
Download
Y t = Y 0 + Y max − ln e Y 0 + e Y max − e Y 0 e − μ max B t , B t = t + 1 4 ln 1 + e − 4 t − λ 1 + e 4 λ 1 Ln λ = a + b eT + fC + c eT + fC 2 + d eT + fC 3 , 2 Ln μ max = a + b eT + fC + c eT + fC 2 + d eT + fC 3 , 3 R 2 = 1 − ∑ e i 2 ∑ y i − y ′ 2 4 A f = 10 ∑ log 10 predicted/observed n 5 B f = 10 ∑ log 10 predicted/observed n 6 RMSE= ∑ observed values − predicted values 2 n 7
Ln λ = 5.220 − 1.966 × 0.072 T- 0.516 C − 0.717 × 0.072 T − 0.516 C 2 + 0.309 × 0.072 T − 0.516 C 3 , 8 Ln μ max = − 5.905 + 4.415 × 0.053 T- 0.248 C − 0.305 × 0.053 T − 0.248 C 2 + 0.239 × 0.053 T − 0.248 C 3 , 9 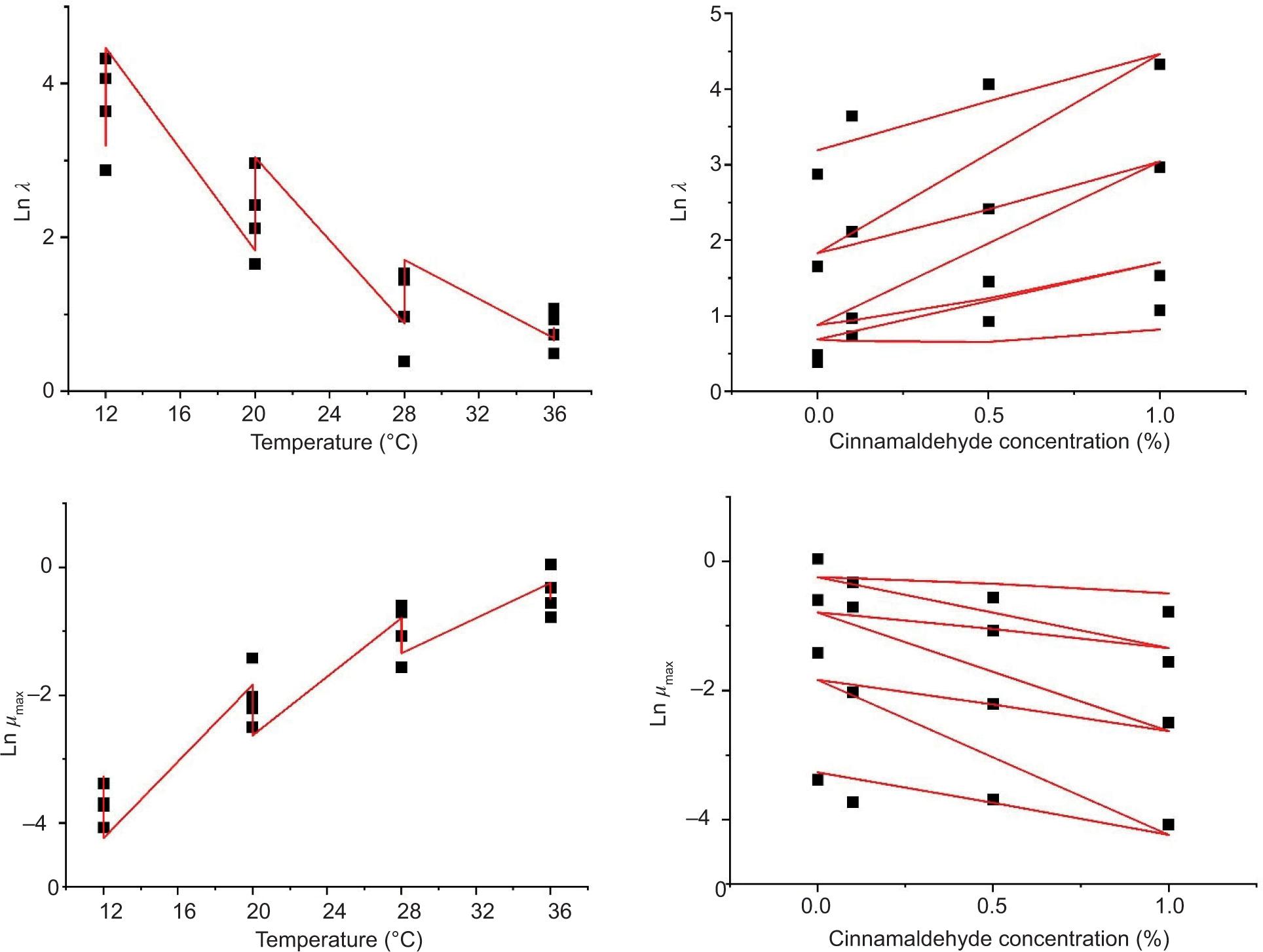
RESEARCH ARTICLE
Effects of cinnamaldehyde on the germination and growth of Bacillus cereus spores in ready-to-eat boiled ground beef
Menghan Zhang, Ruixiang Zhou, Sixu Jin, Yuhang Song, Nuonuo Xu, Yuexia Wang*
College of Life Sciences, Henan Agricultural University, Zhengzhou, China
Abstract
The study quantitatively described the effects of cinnamaldehyde on the germination and growth of Bacillus cereus spores in boiled ready-to-eat ground beef. With the combination of the concentrations of cinnamaldehyde 0, 0.1, 0.5, and 1.0% vol/wt at temperatures 12, 20, 28, and 36°C, the Huang model was successfully used as the primary model to predict the lag time (λ) and maximum growth rate (µmax). Thereafter, the cubic polynomial models were used to estimate the values of Ln λ and Ln µmax considering both the storage temperature and cinnamaldehyde concentration. The models were highly accurate, because they produced acceptable root mean squared error (RMSE) values that were close to 0, while the determination (R2), accuracy factor (Af), and bias factor (Bf) values were all close to 1. As indicated by the fitted models, the supplementary of cinnamaldehyde in samples increased the lag time of B. cereus significantly from 17.7 to 75.8 h at 12°C of storage. Increasing the storage temperature from 12 to 36°C led to only 0.80-fold prolongation of lag time from 1.6 to 2.9 h in the sample containing cinnamaldehyde. However, the µmax value declined most obviously at 20°C, while 66% decrease was determined. According to the results, the cinnamaldehyde can be used as a natural antimicrobial agent in boiled ready-to-eat ground beef in staple food industry by inhibiting the germination and growth of B. cereus. The results also provided regressing models that can be used for effectively designing the storage temperature and cinnamaldehyde concentration for a specific requirement.
