulting in the creation of functional food ideas and nutrition. Hydrolysis of glucosides, demethylation of methoxy groups, oxidation of dibenzylbutanediol to dibenzyl butyrolactone, and reductive cleavage of furanfuran rings contribute to the alteration of STG in the intestinal microflora (Jan et al., 2010). The modification of STG by intestinal bacteria would result in the synthesis of many metabolites, composition of which is greatly regulated by the makeup of intestinal microbiota (Jan et al., 2009a). Furthermore, depending on their chemical structure and quantity, STG and its metabolites can function as activators or inhibitors of bacterial growth in the gut, changing the makeup of intestinal microflora. STG is a big, highly water-soluble molecule that historically has been thought as less likely to be absorbed in the gastrointestinal system. Previous research has demonstrated that only a portion of dietary STG is absorbed, processed, and subsequently transferred to other organs (Jan et al., 2009b, 2011). STG and its metabolites, on the other hand, remain in the gut, suggesting that they may play an important role in maintaining composition and health of intestinal microbiota. Previous research has found that some gut bacteria, such as Bacteroides, Clostridium, Bacillus, and Lactobacillus, are responsible for the metabolism of dietary lignans (Clavel et al., 2006). More than one species is thought to be capable of converting plant lignans in vivo (Woting et al., 2010). To the best of our knowledge, the majority of STG research has been on its metabolism, distribution, and excretion in mice as well as transformation by gut microorganisms (Jan et al., 2009b, 2010; Liao et al., 2010). In this sense, incorporation of probiotics in food and feed attracted the attention of researchers, because such an approach can inhibit food intoxification, improve organoleptical properties, and enhance nutritional quality (Alizadeh et al., 2020; Mokhtarian et al., 2020; Wang et al., 2021). The intestine is home to a diverse microbial community organized around a web of metabolic interconnections. The gut microbiota is now recognized as critical for normal human growth and function, particularly for the priming and maturation of adaptive immune system. Use of antibiotic has been linked with reduction in species diversity, altered metabolic activity, and selection of antibiotic-resistant microbes in the gut microbiota (Ramirez et al., 2020). Moreover, some antibiotics can also act positively on gut microbiota, providing the so-called ‘eubiotic’ effect, by increasing abundance of beneficial bacteria (Ianiro et al., 2016). Increase in the use of antibiotics in recent years suggests that these problems are likely to become more acute or more prevalent in the future.
There is little information on STG in vitro fermentation and its impact on gut flora. For example, Zhu et al. (2013) found that STG substantially increased the numbers of Lactobacillus—Enterococcus and Bifidobacterium—in in vitro fermentation using human fecal microbiota, but it did not promote the bacterial growth of Eubacterium rectale—Clostridium coccoides, Clostridium histolyticum, or Bacteroides—Prevotella. Similarly, Jan et al. (2009a) reported that STG is converted to a catechol moiety by gut bacteria. ST-2 and mammalian lignans were identified as the main STG metabolites. However, no report in the scientific database has assessed lactic acid bacteria or the effects of anaerobic bacteria on sesame lignans and mammalian lignans. As a result, we investigated the impact of dietary intervention on gut microbiota. We discovered that the bioavailability of probiotics and antibiotics in meals influences gut microbiota. The relationship between microbiota and nutritional makeup of food, as well as implications of these interactions on swine health, is the subject of this research. The primary goal of this study was to assess the effects of sesame STG on fermentation by swine fecal bacteria (probiotic diet vs. antibiotic diet). In vitro fermentation was then used to assess the impact of STG on gut microbiota and enterolignan synthesis. This study aims to analyze: (1) effect of dietary antibiotics and lactic acid bacteria on intestinal flora and (2) effect of dietary antibiotics and lactic acid bacteria on the conversion of STG to sesaminol and ST-2.
Material and Methods
Materials and Reagents
XAD–2 gel was purchased from Aldrich (Milwaukee, WI, USA). Triple deionized water (Millipore, Bedford, MA, USA) was used for all preparations. Liquid chromatographic grade solvents were obtained from Mallinckrodt Baker (Phillipsburg, NJ, USA). The rest of the used chemicals were of analytical grade. In order to incubate and identify the species, Rogosa Agar, MRS Broth, Tryptose-sulfite-D-cycloserine agar, and reinforced clostridial media (Darmstadt, Germany) were employed. ChromaDex acquired enterolactone and enterodiol with a purity of 95% (Santa Ana, CA). Sesaminol, sesaminol triglucoside, and the major metabolite 2-episesaminol and R,R–hydroxymethyl sesaminol-tetrahydrofuran were collected and purified in the manner described previously (Jan et al., 2009a).
Animals and treatments
Pigs (Landrace, Yorkshire, and Duroc [LYD] cross pigs) and treatments protocol was authorized, and its implementation was supervised by the Animal Welfare Officer. Crossbred female pigs, 6-month old and weighing 115 ± 4 kg (means ± standard error of mean [SEM]) were kept in separate boxes with tiled floors and walls without litter. Three pigs (n = 3) were brought up, each on probiotic and antibiotic diets. All animals were in good condition and were neither given any medicine for 2 weeks before the research nor throughout it. The component makeup of probiotic (1.5 × 1010 colony- forming units [CFU]/g, including 50% Lactobacillus ATCC4356/ATCC7469/ATCC11842/ATCC1491 and 50% Bifidobacterium ATCC15700/ATCC15697) and antibiotic diets provided to animals is provided in Table 1. Neither diet contained any measurable levels of STG chemicals examined in this investigation.
Table 1. Ingredient composition of diets fed to pigs.
| Ingredient (g/100 g diet) | Blank diet |
Probiotic diet | Antibiotic diet |
|---|---|---|---|
| Corn | 75 | 75 | 75 |
| Soybean meal | 15 | 15 | 15 |
| Probiotics | 4 | 4 | 4 |
| Phosphate | 0.6 | 0.6 | 0.6 |
| CaCO3 | 1.2 | 1.2 | 1.2 |
| NaCl | 0.5 | 0.5 | 0.5 |
| Mineral | 0.1 | 0.1 | 0.1 |
| Choline | 0.1 | 0.1 | 0.1 |
| Probiotics | 0.0 | 2.4 × 10–3 | 0.0 |
| Nosiheptide | 0.0 | 0.0 | 25 × 10–6 |
*Probiotics contained 50% Lactobacillusand 50% Bifidobacterium.
