
Download
RESEARCH ARTICLE
Regulatory effect of intracellular polysaccharides from Antrodia cinnamomea on the intestinal microbiota of mice with antibiotic-associated diarrhea
Chun-lei Lu1#, Hua-xiang Li1#, Xiao-yan Zhu2, Zhi-shan Luo3, Sheng-qi Rao1, Zhen-quan Yang1, 4*
1College of Food Science and Engineering, Yangzhou University, Yangzhou, China;
2Jiangsu Provincial Key Construction Laboratory of Probiotics Preparation, Huaiyin Institute of Technology, Huaian, China;
3School of Biotechnology, Jiangnan University, Wuxi, China;
4Jiangsu Key Laboratory of Dairy Biotechnology and Safety Control, Yangzhou University, Yangzhou China
Abstract
Antrodia cinnamomea is a valuable and scarce medicinal and edible mushroom. Its main active substances are triterpenoids and polysaccharides with many biological activities, such as liver protection, immunity enhancement, antitumor, anti-inflammatory, and antiviral. The long-term use of antibiotics can easily lead to disorders in the intestinal flora and causes diarrhea, inflammation, immune decline, and other symptoms. In this study, A. cinnamomea intracellular polysaccharides (AIPSs) were extracted from mycelia in submerged fermentation, and the AIPSs with a neutral polysaccharide content of 82.74%±0.63% were obtained. Then, the therapeutic effect of AIPSs on diarrhea in mice caused by lincomycin hydrochloride (LIH), and their regulatory and recovery effects on the intestinal flora, was studied with fructooligosaccharide as the positive control. In vivo experiments in mice showed that fed intragastrically with 0.25 g/kg (bodyweight of mice) of AIPSs significantly reduced the weight loss caused by antibiotics and restored the immune-organ index (P<0.05). The AIPSs also significantly lowered the degrees of the inflammatory cytokines IL-6 and TNF-α in mouse serum (P<0.01). Analysis of mouse intestinal flora showed that AIPSs treated diarrhea caused by LIH and regulated and improved the flora’s structure. These effects were primarily manifested in the increased relative abundance of good microorganisms (e.g., Lachnospiraceae_NK4A136_group, Osllospiraceae, and Lachnospiraceae) and reduced relative abundance of harmful microorganisms (e.g., Enterococcus) (P<0.05). This study further expanded the application value of the submerged fermentation products of A. cinnamomea. A new perspective and theoretical basis for developing new multifunctional prebiotics to regulate antibiotic-induced disorders in the intestinal flora was also provided.
Key words: Antrodia cinnamomea, intracellular polysaccharide, lincomycin hydrochloride, diarrhea, intestinal flora
*Corresponding Author: Zhen-quan Yang, No. 196 West Huayang Road, Yangzhou, Jiangsu 225009, China. Email: [email protected]
#These authors contributed equally to this study and share first authorship.
Submitted: 18 February 2022; Accepted: 29 March 2022; Published: 22 June 2022
Doi: http://dx.doi.org/10.15586/qas.v14i3.1073
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
The human intestinal flora is a microbial ecosystem comprising trillions of bacteria. This ecosystem exerts an extremely significant effect on the metabolism and immunity regulation of the host, especially in early child development (Rajakovich et al., 2019). One of the most significant reasons leading to intestinal disease is the use of antibiotics. During the long-term use of antibiotics, many beneficial bacteria are lost, causing drug-resistant, harmful microorganisms to dominate gradually. The ultimate result is a sharp increase in the risk of chronic diseases (Gao et al., 2017; Pelaseyed et al., 2020). Important measures to prevent and treat chronic diseases include the maintenance of the balance of the intestinal flora and the regulation of its structure. Studies have shown that prebiotics can boost the proliferation of beneficial bacteria in the intestine. Everard et al. found that a functional oligosaccharide prebiotic, namely, fructooligosaccharide (FOS), could increase the abundance of Akkermansia muciniphila in the mouse intestine. FOS can also enhance the intestine’s barrier function and reduce inflammatory symptoms (Everard et al., 2013). Fungal polysaccharides have prebiotic roles such as functional oligosaccharides, mostly manifested as the promotion of proliferation of good bacteria and the inhibition of pathogenic bacterial growth (Liang et al., 2021). Edible and medicinal fungi are some of the main sources of functional fungal polysaccharides. These polysaccharides with prebiotic effects include glucans, polysaccharide peptide /protein complex, heteroglycan, chitin, and other indigestible polysaccharides (Wu et al., 2019).