Key words: Bacillus cereus, boiled ready-to-eat beef, cinnamaldehyde, predictive microbiology
*Corresponding Author: Yuexia Wang, College of Life Sciences, Henan Agricultural University, Zhengzhou, Henan, China. Email: [email protected]
Received: 18 January 2022; Accepted: 4 April 2022; Published: 20 May 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Bacillus cereus is widely detected in retailed flour (Kindle et al., 2019), rice and rice derivatives (Rodrigo et al., 2021), ultra-high temperature mild (Alonso et al., 2021), pasteurized rice cakes (Park and Yoon, 2019), pasteurized milk, and flour products (Chitov et al., 2008). In China, a recently systematic investigation on the prevalence and characterization of B. cereus in meat and meat products showed 26.37% positive contamination rate, and all isolates presented with multiple antibiotic resistance, virulence genes, and genetic diversity (Kong et al., 2021). B. cereus is ubiquitous in the environment, especially surviving from non-pressurized thermal treatment by forming endospores and commonly implicated in foodborne outbreaks. Therefore, compared to the raw meat, B. cereus is more likely to contaminate or residue in the cooked meat sample, thus can pose a potential public health threat (Tewari et al., 2015).
Ready-to-eat boiled ground beef is a type of meat products that is commonly used to constitute cooked meals along with the staple rice. As the requirement of food safety control, natural antibacterial agents are widely used to inhibit the spore germination or proliferation of foodborne pathogens including B. cereus (Cayemitte et al., 2021; Hou et al., 2021). Cinnamaldehyde is a natural agent that exists in Cinnamomum cassia, which has exhibited a broad-spectrum antibacterial property in animal feeds and human foods (Friedman, 2017). The cinnamaldehyde has been used as an antifungal agent to inhibit Aspergillus niger in bread preservation (Sun et al., 2020). The supplementation of cinnamaldehyde in ground pork facilitated the thermal inactivation of Salmonella (Suo et al., 2017b) and Listeria monocytogenes (Wang et al., 2020). Application of cinnamaldehyde at high pressures can inactivate B. cereus spores in infant formula (Cetin-Karaca and Morgan, 2018). The addition of cinnamaldehyde in carrot broth inhibited the outgrowth from activated spores of B. cereus (Valero and Francés, 2006). In minced beef meat, the treatment of cinnamaldehyde increased the relative sensitivity of B. cereus spores to radiation, thus inhibited the growth of B. cereus during refrigerated storage (Ayari et al., 2012). However, only little information was reported about the effect of cinnamaldehyde on the germination and growth of B. cereus spores in ready-to-eat boiled ground beef.
Due to the possible health risk and the consideration of cost consumption, the antibacterial agents are normally used in food industry. Even so, the usage of the antibacterial agent is still needed to be well-designed to fulfill the least requirement for inhibiting bacterial growth (Doyle and Stephens, 2019). Predictive microbiology provides a useful tool for describing bacterial growth according to individual food environmental factors, such as the storage temperature and the additive concentration (Omidi-Mirzaei et al., 2020; Wemmenhove et al., 2021). Some studies have predictively modeled the growth of B. cereus during the cooling of cooked rice (Hwang and Huang, 2019; Juneja et al., 2019a) and pasta (Juneja et al., 2019b). However, to our knowledge, sparsely primary and secondary models have been developed or suggested to quantitatively describe the effect of cinnamaldehyde on the germination and growth of B. cereus spores in ready-to-eat boiled ground beef during storage at different temperatures.
The objective of this study was to assess the effect of cinnamaldehyde on the germination and growth of B. cereus in boiled ready-to-eat ground beef. Meanwhile, a secondary model was fitted to describe its effect during storage, which would be helpful for designing the additive concentration and storage temperature, based on the statistically described behavior of B. cereus.
Material and Methods
Bacterial strains and spore production
Four strains are used that include B. cereus CICC 23828 and three isolates from meat samples. The B. cereus CICC 23828 was obtained from China Center of Industrial Culture Collection (CICC), and the isolates were gifted by the Quality Detection Center of Henan Provincial Animal Husbandry Bureau. The isolates were identified using both the partial 16S rRNA gene sequencing and the biochemical identification (Kong et al., 2021). Among the used strains, one isolate is psychrotrophic strain as it can grow slowly at 10°C, while others should be mesophilic strains as non-growth was observed at the same condition within 20 h (Guérin et al., 2016). The strains were stored at −80°C with glycerol.