In Vitro Fermentation System
In the current in vitro fermentation investigation, the feces were quickly suspended in a sterile anaerobic salt medium under a continuous flow of CO2 gas, yielding final fecal slurry of 200 g of feces/L, as described by Jensen et al. (1995). The filter-sterilized cysteine was added to the cooled anaerobic salt medium after the sterile anaerobic salt medium was autoclaved and chilled under high-purity nitrogen. Fresh feces were taken directly from each pig, weighed, and promptly put into a weak solution of peptone saline to determine bacteria in feces. The slurry was magnetically agitated and kept anoxic by sparging with high-purity nitrogen gas. At 0, 3, 6, 9, 20, 31, 51, 72, 120, and 168 h, 10 mL of samples (14 mg/mL sesaminol triglucoside) were aseptically taken from the fermenter for STG metabolite analyses. All samples were kept at 20°C until they were analyzed. In order to collect the corresponding fermenting microbial flora, fresh feces were obtained straight from the rectum of pigs fed with probiotic and antibiotic diets. Bifidobacterium spp. were counted using Bifidobacteria iodoacetate medium agar (Munoa and Pares, 1988), while Clostridium perfringens were counted using tryptose-sulfite-D-cycloserine agar (Oxoid, Nepean, ON, Canada) (Harmon et al., 1971). Bifidobacteria iodoacetate medium and tryptose-sulfite-D-cycloserine agar were grown anaerobically for 72 h at 37°C in an environment of 5% CO2 and 7% H2 in N2. The findings were presented as log CFU/g feces.
For analyses, 0.5 g of the inner part of a fecal sample (to retrieve anaerobic material) was mixed considerably with 15 mL of anaerobic solution, followed by serial dilutions to acquire different concentrations (10–1–10–8 ). Starting from the lowest concentration, 50 µL of solution was inoculated on different media using the spread plate method. For incubating Clostridium perfringens, 1 mL of solutions with suitable concentrations, determined by the result of a pre-experiment, was mixed considerably with TSC agar medium using the pour plate method, followed by mixing with the regular TSC medium.
The plate was put in an anaerobic room once the liquid media was solidified. Plates containing 30–300 colonies were included in the colony count. The number of bacteria was expressed as log CFU/g of feces moist weight. The following calculation equations were based on the US Food and Drug Administration’s (FDA) Bacteriological Analytical Manual (BAM): Bifidobacterium spp., Lactobacillus spp., Coliform organisms, and total anaerobic organisms (Christensen et al., 1999).
Analysis and characterization of sesaminol triglucoside and its metabolites by LC-MS/MS
Sesaminol metabolites were identified using liquid chromatography with tandem mass spectrometry (LC-MS-MS) with a Thermo high-performance liquid chromatography (HPLC) system equipped with an electrospray ionization ion trap mass spectrometer (Thermo Scientific hybrid quadrupole-Orbitrap mass spectrometers, San Jose, CA, USA). YMC Hydrosphere C18 column (YMC, Tokyo, Japan) was used for separation. The solvents for elution were 100% methanol (solvent A) and 1% acetic acid, pH 6 (solvent B). The elution program of swine feces at a flow rate of 0.3 mL/min was as follows: 0–15 min, linear gradient from 40–70% A; 15–25 min, linear gradient from 70–100% A. Peaks were detected with an ultraviolet/visible light (UV-Vis) detector at 290 nm. The following electrospray ionization (ESI) parameters were used for analyses: spray voltage, 3,500 V for negative (or positive) mode; CE-Inject voltage, 3.8 kV; DE-Inject voltage, 40 V; sheath gas flow rate, 10 au; capillary temperature, 320°C; and Z-lens voltage, 230 V.
The collision-activated dissociation of selected precursor ions in the collision cell of Orbitrap mass spectrometer resulted in MS/MS product ions, the mass of which was determined using the instrument’s second analyzer. For each study, selected reaction monitoring was utilized to monitor three transitions: sesaminol [M—H]– m/z 369.10→340.1; hydroxymethyl sesaminol-tetrahydrofuran [M—H]– m/z 359.35→221.11; and sesaminol triglycoside [M+Na]+ m/z 879.25→359.11. The xcalibur data acquisition software, version 4.0, was used to process all LC-MS/MS data. A prior study established and validated analytical techniques for detecting STG and its metabolites (sesaminol and hydroxymethyl sesaminol- tetrahydrofuran) in swine feces (Jan et al., 2011). In the concentration range of 0.5−20.0 g/mL, coefficients of variation of interday and intraday tests were <5%. The lower limits of quantification for sesaminol, hydroxymethyl sesaminol-tetrahydrofuran, enterolactone, and enterodiol were 0.2, 0.3, 0.1, and 0.3 g/mL, while the limits of detection were 0.05, 0.07, 0.03, and 0.04 g/mL respectively. This trial was partially completed at National Pingtung University of Science and Technology.
Method validation
The linearity, accuracy, and precision of the method were tested. Methanol was used to make an internal standard stock solution of hesperetin (1 mg/mL). Standard ST-2, sesaminol, and sesaminol triglucoside solutions of 0.02–25.0 lg/mL were produced by successive dilutions from stock solution to test linearity. A linear least-square regression equation generated from the peak area was used to create standard curve. The interday and intraday variability of the technique was tested at seven concentrations on the same day (six replications) as well as on six subsequent days. The precision evaluation by relative standard deviation (RSD) was calculated from the observed concentration (Cobs) as follows: %RSD = [SD/ (Cobs)] × 100. The accuracy (bias%) was evaluated by calculating nominal concentration (Cnom) and mean value of the observed concentration by the following equation: bias (%) = [(Cobs – Cnom)/Cnom] × 100.
Statistical analysis
All samples (during in vitro fermentation period) were extracted in triplicate. Sesaminol triglycoside and its metabolites were expressed in μmol/mL feces. Data were analyzed by ANOVA analysis of variance, and differences were considered statistically significant at P < 0.05.