Antrodia cinnamomea (syn. Antrodia camphorata and Taiwanofungus camphoratus) is a precious fungus used as medicine and food. This fungus belongs to Antrodia, Polyporaceae, and Basidiomycetes. A. cinnamomea exhibits anti-inflammation (Lu et al., 2013) and antitumor (Song et al., 2005; Yen et al., 2015) activities. It is also used to treat liver diseases (Liu et al., 2017b) and alcoholism (Yang et al., 2015), lower blood pressure (Chen et al., 2018), reduce obesity (Chang et al., 2018), and resist fatigue (Liu et al., 2017a). The main active substances in A. cinnamomea are triterpenoids and polysaccharides (Geethangili et al., 2011; Ganesan et al., 2019).
As a fungal polysaccharide, A. cinnamomea polysaccharide has various organisms and activities. For instance, the polysaccharides of A. cinnamomea were confirmed to have anti-hepatitis B viral (Ker et al., 2014), antivascular growth, antitumor, and other clinical effects (Yang et al., 2009; Zhang et al., 2018b). The polysaccharides of A. cinnamomea also have therapeutic potential as an adjuvant in the immunomodulation treatment of allergic asthma (Liu et al., 2010). In addition, the water-soluble polysaccharides comprising β-glucan in A. cinnamomea can alleviate the inflammatory response induced by lipopolysaccharide (Chen et al., 2017).In the present study, the therapeutic and regulatory or restoration effects of intracellular polysaccharide from A. cinnamomea on antibiotic-induced diarrhea and disorder in mouse intestinal flora were investigated. A new concept and theoretical basis for developing novel multifunctional prebiotics were provided.
Materials and Methods
Materials and experimental animals
The A. cinnamomea strain was purchased from the American Type Culture Collection (No. 200183). Absolute alcohol, n-butanol, trichloromethane, phenol, sulfuric acid were purchased from Sinopharm Chemical Reagent Co., Ltd. lincomycin hydrochloride (LIH) and fructooligosaccharide (FOS) were purchased from Shanghai Macklin Biochemical Co., Ltd. These chemical reagents were analytical grade.
The Comparative Medicine Centre of Yangzhou University (Yangzhou, China) provided 42 Female ICR mice (specific pathogen-free, 6 weeks old, and weighing 20±2 g). All mice received humanitarian care. All animal experiments were conducted based on the related rules of the Experimental Animal Ethics Committee of Yangzhou University (SYXK2021-0019).
Extraction and purification of AIPSs
After submerged fermentation for 12 days with 150 r/min at 26°C (Li et al., 2015), the fermentation broth of A. cinnamomea was screened with four layers of gauze. The mycelia were collected and crushed, then extracted with water at a solid-liquid ratio of 1 to 10 at 90°C for 2 hours. Subsequently, the extract was centrifuged at 6000 r/min for 10 min at 4°C and the supernatant was collected. Then, anhydrous ethanol was added to the supernatant at a volume ratio (v/v) of 1:3, and the solution was allowed to stand overnight at 4°C. The sediment was centrifuged at 8000 r/min at 4°C for 10 min, collected, and dried at 75°C, and then the AIPSs were obtained (Hooper et al., 2010).
Determination of polysaccharide and protein content
With glucose as a standard, the total neutral sugar in the AIPSs was determined with the phenol–sulfuric acid approach (Masuko et al., 2005). The total protein in the AIPSs was quantified using a BCA protein kit (Nanjing Jiancheng Bioengineering Institute).
Effect of AIPSs on the Intestinal Microbiota of Mice with Diarrhea
Feeding of mice
All ICR mice (5–7 weeks old, weighing 20±2 g, and female) moved freely at the room temperature of 24±2°C, relative humidity of 55%±5%, and light/dark cycle for 12 h. The food, water, and disinfection pad were changed daily at 8:00 p.m. The mice were fed separately in cages, and basic food and water were provided ad libitum for each cage. After adaptive feeding for 1 week, mice that could eat and move normally were selected for the -follow-up experiment.