For sporulation, the cryo-preserved cultures of all strains were plated on tryptic soy agar (TSA, Land Bridge Technology Co. Ltd, Beijing, China), and then a single colony was picked and inoculated into 0.1 mL tryptic soy broth (TSB, Land Bridge Technology Co. Ltd, Beijing, China). The cells were grown at 37°C for 24 h, and 0.1 mL of the enriched culture was spread onto TSA supplemented with 0.05 g/L manganese sulfate (MnSO4; Sinopharm Chemical Reagent Co., Ltd, Shanghai, China), as described by Juneja et al. (2018). The cells were incubated at 37°C for 7 days for sporulation according to the method of Kim et al. (2017) with minor modifications. Cell sporulation was monitored via a microscope attached with a blood counting chamber until around 85% of the cells were sporulated. The spores on each plate were submerged into 2 mL of 0.1% sterile peptone water (w/v), and then harvested using a sterile loop. The spore suspension harvested from five TSA plates were combined into a conical tube, and then incubated in a water bath at 80°C for 10 min for fully inactivating the residue vegetative cells. The spore pellets were collected by centrifugation at 4000 × g for 20 min at 4°C, after which the pellets were washed three times and then resuspended in 0.1% sterile peptone water. The pores from each strain were combined in equal proportions to forming a B. cereus spore cocktail of approximately 108 spores/mL for meat inoculation.
Preparation of ready-to-eat ground beef and cinnamaldehyde supplementation
The raw ground beef was purchased from a local supermarket and stored at −18°C until use. The ground beef was confirmed to be free from B. cereus before spiking by plating on the MYP agar (MYP, Land Bridge Technology Co. Ltd, Beijing, China). Prior to boiling, the frozen meat was thawed overnight at 4°C. The meat was boiled in water containing 5 g/L NaCl and 1 g/L monosodium L-glutamate (food grade, Wamole Food Co., LTD, Shanghai, China) for 15 min. The boiled ground beef was left at room temperature to cool for a period of about an hour. Note that 2 kg of the boiled ground beef was transferred to a sterile plastic bag, and then mixed thoroughly with cinnamaldehyde (Food grade, >99% pure, Guangfu Institute of Superfine Chemical Industry, Tianjin, China) for the final concentrations of 0, 0.1, 0.5, and 1.0% (vol/wt), respectively. All the bags were stored at −18°C until theirs use within 90 days.
Inoculation
Prior to spore inoculation, the frozen boiled ground beef was thawed overnight at 4°C, and 10 gm of thawed ready-to-eat beef was transferred to a new sterile plastic bag and inoculated with 100 µL of an appropriate dilution of the prepared B. cereus spore mixture so that the final concentration was approximately 2.0 log spores/g. Thereafter, the inoculated boiled beef was thoroughly mixed manually in bags to ensure an even distribution of spores. The sample in each bag was adjusted to remove air and form an approximately 1-mm-thick layer by firmly pressing the sample against a flat surface. The bags were immediately sealed prior to bacterial growth assay.
Growth study and enumeration
The inoculated boiled ground beef samples were incubated at static temperatures of 12, 20, 28, and 36°C in an incubator (LHP-250, Hongdu Electronic Technology Co. Ltd, Shanghai, China). The samples were removed from the incubators for B. cereus enumeration at specific time intervals corresponding to the cinnamaldehyde concentration and incubation temperature. Prior to cell enumeration, each sample was mixed in duplicate with 20 mL of sterile 0.1% peptone water (PW, Land Bridge Technology Co. Ltd, Beijing, China) and homogenized for 4 min in a stomacher (SCIENTZ-11, Xinzhi Biological Technology Co. Ltd, Ningbo, China). Thereafter, 100 µL of the liquid portion of the stomached samples were plated either directly or after serial dilution in 0.85% NaCl (w/v) on MYP plate. Three independent growth experiments were performed for each condition. The bacterial colonies were counted and expressed as log CFU/g after incubation at 30°C for 24 h.
Mathematical Modeling of Bacterial Growth
Primary modeling
In the primary modeling analysis, all 16 data combinations were used to fit the inactivation curves by employing the integrated Pathogen Modeling Program (IPMP) 2013 (Huang, 2014). After thoroughly evaluating the parameters SSE, MSE, and RMSE, the growth of B. cereus was described by the Huang model that included lag, exponential, and stationary phases (Equation 1) (Huang, 2008, 2013). The equation can be expressed as
The dependent variable used in the regressions is the nature logarithm of the observed number of cells Y(t) at storage time t, where Y0 and Ymax were the initial and maximal number of cells, respectively. µmax is the specific growth rate and λ is the lag phase duration.
Secondary modeling
Three independent variables, the cinnamaldehyde concentration, temperature, and time, were considered for regressing the secondary modeling of B. cereus growth during the storage period. Thereafter, a cubic polynomial model was used to estimate the values of the lag phase duration (λ) and the specific growth rate (µmax) as described by Equations (2) and (3), which are expressed as
In Equations (2) and (3), T is the storage temperature and C is the concentration of cinnamaldehyde.
Model evaluation
The coefficient of the determination (R2), accuracy factor (Af), bias factor (Bf), and root mean squared error (RMSE) were calculated according to Equations (4)–(7), respectively. These parameters were employed to evaluate the performance of the regressed model based on the difference between the predictive and experimental values.
where ei is the error of predictive data, yi is the predictive data, and y' is the average of predictive data.
where n represents the number of trials.
Furthermore, the difference between the predictions and observations (observation – model) was calculated using the SPSS software (version 19.0, IBM SPSS Statistics, Chicago, IL) and the Microsoft Excel 2016 to determine the distribution of the residual errors and the linear fit chart, respectively.