Results and Discussion
Determination of sesaminol triglucoside and its metabolites
Sesame seeds contain 224–1148 mg/100 g lignans, depicting that it is a good source of lignan comparable to flax-seed. STG consumption per capita is estimated as 6–22 mg/day (Tufail et al., 2020). As a result, although being a small component of daily diet, STG makes a significant contribution to the preservation of human intestinal microbiota and production of enterolignans. The LC-MS/ MS was used to determine STG and its metabolites. The chromatograms reveal that STG was primarily present as enterolignans in the feces of pigs fed with the blank diet (standard swine feed without additional probiotics or antibiotics; Table 2). STG, sesaminol, and ST-2 were also discovered. LC-MS/MS was used to characterize structures of enterolignan. A complete scan in negative ion mode (scan range from 50 to 1,000 m/z) was utilized to identify analyte for analytical condition of sesaminol triglucoside. Table 2 displays parent ion and fragment ions of sesaminol triglucoside and its metabolites (sesaminol and ST-2) in swine feces. An analytical technique was devised to examine the metabolism of enterolignans. Owing to intricacies of enterolignans’ LC-MS/MS profile (Figure 1), a more sensitive selected reaction monitoring (SRM) mass spectrometry MS technique was utilized to conduct their studies. Intra- and inter-assay (Tables 3, 4, and 5) accuracy of STG, sesaminol, and ST-2 levels fell well within predefined limits of acceptability. All bias (%) and RSD values were within ±15% limits. The in vivo accuracy of STG and its metabolites indicates that this method is sufficiently sensitive to measure STG and its metabolites in swine feces for this study.
Table 2. LC-MS-MS detection of sesaminol triglucoside and its metabolites in swine feces.
| Metabolite | LC-MS-MS parent ion selection | Fragment ions, negatively or positively charged |
|---|---|---|
| Sesaminol triglucoside | 879.25 [M+ Na]+ | 717.20, 509.15, 347.09, 331.10 |
| Sesaminol | 369.10 [M–H]– | 340.10, 339.09, 311.09, 219.07, 177.02 |
| ST-2 | 359.35 [M–H]– | 311.15, 221.11, 203.24, 191.31 |
Table 3. Intraday and interday precision and accuracy of the LC-MS-MS method for determination of total form of STG in swine feces.
| Nominal conc. (μg/mL) |
Intraday | Interday | ||||
|---|---|---|---|---|---|---|
| Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) |
Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) | |
| 25 | 25.3 ± 0.01 | 0.048 | 1.059 | 25.2 ± 0.15 | 0.589 | 0.641 |
| 12.5 | 11.9 ± 0.13 | 1.114 | –4.732 | 12.2 ± 0.07 | 0.569 | –2.75 |
| 6.25 | 6.01 ± 0.04 | 0.592 | –3.273 | 6.16 ± 0.10 | 1.647 | –1.402 |
| 3.13 | 3.12 ± 0.02 | 0.601 | –0.212 | 3.14 ± 0.04 | 1.425 | 0.471 |
| 1.56 | 1.58 ± 0.03 | 1.678 | 0.804 | 1.60 ± 0.04 | 2.655 | 2.701 |
| 0.78 | 0.80 ± 0.002 | 0.277 | 2.251 | 0.77 ± 0.02 | 2.643 | –1.725 |
| 0.39 | 0.39 ± 0.0003 | 0.087 | –1.166 | 0.40 ± 0.02 | 4.470 | 3.669 |
| 0.20 | 0.2 ± 0.0002 | 0.101 | 1.480 | 0.20 ± 0.002 | 0.787 | 1.992 |
Data are expressed as mean ± SD (n = 6).
Table 4. Intraday and interday precision and accuracy of the LC-MS-MS method for determination of total form of sesaminol in swine feces.
| Nominal conc. (μg/mL) |
Intraday | Interday | ||||
|---|---|---|---|---|---|---|
| Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) |
Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) | |
| 25 | 24.8 ± 0.10 | 0.106 | 0.429 | 24.2 ± 0.23 | 0.965 | –3.142 |
| 12.5 | 12.8 ± 0.04 | 0.339 | 3.122 | 12.6 ± 0.07 | 0.558 | 1.011 |
| 6.25 | 6.34 ± 0.07 | 1.154 | 1.450 | 6.13 ± 0.06 | 0.969 | –1.955 |
| 3.13 | 3.02 ± 0.06 | 1.833 | –3.333 | 3.11 ± 0.03 | 0.896 | –0.454 |
| 1.56 | 1.58 ± 0.01 | 0.716 | 0.807 | 1.60 ± 0.02 | 1.431 | 2.210 |
| 0.78 | 0.75 ± 0.02 | 2.001 | –3.722 | 0.80 ± 0.02 | 2.958 | 2.391 |
| 0.39 | 0.38 ± 0.004 | 1.107 | –2.820 | 0.39 ± 0.004 | 1.203 | 0.415 |
| 0.20 | 0.2 ± 0.004 | 2.231 | –0.168 | 0.19 ± 0.004 | 2.075 | –4.242 |
Data are expressed as mean ± SD (n = 6).
Table 5. Intraday and interday precision and accuracy of the LC-MS-MS method for determination of total form of ST-2 in swine feces.
| Nominal conc. (μg/mL) |
Intraday | Interday | ||||
|---|---|---|---|---|---|---|
| Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) |
Observed conc. (μg/mL) |
Precision (% CV) |
Accuracy (% bias) | |
| 25 | 25.0 ± 0.10 | 0.401 | 0.029 | 25.0 ± 0.08 | 0.334 | 0.004 |
| 12.5 | 12.5 ± 0.02 | 0.121 | –0.067 | 12.5 ± 0.10 | 0.803 | 0.081 |
| 6.25 | 6.25 ± 0.004 | 0.056 | 0.059 | 6.23 ± 0.21 | 0.346 | –0.253 |
| 3.13 | 3.13 ± 0.004 | 0.129 | 0.042 | 3.11 ± 0.02 | 0.506 | –0.289 |
| 1.56 | 1.59 ± 0.05 | 2.854 | 1.723 | 1.57 ± 0.01 | 0.911 | 0.768 |
| 0.78 | 0.80 ± 0.009 | 1.067 | 2.872 | 0.81 ± 0.003 | 0.426 | 3.873 |
| 0.39 | 0.40 ± 0.004 | 0.992 | 3.595 | 0.37 ± 0.005 | 1.386 | –4.182 |
| 0.20 | 0.19 ± 0.001 | 0.669 | –2.935 | 0.20 ± 0.001 | 0.460 | 2.009 |
Data are expressed as mean ± SD (n = 6).