Construction of a mouse model of diarrhea
A total of 42 mice were randomly grouped into six (n=7), namely, the normal control (NC), model group (RG), low dose of AIPSs treatment group (AP-L), a middle dose of AIPSs treatment group (AP-M), high dose of AIPSs treatment group (AP-H) and FOS positive-control group (FOS). On days 1–3, the NC group was fed with sterile normal saline at 3.7 g/kg (bodyweight of mice), whereas the RG, AP-L, AP-M, AP-H, and FOS groups were fed with LIH at a dose of 3.7 g/kg (bodyweight of mice). Gavage was administered twice a day at 9:00 a.m. and 7:00 p.m. In the next 4–11 days, the NC and RG groups were fed intragastrically with sterile normal saline at 1.25 g/kg (bodyweight of mice) at 7:00 p.m.; the AP-L and AP-M group were fed intragastrically with AIPSs at 0.05 g/kg and 0.25 g/kg (bodyweight of mice) respectively; the AP-H and FOS group were accordingly fed intragastrically with AIPSs or FOS at 1.25 g/kg (bodyweight of mice).
General observation of diarrhea
On days 4–6 (1–3 days after discontinuation of medication), feces were collected. Based on the mouse state and fecal shape, the diarrhea symptoms caused by LIH were evaluated, and the severity of diarrhea symptoms in the various groups was compared. Diarrhea symptoms were scored as follows: 0 (normal state or no symptom), 1 (in general conditions, with a small amount of wet and shapeless feces), 2 (down in spirit, with substantial wet and shapeless feces), and 3 (depressed, with substantial severely watery feces).
Determination of immune-organ index
On day 12, all mice were weighed and then killed. The spleen and thymus were separated and weighed. The spleen index (spleen mass/body weight, mg/kg) and thymus index (thymus mass/body weight, mg/kg) were calculated.
Determination of inflammatory factors
The inflammatory factors IL-6 and TNF-α in mouse serum were determined using a mouse TNF-α ELISA Kit and a mouse IL-6 ELISA Kit (SenBeiJia Biological Technology Co., Ltd.). Eyeballs were enucleated before the mice were killed on day 12, and then their blood was collected and centrifuged for 20 min at 3000 r/min and 4°C. Afterward, the light-yellow supernatant was collected for detection. The sample serum was placed in solid-state antibody microplate wells, and an enzyme label was added to detect the antibody. Then, matrix TMB was put to vary the color from blue to yellow. The absorbance value at 450 nm wavelength was measured, and the concentration of every indicator was calculated.
Determination of intestinal flora
On day 12, the mice were dissected, and samples of cecal contents from the mouse were collected. After quick freezing with liquid nitrogen, the samples were placed in dry ice and sent to the Beijing Novogene Bioinformatics Technology Co., Ltd. (Novogene, Beijing, China) for high-throughput sequencing. The sequencing platform used was Illumina. The instrument was NovaSeq6000. A polymerase chain reaction was adopted to amplify the V3–V4 area of the 16 S rDNA gene by applying a forward primer (5'-CCTACGGGNGGCWGCAG-3') and a reverse primer (5'-GGACTACHVGGGTATCTAAT-3'). The initial DNA fragments were read using Flash software, and the quality of splicing data was controlled using QIIM software. Then, the operational taxonomic unit clustering and analysis of species classification were performed for the useful tags of all samples with 97% identity.
Statistical analysis
Data are shown as the mean ± SD and statistically compared to decide the significant differences (P<0.05) using one-way ANOVA with SPSS 22.0 software. The graphs were plotted using Origin 2021.
Results
Content analysis of AIPSs
The polysaccharide and protein contents of AIPSs were determined with the phenol–sulfuric acid approach and the BCA protein kit, and the contents of neutral polysaccharide and protein were 82.74%±0.63% and 1.32%±0.11%, respectively. The results showed that the purity of AIPSs obtained here was high enough.
Impact of AIPSs on diarrhea and immune organs of mice
Table 1 shows the scores of diarrhea and immune-organ indices. Great diversities were observed in the thymus and spleen indices between the RG and NC groups (P<0.05), which means LIH had an immunosuppressive effect on the thymus and spleen of mice. Nevertheless, no significant difference was found in the thymus and spleen indices between the AP-L, AP-M, AP-H, and NC groups (P>0.05), which means the immune-organ indices returned to their normal levels after feeding the mice with AIPSs.