Results and Discussion
Primary growth model development
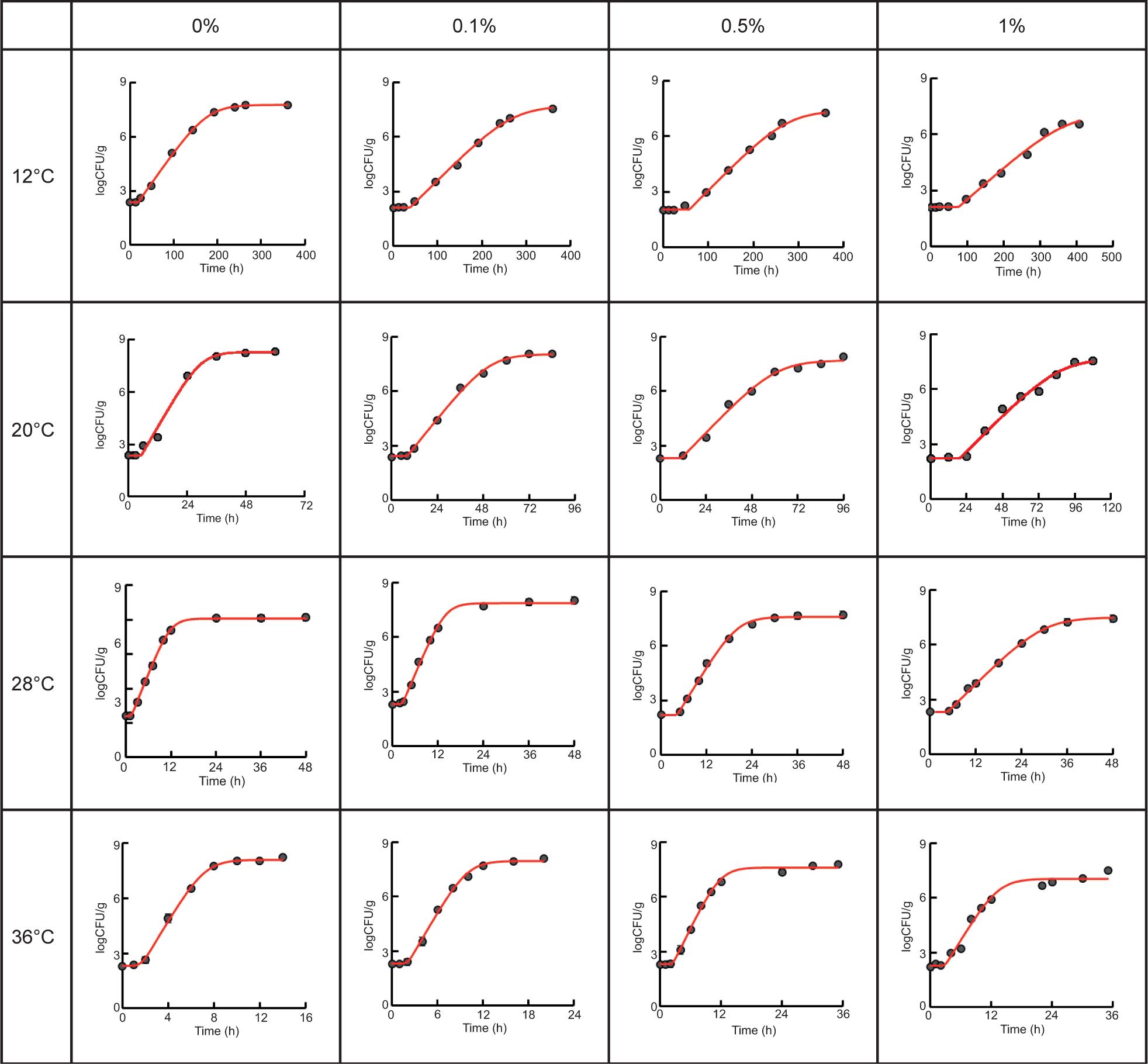
This study investigated the possible effect of the adding cinnamaldehyde and storage temperature on the germination and growth of B. cereus spores in ready-to-eat boiled beef. The fitting curves described by Huang model in IPMP program are represented in Figure 1, while the goodness-of-fit are shown in Table 1 as indicated by the values of RMSE, R2, Af, and Bf. The results showed that the values of RMSE ranged from 0.048 to 0.331, while only 1 within 16 treatment combinations were higher than 0.3. It is known that lower RMSE value normally represents a better goodness-of-fit of the models (Wang et al., 2017), although value 0.4 of RMSE is still acceptable (Jia et al., 2020). The values of Af are of the range 1.005 to 1.043 while Bf range is 0.998 to 1.004, both ranges close to 1. The Af provides an average accuracy of prediction by calling off under or over estimations. Bf value provides an overall objective indication of model performance by describing the consistency between experimental value and predicted data (Ye et al., 2013). Other studies have suggested that models with Bf values of 0.7–1.15 can be regarded as accepted for describing a pathogen growth rate (Oscar, 2005), and the growth model is regarded as quite good if Bf value determined in the range of 0.9–1.05 (Zhao et al., 2020). The determined R2 values were all higher than 0.98. Therefore, the result indicated that the RMSE, R2, Af, and Bf values were all acceptable when Huang model was used to describe the primary growth curves of B. cereus in boiled ground beef supplemented with cinnamaldehyde. Comparatively, previous reports on the B. cereus in cooked beans (Juneja et al., 2018) and cooked pasta (Juneja et al., 2019b) showed that the Baranyi model had a better performance than Huang model in describing the bacterial growth. It should be noted that both the Huang and Baranyi models are mechanistic models that allows characterizing the transition from lag phase to exponential phase, accounting for biological factors that bacteria encounter during adaption and growth (Baranyi et al., 1999; Huang, 2008).