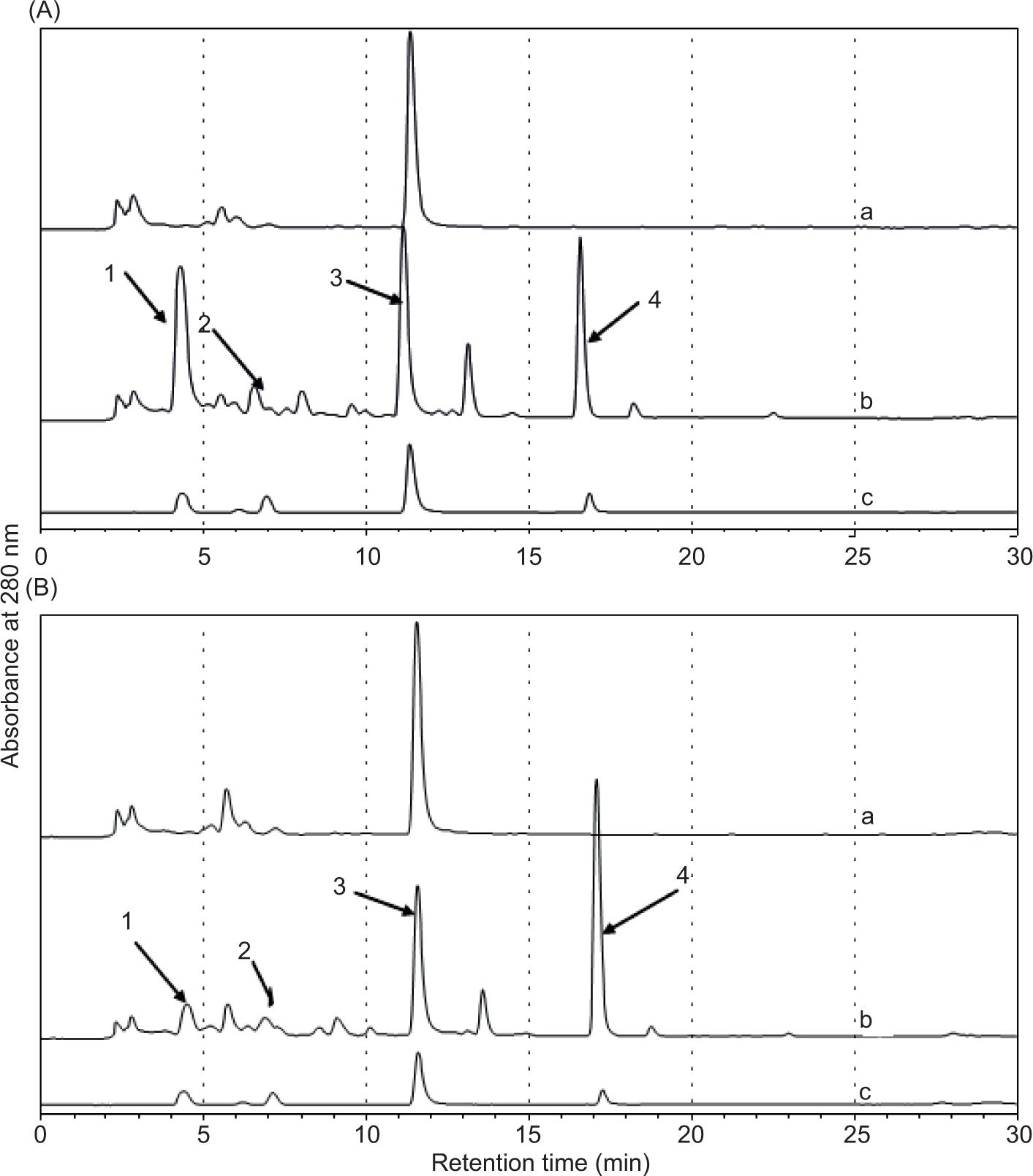
Figure 1. HPLC chromatogram of fermented solution from swine feces. (A) Feces of probiotic group and (B) feces of antibiotic group (1: 2-hydroxymethyl sesaminol-tetrahydrofuran (ST2); 2: sesaminol triglycoside (STG); 3: internal standard; 4: sesaminol. a: feces with phosphate buffer solution (PBS); b: feces with STG; c: standard containing ST-2, STG, hesperetin, and sesaminol).
Microbiota may convert lignans from various cereals into enterolignans (enterodiol and enterolactone). Further metabolism of the furan ring, demethylation, and dehydroxylation of intermediate metabolites, similar to STG, might be carried out by intestinal microbiota. As a result, blank of fermentation broths identified STG-metabolized sesaminol and ST-2. Under the same operational circumstances, samples from blank probiotic and antibiotic diets added STG to ferment. The results reveal that sesaminol and ST-2 were found in fecal samples of pigs fed with both probiotic and antibiotic diets (Figure 1). Enterolignans were metabolized probably from a feed mixture of maize and soybean (Pealvo et al., 2005).
Effect of sesaminol triglucoside on swine intestinal microbiota
Table 6 shows changes in bacterial populations that occurred during the in vitro fermentation of STG at 12 h. Lactobacillus and Bifidobacterium counts were substantially higher in probiotic-fed swine feces than in antibiotic diet samples. Lactic acid bacteria counts were higher in probiotic diet samples whereas counts of Clostridium perfringens were higher in antibiotic diet samples, albeit neither difference was statistically significant. These findings suggest that a probiotic diet may enhance the growth of Lactobacillus and Bifidobacterium, compared to antibiotic diet. STG substantially increased the populations of Lactobacillus–Enterococcus and Bifidobacterium during in vitro fermentation with fecal microbiota, but it did not promote the development of Clostridium histolyticum, Eubacterium rectale–Clostridium coccoides, or Bacteroides–Prevotella (Zhu et al., 2013).
Table 6. Quantification of microorganisms from swine feces cultured with STG at 12 h.
| Microorganisms | Log (CFU/g feces) | |
|---|---|---|
| Antibiotic-fed swine |
Probiotic-fed swine | |
| Lactobacillus | 8.96 ± 0.13 | 9.37 ± 0.40* |
| Bifidobacterium | 8.67 ± 0.15 | 9.36 ± 0.41** |
| Lactic acid bacteria | 9.21 ± 0.16 | 9.52 ± 0.42 |
| Clostridium perfringens | 6.58 ± 0.20 | 6.10 ± 0.69 |
*P < 0.05, **P < 0.01.