Table 1. Diarrhea scores and immune organ indices of mice.
| Groups | Diarrhea score | Thymus index (mg/g) | Spleen index (mg/g) |
|---|---|---|---|
| NC | 0a | 1.587±0.008a | 3.451±0.305b |
| RG | 2±0.5b | 0.687±0.280b | 4.737±0.377a |
| AP-L | 1.1±0.3a | 1.349±0.331a | 4.135±0.362b |
| AP-M | 0.6±0.4a | 1.428±0.202a | 3.554±0.301b |
| AP-H | 1.5±0.3a | 1.302±0.273a | 4.146±0.285b |
| FOS | 1.2±0.3a | 1.283±0.283a | 3.651±0.318b |
Note: NC, normal group; RG, model group; AP-L, low dose of Antrodia cinnamomea intracellular polysaccharide treatment group; AP-M, middle dose of Antrodia cinnamomea intracellular polysaccharide treatment group; AP-H, high dose of Antrodia cinnamomea intracellular polysaccharide treatment group; and FOS, fructooligosaccharide positive-control group. Each data group is shown as the mean ± standard deviation and explored by one-way ANOVA. Various letters in the same column show significant differences (P<0.05); n=7.
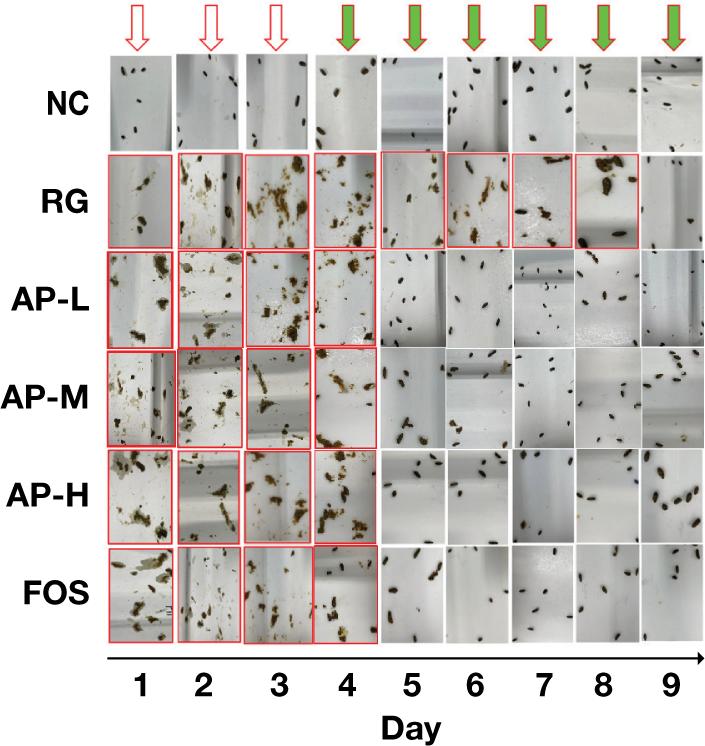
Moreover, diarrhea scores decreased after feeding the mice with AIPSs, and the score greatly differed from that of the RG group (P<0.05). This result suggested that AIPSs benefited the alleviation of the LIH-induced diarrhea of mice. Analysis of the fecal shape of mice (Figure 1) revealed that after the intragastric administration of LIH, the RG group had continuous diarrhea reaction until day 8. Conversely, the diarrhea of the AIPS and FOS groups significantly improved. The feces of all groups returned to normal after 9 days.
Figure 1. State of mouse feces. NC, normal group; RG, model group; AP-L, low dose of Antrodia cinnamomea intracellular polysaccharide treatment group; AP-M, middle dose of Antrodia cinnamomea intracellular polysaccharide treatment group; AP-H, high dose of Antrodia cinnamomea intracellular polysaccharide treatment group; FOS, FOS positive-control group. The red hollow arrow indicates the period of intragastric feeding of LIH. The green arrow indicates the intragastric feeding after drug withdrawal. The red box indicates the abnormal feces of mice (diarrhea). The feces of all mice returned to normal after 9 days.