Table 1. Experimental parameters of cinnamaldehyde inhibiting Bacillus cereus spore germination and growth in boiled ready-to-eat beef simulated by Huang model.
| Growth temperature (°C) | Concentration of cinnamaldehyde (vol/wt) (%) | λ (h) | μmax(logCFU/g per h) | Ymax(logCFU/g) | RMSE | Af | Bf | R2 |
|---|---|---|---|---|---|---|---|---|
| 12 | 0.0 | 17.730 | 0.034 | 7.780 | 0.067 | 1.009 | 1.000 | 0.999 |
| 12 | 0.1 | 38.200 | 0.024 | 7.723 | 0.107 | 1.015 | 0.998 | 0.998 |
| 12 | 0.5 | 58.189 | 0.025 | 7.416 | 0.119 | 1.023 | 1.000 | 0.998 |
| 12 | 1.0 | 75.758 | 0.017 | 7.120 | 0.184 | 1.024 | 1.000 | 0.993 |
| 20 | 0.0 | 5.221 | 0.242 | 8.285 | 0.331 | 1.040 | 1.000 | 0.991 |
| 20 | 0.1 | 8.290 | 0.132 | 8.039 | 0.134 | 1.017 | 1.001 | 0.998 |
| 20 | 0.5 | 11.194 | 0.110 | 7.685 | 0.244 | 1.030 | 1.000 | 0.993 |
| 20 | 1.0 | 19.393 | 0.082 | 7.680 | 0.274 | 1.041 | 1.004 | 0.989 |
| 28 | 0.0 | 1.466 | 0.551 | 8.086 | 0.048 | 1.005 | 1.000 | 0.999 |
| 28 | 0.1 | 2.641 | 0.494 | 7.886 | 0.141 | 1.018 | 1.001 | 0.998 |
| 28 | 0.5 | 4.254 | 0.342 | 7.588 | 0.150 | 1.017 | 1.002 | 0.997 |
| 28 | 1.0 | 4.616 | 0.210 | 7.438 | 0.079 | 1.012 | 1.001 | 0.999 |
| 36 | 0.0 | 1.626 | 1.042 | 8.113 | 0.119 | 1.015 | 1.001 | 0.999 |
| 36 | 0.1 | 2.085 | 0.724 | 7.955 | 0.153 | 1.016 | 1.001 | 0.998 |
| 36 | 0.5 | 2.524 | 0.571 | 7.594 | 0.152 | 1.014 | 1.000 | 0.997 |
| 36 | 1 | 2.923 | 0.459 | 7.035 | 0.294 | 1.043 | 1.002 | 0.987 |
Figure 1. Fitting curve of Huang model for germination and growth of Bacillus cereus spores in ready-to-eat beef with cinnamaldehyde at different temperatures.
Secondary model regression and validation
To describe the synergistic effect of temperature and cinnamaldehyde concentration on the growth of B. cereus in boiled ground beef, secondary models were regressed to predict the natural logarithm values of the lag phase duration (λ) and specific growth rate (µmax). The cubic polynomial models are presented as Equations (8) and (9) and they are
where Ln λ and Ln µmax represent the natural logarithms of the lag phase duration and the specific growth rate, respectively. T is the treatment temperature and C is the concentration of cinnamaldehyde concentration supplemented in the boiled ready-to-eat beef.
As shown in Figure 2, the two factors including the temperature and cinnamaldehyde concentration significantly influenced the growth of B. cereus during storage. The interaction between T and C was also significant, whereby an increase in cinnamaldehyde concentration required higher temperature for an equal cell population, substantiating a synergistic effect of the two parameters on the bacterial growth. Comparatively, more significant downslopes in Ln λ and upslopes in Ln µmax were observed when temperature increased from 12 to 36°C, than when the cinnamaldehyde concentration increased from 0 to 1.0%, indicating that temperature was the most important variable influencing the B. cereus growth in boiled ground beef.
Figure 2. Fitting diagram of temperature and cinnamaldehyde concentration with the lag time (Ln λ) and maximum specific growth rate (Ln μmax) of Bacillus cereus spores in boiled ready-to-eat beef.
As shown in Table 2, both the models had an acceptable goodness because the RMSE values were 0.230 and 0.203 for Ln λ and Ln µmax, respectively. The R2 values respectively were 0.965 and 0.974 (Figure 3), suggesting both equations have a satisfactory correlation between the experimental data and regression model predictions for the dependent variable of B. cereus growth. The Af values were 1.185 and 1.291 for Ln λ and Ln µmax, respectively, suggesting the predicted data were acceptably close to the experimental data. The Bf values were 1.023 and 1.086, indicating the predictions were within an acceptable range in this study (Nyhan et al., 2018).
Table 2. Goodness of fitting results of the secondary polynomial models.
| Model parameters | RMSE | F | R2 | Af | Bf |
|---|---|---|---|---|---|
| Ln λ | 0.230 | 55.641 | 0.965 | 1.185 | 1.023 |
| Ln μmax | 0.203 | 76.167 | 0.974 | 1.291 | 1.086 |
Figure 3. Predicted versus observed values of Ln λ (A) and Ln μmax (B) for the germination and growth of Bacillus cereus spores in boiled ready-to-eat beef supplemented with cinnamaldehyde.