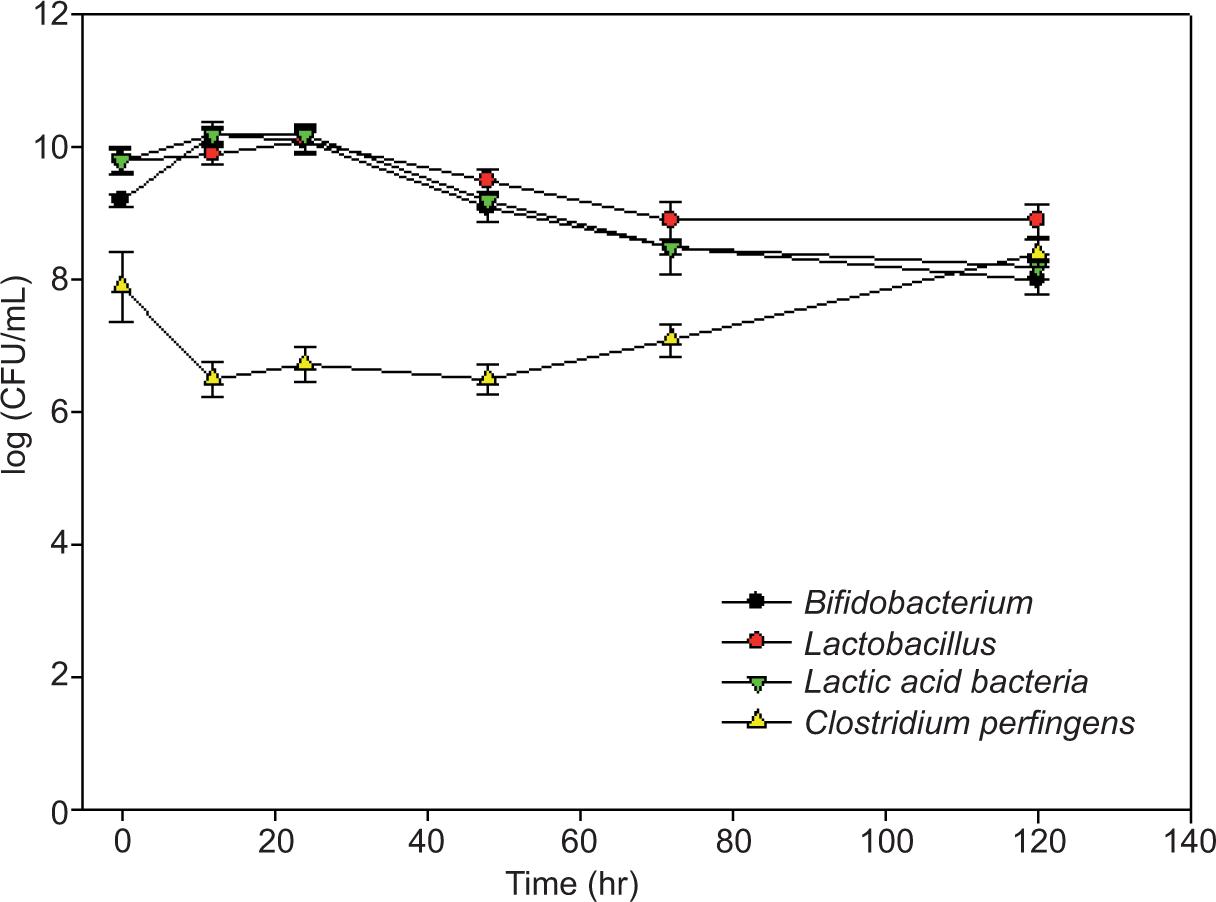
Figure 2 depicts changes in bacterial populations that occurred during STG fermentation in probiotic diet samples. Although Lactobacillus and Bifidobacterium populations were substantially greater than Clostridium perfringens’ populations when fermentation commenced, these dropped somewhat over time. Lactic acid bacteria grew slightly after 3, 12, and 24 h of fermentation but dramatically reduced between 72 and 168 h (Figure 2). After a brief decline at 12 h, the Clostridium perfringens’ population rebounded and was considerably higher after fermentation period than at the beginning. The numbers converged with similar amounts of all four species of bacteria toward the conclusion of 120-h fermentation.
Figure 2. Quantification of microorganisms in the feces of swine fed with probiotic diet over a time course of STG fermentation.
Effect of sesaminol triglucoside on metabolites production
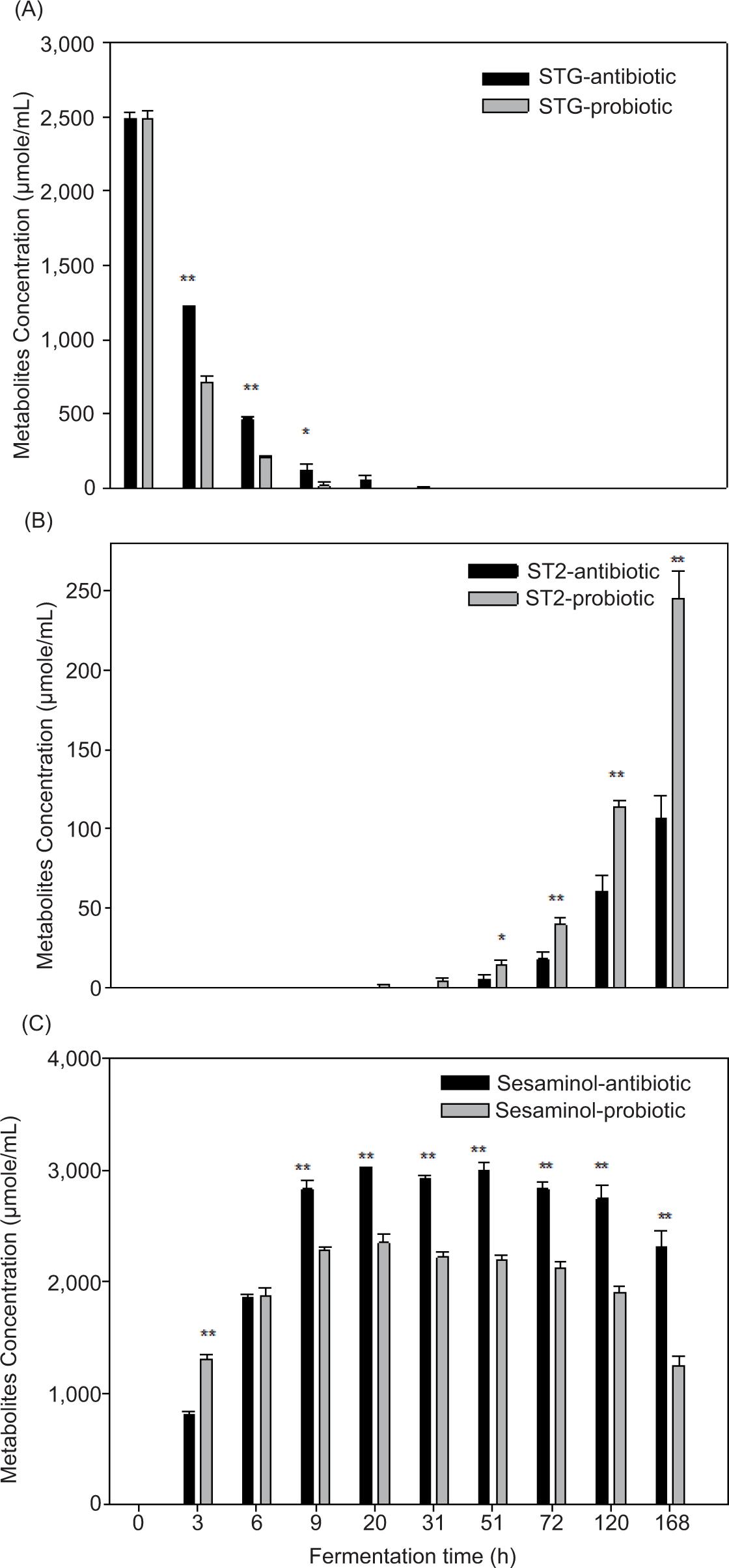
Sesaminol is a crucial molecule generated during the fermentation process through deglycosylation route. During fermentation, STG is quickly transformed into sesaminol by microorganisms in the swine intestinal system. High numbers of Lactobacillus and Bifidobacterium may have increased sesaminol in the early stages of fermentation. Following this, a decrease in sesaminol concentration was observed between 31 h and 168 h. This correlates to an increase in ST-2 concentration as well as an increase in Clostridium perfringens’ population (Figure 2), indicating that this bacterium could play a role in the dynamics of these metabolites. After 6 h of fermentation, we discovered that just a little amount of STG (0.6% probiotic diet and 4.4% antibiotic diet) remained in the medium (Figure 3). STG vanished between 31 h and 51 h of fermentation. The concentration of metabolite sesaminol rose substantially in both treatments. The amount of sesaminol was higher than that of STG after 6 h of fermentation, and the maximum production of sesaminol was recorded between 9 h and 72 h of fermentation. While synthesis of sesaminol rose considerably, molar proportions were seen to grow initially and reach maximum at 9–51 h but declined progressively after that. Sesaminol content was higher in antibiotic-fed swine samples than in probiotic-fed pigs at 9 and 20 h of fermentation. The increased synthesis of sesaminol is related to the metabolic activities of Bifidobacterium and Lactobacillus. Presence of deglycosylation, as indicated by deglycosylated sesaminol discovered here, is consistent with the metabolic activity of potentially helpful Bifidobacterium and Lactobacillus bacteria. During the same fermentation period, the samples from antibiotic diet-fed swine generated more sesaminol than the samples from pro-biotic diet-fed swine, as shown in Figure 3. However, for both treatments, the molar percentage of STG fell significantly during the first 20 h of fermentation; however, the molar proportions of sesaminol and ST-2 rose significantly (Figure 3). ST-2 was identified for the first time after 6 h of fermentation and grew throughout rest of the period. Levels of ST-2 were substantially higher in probiotic samples than in antibiotic samples. Synthesis of ST-2 at a later stage suggests that it may be connected to the transformation of other bacterial metabolites produced during fermentation or changes in bacterial populations. Sesaminol levels were high in both groups after 3 h. Although the amount of sesaminol in antibiotic diet was higher than in probiotic diet at 3 h and was equal at 6 h, levels in antibiotic diet were considerably higher throughout the fermentation.