The goodness of the regressed secondary cubic polynomial model was further evaluated by the residue errors. As shown in Figure 4, the difference between the observation and prediction values for Ln λ and Ln µmax both followed normal distributions, because the skewness and kurtosis values were all less than 1, in that the two values were 0.628 and −0.876 for Ln λ, and −0.495 and −1.076 for Ln µmax, respectively. The means of the residual errors were 3.89 × 10−16 and −5.97 × 10−16 for Ln λ and Ln µmax, respectively, while the Standard deviation (SD) value of both is 0.966. Moreover, 94% values (15 in a total of 16 observed combinations) for Ln λ and 100% values (all observed combinations) for Ln µmax of the regression standardized residual are between ±2, indicating that the predictive models are sufficiently accurate in predicting the lag phase duration and the specific growth rate of B. cereus in boiled ready-to-eat ground beef supplemented with cinnamaldehyde.
Figure 4. Histogram of residual distribution of Ln λ (A) and Ln μmax (B).
Analysis of the lag time and growth patterns
B. cereus is a foodborne pathogen and the psychrotrophic strains B. cereus could grow under low temperatures, such as at 8°C in BHI broth in aerobiosis (Guérin et al., 2016), even at 7°C in TSB broth (Samapundo et al., 2011) or TSA agar (Park et al., 2020). Comparatively, mesophilic B. cereus more likely grow at 12°C or higher temperature in a broth (Baranyi et al., 2017; Le Marc et al., 2021). Comparatively, in food assessment, the minimum growth temperature of B. cereus was 8.2°C from spores in cooked rice (Hwang and Huang, 2019). Another study proposed that the growth of B. cereus was only happened when the temperature was higher than 13°C during cooling of cooked rice (Juneja et al., 2019a). However, B. cereus can only grow slightly at a low temperature as 5°C in natural carrot substrate (Valero et al., 2000). In this study, B. cereus can grow to the maximum population at 12°C in boiled ready-to-eat ground beef, which should be attributed that the food-derived ingredient, such as fatty acids, offer a protection on the membrane, thus improving the growth of cells under cold conditions (de Sarrau et al., 2013).
The determination of primary and second model parameters provided statistical information about the effects of the environmental factors, including the temperature and cinnamaldehyde concentration, on the lag time and growth rate of B. cereus in boiled beef during storage. As shown, the microbial growth was temperature dependent. Higher storage temperatures facilitated the growth of B. cereus as evidenced by the increasing values of µmax and Ymax, and lower λ value, compared to their corresponding 12°C treatments. For example, the determined µmax, Ymax, and λ values were 0.034 log CFU/g per h, 7.780 log CFU/g, and 17.730 h, respectively, at 12°C-treated sample without cinnamaldehyde supplementary. However, the values of the corresponding samples were 1.042 log CFU/g per h, 8.113 log CFU/g, and 1.626 h when treated at 36°C. The highest specific growth rate observed at 36°C-treated sample without cinnamaldehyde supplementary was comparable to the model evaluated result of 0.96 log CFU/g per h in cooked rice at 37.6°C (Hwang and Huang, 2019). The agreed results are another indication that the growth rates of B. cereus were similar between the independent data observed in meat- and cereal- based matrices (Ellouze et al., 2021).
Additives are normally used restrictively in food products, and then researchers keep optimizing their usage and dose to pursue their maximum antibacterial effect. Nonoptimal low temperatures normally impose additional damage on bacterial cells, and thus more effectively inhibit the growth of bacterial cells (Suo et al., 2017a). In present study, compared to the lower storage temperature (12°C), along with the augment of cinnamaldehyde concentration from 0 to 1%, the increase of λ value was more significant than that individually stored at higher temperature, while the highest increase of 3.27-fold was observed at 12°C. The µmax value declined most obviously at 20°C, while 66% decrease was determined. The result indicated that the additive cinnamaldehyde functioned as an inhibitor on B. cereus germination and growth most effectively at suboptimal temperature. In detail, the most effective contribution of cinnamaldehyde in boiled ground beef at 12°C is supposed to prolong the lag time of B. cereus, while is to inhibit its growth at 20°C. In carrot broth, according to the regressed Baranyi model, the cinnamaldehyde prolonged 33–301% lag phase of B. cereus at 16°C (Valero and Francés, 2006).
Conclusions
The study quantitatively described the effect of the cinnamaldehyde on the germination and growth of B. cereus spores in boiled ready-to-eat ground beef. The Huang model can be used to predict the lag time and maximum growth rate with high accuracy. The supplementary of cinnamaldehyde in samples increased the lag time of B. cereus, significantly from 17.7 to 75.8 h at 12°C of storage. The cubic polynomial models were successfully used to estimate the values of Ln λ and Ln µmax considering both the storage temperature and cinnamaldehyde concentration. According to the results, the cinnamaldehyde can be used as a natural antimicrobial agent in boiled ready-to-eat ground beef in staple food industry which inhibits the germination and growth of B. cereus. The results also provided regressing models that can be used effectively for designing the storage temperature and cinnamaldehyde concentration for a specific requirement.
Acknowledgments
The authors wish to thank the Department of Human Resources and Social Security of Henan Province for supporting this research.
Conflict of interest
There is no conflict of interest.