Figure 3. Biotransformation of sesaminol triglycoside (STG) to its metabolites in feces of probiotic- and antibiotic-fed swine. (A) STG, (B) sesaminol, and (C) ST-2 (*P < 0.05, **P < 0.01).
The findings demonstrated that the biotransformation of STG to mammalian lignans by intestinal microbiota involved the following four types of reactions: lignan glucoside hydrolysis, methoxy group demethylation, oxidation of dibenzylbutanediol to dibenzylbutyrolac-tone, and reductive cleavage of furofuran rings (Jan et al., 2010; Landete, 2012; Suja et al., 2005; Zhu et al., 2013). When it comes to describing the health advantages of dietary components, it is crucial to distinguish between content and bioavailability (i.e., the fraction that is eventually absorbed and available for physiological functions or deposition), because greater the amount of chemical available, greater the quantity engaged in biological processes (Rodriguez-Concepcion et al., 2018). It is crucial to note at this point that absorption of lipophilic chemicals (carotenoids) requires their release from the matrix, and because they cannot dissolve in aqueous environments, they must be integrated into mixed micelles. These, which require the formation of lipids and bile components, can be taken up by the apical surface of enterocytes, from which lipophilic substances can be finally absorbed, that is, integrated into circulation (Meléndez-Martínez et al., 2017). Microbiota metabolites of sesame lignans are more absorbed in the gut, and enterohepatic circulation guarantees that duration of stay of metabolites in plasma is longer than that of their parent components. Finally, metabolites are eliminated in urine (Jan et al., 2011). Previous research investigated the variety and occurrence of lignan-transforming gut bacteria using in vitro culture-based tests. Several publications have proved the production of enterolignans by rat intestinal microbiota (Hung et al., 2016) and human fecal microbiota (Zhu et al., 2013). We discovered that feces of probiotic-fed pigs had a lower pH than that of antibiotic-fed swine, implying another health advantage of probiotic intake. In the current investigation, we discovered that STG content was higher in fecal samples of pigs offered antibiotic diet than those administered probiotic diet at 3 and 6 h of fermentation. Furthermore, sesaminol concentrations increased considerably in both groups toward the end of fermentation. STG was critical in stimulating the development of lactic acid bacteria during the early stages of fermentation. However, the amount of ST2 production was very low, which further affected the amount of enterodiol production. However, sesaminol was also active physiologically in swine intestinal micro-biota. These findings imply that intestinal microbiota in humans, mice, and pigs are very different, and that swine intestinal bacteria are unable to convert plant lignin to mammalian lignans. Perhaps this is the best explanation for the above-mentioned changes in swine intestinal bacteria and sesame metabolites that occur during fermentation. STG metabolism provided more evidence for the role of swine gut bacteria in maintenance of organism’s health.
Conclusion
The in vitro transformation of sesaminol triglucoside to enterolignan was demonstrated by utilizing a specified mixed culture of bacteria. The metabolism by micro-organisms is a key step to metabolize lignans, and the intake of antibiotics affects the conversions of lignans to enterolactone. This study has established that probiotic swine microorganisms can readily hydrolize the glucose unit of STG and get more ST-2 metabolite than the antibiotic group. The study also discovered that micro-organisms that converted sesaminol to ST-2 were sensitive to oxygen, and might be absolute anaerobes. On the other hand, probiotic swine feces had more Lactobacilus and Bifidobacterium and both groups had a great balance of good and bad microorganisms. All selective microorganisms, Lactobacillus, Bifidobacterium, lactic acid bacteria, and Clostridium, hydrolized the glucose unit of STG, but the contribution of lactic acid bacteria was less. In conclusion, changes in diet affect intestinal microorganisms, and therefore alter the metabolism of STG. Microorganisms present in antibiotic swine feces have stronger capacity to metabolize STG. Biotransformation of STG could be due to presence of other microorganisms in feces and not because of Lactobacillus and Bifidobacterium. Diet significantly affects intestinal environment, influencing the makeup of intestinal microbiota and perhaps its functional effects on mammalian lignans. These findings offered more evidence for the relevance of animal gut bacteria in the bioactivity of STG and its metabolites in maintenance of animal health.
Ethics statement/Ethical approval
Consideration of ethical and welfare aspects by the Animal Care & Ethics Committee (ACEC) was not required for developing the current study. Animals were subjected to standard production practices during both growing and finishing stages in compliance with the EU Council 2008/120 directive regarding the minimum standards for the protection of pigs, which was transposed into national legislation for the care and handling of animals by RD 1392/2012, and to pigs reared on extensive systems by RD 1221/2009, and with slaughter conditions. No additional measures were required.
Declaration of interest
The authors declared no conflicts of interest.
Data availability statement
Data are available on request from authors.
Authors contribution
Kuo-Ching Jan: conceptualization, methodology, software validation, formal analysis, investigation, resources, data curation, writing of original draft, writing of review & editing, visualization, supervision, project administration, funding acquisition. Mohsen Gavahian: writing— review & editing.
REFERENCES
Alizadeh, A.M., Hashempour-Baltork, F., Alizadeh-Sani, M., Maleki, M., Azizi-Lalabad, M. and Khosravi-Darani, K., 2020. Inhibition of Clostridium (C.) botulinum and its toxins by pro-biotic bacteria and their metabolites: an update review. Quality Assurance and Safety of Crops & Foods 12(SP1): 59–68. 10.15586/qas.v12iSP1.823
Andargie, M., Vinas, M., Rathgeb, A., Möller, E. and Karlovsky, P., 2021. Lignans of sesame (Sesamum indicum L.): a comprehensive review. Molecules 26(4): 883. 10.3390/molecules26040883
Chhillar, H., Chopra, P. and Ashfaq, M.A., 2021. Lignans from linseed (Linum usitatissimum L.) and its allied species: retrospect, introspect and prospect. Critical Reviews in Food Science and Nutrition 61(16): 2719–2741. 10.1080/10408398.2020.1784840
Christensen, D.N., Bach Knudsen, K.E., Wolstrup, J. and Jensen, B.B., 1999. Integration of ileum cannulated pigs and in vitro fermentation to quantify the effect of diet composition on the amount of short-chain fatty acids available from fermentation in the large intestine. Journal of the Science of Food and Agriculture 79: 755–762. 10.1002/(SICI)1097-0010(199904)79:5<755::AID-JSFA248>3.0.CO;2-2
Clavel, T., Borrmann, D., Braune, A., Doré, J. and Blauta, M., 2006. Occurrence and activity of human intestinal bacteria involved in the conversion of dietary lignans. Anaerobe 12: 140−147. 10.1016/j.anaerobe.2005.11.002
Gavahian, M., Khaneghah, A.M., Lorenzo, J.M., Munekata, P.E., Garcia-Mantrana, I., Collado, M.C. and Barba, F.J., 2019. Health benefits of olive oil and its components: impacts on gut micro-biota antioxidant activities, and prevention of noncommunicable diseases. Trends in Food Science Technology 88: 220–227. 10.1016/j.tifs.2019.03.008
Hameed, A.S.S., Rawat, P.S., Meng, X. and Liu, W., 2020. Biotransformation of dietary phytoestrogens by gut microbes: a review on bi-directional interaction between phytoestrogen metabolism and gut microbiota. Biotechnology Advances 43: 107576. 10.1016/j.biotechadv.2020.107576
Hano, C.F., Dinkova-Kostova, A.T., Davin, L.B., Cort, J.R. and Lewis, N.G., 2021. Lignans: insights into their biosynthesis, metabolic engineering, analytical methods and health benefits. Frontiers in Plant Science 11: 2142. 10.3389/978-2-88966-491-7; 10.3389/fpls.2020.630327
Harmon, S.M., Kautter, D.A. and Peeler, J.T., 1971. Improved medium for enumeration of Clostridium perfringens. Journal of Applied Microbiology 22: 688–692. 10.1128/am.22.4.688-692.1971
Hung W.L., Liao C.D., Lu W.C., Ho C.T., Hwang L.S., 2016. Lignan glycosides from sesame meal exhibit higher oral bioavail-ability and antioxidant activity in rat after nano/submicro-sizing. Journal of Functional Foods 23: 511–522. 10.1016/j.jff.2016.03.008