REFERENCES
Alonso, V.P.P., de Oliveira Morais, J. and Kabuki, D.Y., 2021. Incidence of Bacillus cereus, Bacillus sporothermodurans and Geobacillus stearothermophilus in ultra-high temperature milk and biofilm formation capacity of isolates. International Journal of Food Microbiology 354: 109318. 10.1016/j.ijfoodmicro.2021.109318
Ayari, S., Dussault, D., Jerbi, T., Hamdi, M. and Lacroix, M., 2012. Radiosensitization of Bacillus cereus spores in minced meat treated with cinnamaldehyde. Radiation Physics and Chemistry 81: 1173–1176. 10.1016/j.radphyschem.2012.02.022
Baranyi, J., Buss da Silva, N. and Ellouze, M., 2017. Rethinking tertiary models: relationships between growth parameters of Bacillus cereus strains. Frontiers in Microbiology 8: 1890. 10.3389/fmicb.2017.01890
Baranyi, J., Pin, C. and Ross, T., 1999. Validating and comparing predictive models. International Journal of Food Microbiology 48: 159–166. 10.1016/S0168-1605(99)00035-5
Cayemitte, P.E., Gerliani, N., Raymond, P. and Aider, M., 2021. Study of the impacts of electro-activated solutions of calcium lactate, calcium ascorbate and their equimolar mixture combined with moderate heat treatments on the spores of Bacillus cereus ATCC 14579 under model conditions and in fresh salmon. International Journal of Food Microbiology 358: 109285. 10.1016/j.ijfoodmicro.2021.109285
Cetin-Karaca, H. and Morgan, M.C., 2018. Inactivation of Bacillus cereus spores in infant formula by combination of high pressure and trans-cinnamaldehyde. LWT-Food Science and Technology 97: 254–260. 10.1016/j.lwt.2018.07.001
Chitov, T., Dispan, R. and Kasinrerk, W., 2008. Incidence and diarrhegenic potential of Bacillus cereus in pasteurized milk and cereal products in Thailand. Journal of Food Safety 28: 467–481. 10.1111/j.1745-4565.2008.00125.x
de Sarrau, B., Clavel, T., Zwickel, N., Despres, J., Dupont, S., Beney, L., et al., 2013. Unsaturated fatty acids from food and in the growth medium improve growth of Bacillus cereus under cold and anaerobic conditions. Food Microbiology 36: 113–122. 10.1016/j.fm.2013.04.008
Doyle, A.A. and Stephens, J.C., 2019. A review of cinnamaldehyde and its derivatives as antibacterial agents. Fitoterapia 139: 104405. 10.1016/j.fitote.2019.104405
Ellouze, M., Buss Da Silva, N., Rouzeau-Szynalski, K., Coisne, L., Cantergiani, F. and Baranyi, J., 2021. Modeling Bacillus cereus growth and cereulide formation in cereal-, dairy-, meat-, vegetable-based food and culture medium. Frontiers in Microbiology 12. 10.3389/fmicb.2021.639546
Friedman, M., 2017. Chemistry, antimicrobial mechanisms, and antibiotic activities of cinnamaldehyde against pathogenic bacteria in animal feeds and human foods. Journal of Agricultural and Food Chemistry 65: 10406–10423. 10.1021/acs.jafc.7b04344
Guérin, A., Dargaignaratz, C., Broussolle, V., Clavel, T. and Nguyen-the, C., 2016. Combined effect of anaerobiosis, low pH and cold temperatures on the growth capacities of psychrotrophic Bacillus cereus. Food Microbiology 59: 119–123. 10.1016/j.fm.2016.05.015
Hou, J., Luo, R., Ni, H., Li, K., Mgomi, F.C., Fan, L. et al., 2021. Antimicrobial potential of kombucha against foodborne pathogens: a review. Quality Assurance and Safety of Crops & Foods 13: 53–61. 10.15586/qas.v13i3.920
Huang, L., 2008. Growth kinetics of Listeria monocytogenes in broth and beef frankfurters–determination of lag phase duration and exponential growth rate under isothermal conditions. Journal of Food Science 73: E235–E242. 10.1111/j.1750-3841.2008.00785.x
Huang, L., 2013. Optimization of a new mathematical model for bacterial growth. Food Control 32: 283–288. 10.1016/j.foodcont.2012.11.019
Huang, L., 2014. IPMP 2013–a comprehensive data analysis tool for predictive microbiology. International Journal of Food Microbiology 171: 100–107. 10.1016/j.ijfoodmicro.2013.11.019
Hwang, C.-A. and Huang, L., 2019. Growth and survival of Bacillus cereus from spores in cooked rice–One-step dynamic analysis and predictive modeling. Food Control 96: 403–409. 10.1016/j.foodcont.2018.09.036
Jia, Z., Bai, W., Li, X., Fang, T. and Li, C., 2020. Assessing the growth of Listeria monocytogenes in salmon with or without the competition of background microflora–A one-step kinetic analysis. Food Control: 107139. 10.1016/j.foodcont.2020.107139
Juneja, V.K., Golden, C.E., Mishra, A., Harrison, M.A., Mohr, T. and Silverman, M., 2019a. Predictive model for growth of Bacillus cereus during cooling of cooked rice. International Journal of Food Microbiology 290: 49–58. 10.1016/j.ijfoodmicro.2018.09.023
Juneja, V.K., Golden, C.E., Mishra, A., Harrison, M.A. and Mohr, T.B., 2019b. Predictive model for growth of Bacillus cereus at temperatures applicable to cooling of cooked pasta. Journal of Food Science 84: 590–598. 10.1111/1750-3841.14448
Juneja, V.K., Mishra, A. and Pradhan, A.K., 2018. Dynamic predictive model for growth of Bacillus cereus from spores in cooked beans. Journal of Food Protection 81: 308–315. 10.4315/0362-028X.JFP-17-391
Kim, J.E., Choi, H.S., Lee, D.U. and Min, S.C., 2017. Effects of processing parameters on the inactivation of Bacillus cereus spores on red pepper (Capsicum annum L.) flakes by microwave-combined cold plasma treatment. International Journal of Food Microbiology 263: 61–66. 10.1016/j.ijfoodmicro.2017.09.014