Ianiro G., Tilg H. and Gasbarrini A., 2016. Antibiotics as deep modulators of gut microbiota: between good and evil. Gut 65(11): 1906–1915. 10.1136/gutjnl-2016-312297
Jan, K.C., Hwang, L.S. and Ho, C.T., 2009a. Biotransformation of sesaminol triglucoside to mammalian lignans by intestinal microbiota. Journal of Agricultural and Food Chemistry 57: 6101−6106. 10.1021/jf901215j
Jan, K.C., Hwang, L.S. and Ho, C.T., 2009b. Tissue distribution and elimination of sesaminol triglucoside and its metabolites in rat. Molecular Nutrition & Food Research 53: 815−825. 10.1002/mnfr.200800380
Jan, K.C., Ku, K.L., Chu, Y.H., Hwang, L.S. and Ho, C.T., 2010. Tissue distribution and elimination of estrogenic and anti-inflammatory catechol metabolites from sesaminol trigluco-side in rats. Journal of Agricultural and Food Chemistry 58: 7693−7700. 10.1021/jf1009632
Jan, K.C., Ku, K.L., Chu, Y.H., Hwang, L.S. and Ho, C.T., 2011. Intestinal distribution and excretion of sesaminol and its tetrahydrofuranoid metabolites in rats. Journal of Agricultural and Food Chemistry 59: 3078−3086. 10.1021/jf105012v
Jensen, M.T., Cox, R.P. and Jensen, B.B., 1995. Microbial production of skatole in the hind gut of pigs fed different diets and its relation to skatole deposition in backfat. Journal of Animal Science, 61: 293–304. 10.1017/S1357729800013837
Kamal-Eldin, A., Moazzami, A. and Washi, S., 2011. Sesame seed lignans: potent physiological modulators and possible ingredients in functional foods & nutraceuticals. Recent Patents on Food, Nutrition & Agriculture 3: 17−29. 10.2174/2212798411103010017
Landete, J.M., 2012. Plant and mammalian lignans: a review of source, intake, metabolism, intestinal bacterial and health. Food Research International 46: 410−424. 10.1016/j.foodres.2011.12.023
Ley, R.E., Lozupone, C.A., Hamady, C.A., Knight, R. and Gordon, J.I., 2008. Worlds within worlds: evolution of the vertebrate gut microbiota. Nature Reviews Microbiology 6: 776−788. 10.1038/nrmicro1978
Liao, C.D., Hung, W.L., Lu, W.C., Jan, K.C., Shih, D.Y.C., Yeh, A.I., Ho, C.T. and Hwang, L.S., 2010. Differential tissue distribution of sesaminol triglucoside and its metabolites in rats fed with lignan glycosides from sesame meal with or without nano/submicrosizing. Journal of Agricultural and Food Chemistry 58: 563−569. 10.1021/jf9028046
Meléndez-Martínez, A.J., Pérez-Gálvez, A., Roca, M., Estévez-Santiago, R., OlmedillaAlonso, B., Mercadante, A.Z., et al., 2017. Biodisponibilidad de carotenoides, factores que la determinan y métodos de estimación. In: Meléndez-Martínez, A.J. (ed.) Carotenoides en agroalimentación Y salud. CYTED: Madrid, Spain; pp. 574–608.
Mitsiopoulou, C., Karaiskou, C., Simoni, M., Righi, F., Pappas, A.C., Sotirakoglou, K. and Tsiplakou, E., 2021. Influence of dietary sesame meal, vitamin E and selenium supplementation on milk production, composition, and fatty acid profile in dairy goats. Livestock Science 244: 104336. 10.1016/j.livsci.2020.104336
Mokhtarian, M., Tavakolipour, H., Bagheri, F., Oliveira, C.A.F., Corassin, C.H. and Khaneghah, A.M., 2020. Aflatoxin B1 in the Iranian pistachio nut and decontamination methods: a systematic review. Quality Assurance and Safety of Crops & Foods 12(4): 15–25. 10.15586/qas.v12i4.784
Munoa, F.J. and Pares, R., 1988. Selective medium for isolation and enumeration of Bifidobacterium spp. Applied and Environmental Microbiology 54: 1715–1718. 10.1128/aem.54.7.1715-1718.1988
Ramirez, J., Guarner, F., Bustos Fernandez, L., Maruy, A., Sdepanian, V.L., Cohen, H., 2020. Antibiotics as major disruptors of gut microbiota. Frontiers in Cellular and Infection Microbiology 24(10): 572912. 10.3389/fcimb.2020.572912
Rietjens, I.M., Louisse, J. and Beekmann, K., 2017. The potential health effects of dietary phytoestrogens. British Journal of Pharmacology 174: 1263–1280. 10.1111/bph.13622
Rodriguez-Concepcion, M., Avalos, J., Bonet, M.L., Boronat, A., Gomez-Gomez, L., Hornero-Mendez, D., 2018. A global perspective on carotenoids: metabolism, biotechnology, and benefits for nutrition and health. Progress in Lipid Research 70: 62–93. 10.1016/j.plipres.2018.04.004
Suja, K.P., Jayalekshmy, A., Arumughan, C., 2005. Antioxidant activity of sesame cake extract. Food Chemistry 91: 213−219. 10.1016/j.foodchem.2003.09.001
Tufail, T., Riaz, M., Arshad, M.U., Gilani, S.A., Ul Ain, H.B., Khursheed, T., Islam, Z., Imran, M., Bashir, S., Zia Shahid, M., Kazmi, S.M.U., Saqib, A., 2020. Functional and nutraceutical scenario of flaxseed and sesame. International Journal of Biological Sciences 17: 173–190.
Wang, J.L., Zhao, F., Cairang, Z.M., Li, X.Y., Kong, J., Zeng, S.Y., et al., 2021. Correlation between the bacterial community and flavour of fermented fish. Quality Assurance and Safety of Crops & Foods 13(3): 82–91. 10.15586/qas.v13i3.908
Woting, A., Clavel, T., Loh, G. and Blaut, M., 2010. Bacterial transformation of dietary lignans in gnotobiotic rats. FEMS Microbiology Ecology 72: 507–514. 10.1111/j.1574-6941.2010.00863.x
Zhu, X., Zhang, X., Sun, Y., Su, D., Sun, Y., Hu, B. and Zeng, X., 2013. Purification and fermentation in vitro of sesaminol triglucoside from sesame cake by human intestinal microbiota. Journal of Agricultural and Food Chemistry 61: 1868–1877. 10.1021/jf304643k