Kindle, P., Etter, D., Stephan, R. and Johler, S., 2019. Population structure and toxin gene profiles of Bacillus cereus sensu lato isolated from flour products. FEMS Microbiology Letters 366. 10.1093/femsle/fnz240
Kong, L., Yu, S., Yuan, X., Li, C., Yu, P., Wang, J., et al., 2021. An investigation on the occurrence and molecular characterization of Bacillus cereus in meat and meat products in China. Foodborne Pathogens and Disease 18: 306–314. 10.1089/fpd.2020.2885
Le Marc, Y., Baert, L., Buss da Silva, N., Postollec, F., Huchet, V., Baranyi, J. et al., 2021. The effect of pH on the growth rate of Bacillus cereus sensu lato: Quantifying strain variability and modelling the combined effects of temperature and pH. International Journal of Food Microbiology 360: 109420. 10.1016/j.ijfoodmicro.2021.109420
Nyhan, L., Begley, M., Mutel, A., Qu, Y., Johnson, N. and Callanan, M., 2018. Predicting the combinatorial effects of water activity, pH and organic acids on Listeria growth in media and complex food matrices. Food Microbiology 74: 75–85. 10.1016/j.fm.2018.03.002
Omidi-Mirzaei, M., Hojjati, M., Behbahani, B.A. and Noshad, M., 2020. Modeling the growth rate of Listeria innocua influenced by coriander seed essential oil and storage temperature in meat using FTIR. Quality Assurance and Safety of Crops & Foods 12: 1–8. 10.15586/qas.v12iSP1.776
Oscar, T.E., 2005. Validation of lag time and growth rate models for Salmonella Typhimurium: acceptable prediction zone method. Journal of Food Science 70: M129–M137. 10.1111/j.1365-2621.2005.tb07103.x
Park, H.W. and Yoon, W.B., 2019. A quantitative microbiological exposure assessment model for Bacillus cereus in pasteurized rice cakes using computational fluid dynamics and Monte Carlo simulation. Food Research International 125: 108562. 10.1016/j.foodres.2019.108562
Park, K.M., Kim, H.J., Jeong, M. and Koo, M., 2020. Enterotoxin genes, antibiotic susceptibility, and biofilm formation of low-temperature-tolerant Bacillus cereus isolated from green leaf lettuce in the cold chain. Foods 9: 249. 10.3390/foods9030249
Rodrigo, D., Rosell, C.M. and Martinez, A., 2021. Risk of Bacillus cereus in relation to rice and derivatives. Foods 10: 302. 10.3390/foods10020302
Samapundo, S., Heyndrickx, M., Xhaferi, R. and Devlieghere, F., 2011. Incidence, diversity and toxin gene characteristics of Bacillus cereus group strains isolated from food products marketed in Belgium. International Journal of Food Microbiology 150: 34–41. 10.1016/j.ijfoodmicro.2011.07.013
Sun, Q., Li, J., Sun, Y., Chen, Q., Zhang, L. and Le, T., 2020. The antifungal effects of cinnamaldehyde against Aspergillus niger and its application in bread preservation. Food Chemistry 317: 126405. 10.1016/j.foodchem.2020.126405
Suo, B., Li, H., Wang, Y., Li, Z., Pan, Z. and Ai, Z., 2017a. Effects of ZnO nanoparticle-coated packaging film on pork meat quality during cold storage. Journal of the Science of Food and Agriculture 97: 2023–2029. 10.1002/jsfa.8003
Suo, B., Lu, Y.L., Wang, Y.X., Xie, X.H., Xu, C. and Ai, Z.L., 2017b. Thermal inactivation kinetics of Salmonella spp. in ground pork supplemented with cinnamaldehyde. Journal of Food Safety 37: e12322. 10.1111/jfs.12322
Tewari, A., Singh, S.P. and Singh, R., 2015. Incidence and enterotoxigenic profile of Bacillus cereus in meat and meat products of Uttarakhand, India. Journal of Food Science and Technology 52: 1796–1801. 10.1007/s13197-013-1162-0
Valero, M. and Francés, E., 2006. Synergistic bactericidal effect of carvacrol, cinnamaldehyde or thymol and refrigeration to inhibit Bacillus cereus in carrot broth. Food Microbiology 23: 68–73. 10.1016/j.fm.2005.01.016
Valero, M., Leontidis, S., Fernández, P.S., Martı´nez, A. and Salmerón, M.C., 2000. Growth of Bacillus cereus in natural and acidified carrot substrates over the temperature range 5–30°C. Food Microbiology 17: 605–612. 10.1006/fmic.2000.0352
Wang, J., Koseki, S., Chung, M.J. and Oh, D.H., 2017. A novel approach to predict the growth of Staphylococcus aureus on rice cake. Frontiers in Microbiology 8: 1140. 10.3389/fmicb.2017.01140
Wang, Y., Li, X., Lu, Y., Wang, J. and Suo, B., 2020. Synergistic effect of cinnamaldehyde on the thermal inactivation of Listeria monocytogenes in ground pork. Food Science and Technology International 26: 28–37. 10.1177/1082013219867190
Wemmenhove, E., Wells-Bennik, M.H.J. and Zwietering, M.H., 2021. A model to predict the fate of Listeria monocytogenes in different cheese types–A major role for undissociated lactic acid in addition to pH, water activity, and temperature. International Journal of Food Microbiology 357: 109350. 10.1016/j.ijfoodmicro.2021.109350
Ye, K., Wang, H., Zhang, X., Jiang, Y., Xu, X. and Zhou, G., 2013. Development and validation of a molecular predictive model to describe the growth of Listeria monocytogenes in vacuum-packaged chilled pork. Food Control 32: 246–254. 10.1016/j.foodcont.2012.11.017
Zhao, X., Chen, L., Zhao, L., He, Y. and Yang, H.S., 2020. Antimicrobial kinetics of nisin and grape seed extract against inoculated Listeria monocytogenes on cooked shrimps: Survival and residual effects. Food Control 115: 107278. 10.1016/j.foodcont.2020.107278