Impact of AIPSs on the bodyweight of mice
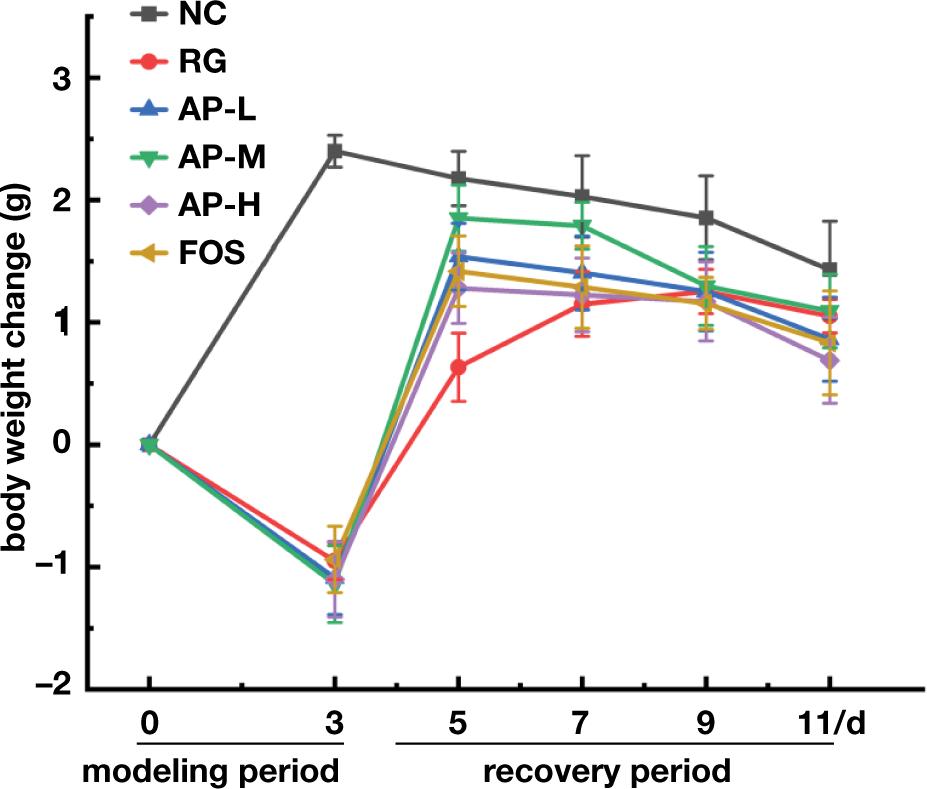
On days 1–3, compared with mice in the NC group, the bodyweight of mice intragastrically fed with LIH significantly decreased, indicating that LIH negatively affected mouse growth. After the medication was discontinued (4–7 days), the bodyweight of the mice gradually increased, but the AP-M group showed greatly higher recovery and growth rates and body weight than the RG group (Figure 2). This outcome indicated that the intragastric administration of AIPSs was beneficial in improving the body-weight loss caused by LIH, and the effect was equivalent to that of FOS. After 7 days, the weight growth of all groups stabilized.
Figure 2. Effects of AIPSs on the change in average body weight.
Impact of AIPSs on the inflammatory factors of mice
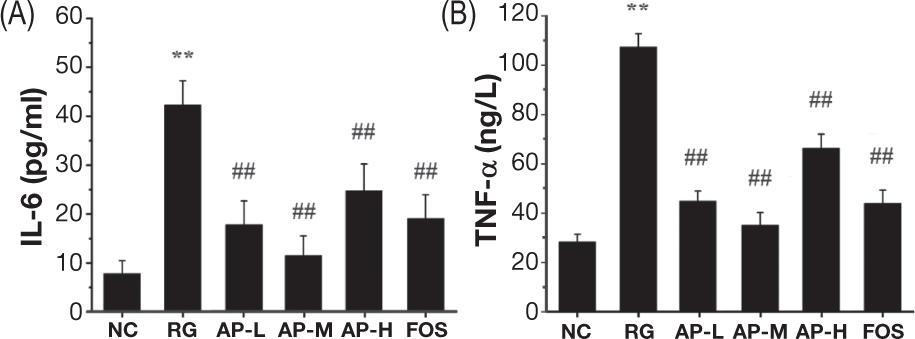
The inflammatory factors in mouse serum were assessed by detecting the degrees of IL-6 and TNF-α (Figure 3). The RG group showed greatly higher levels of these two factors in sera than the NC group (P<0.01), suggesting that LIH could lead to severe inflammatory symptoms in mice. The degrees of IL-6 and TNF-α in the sera of mice intragastrically fed with AIPSs and FOS were significantly lower than those in the RG group (P<0.01). This result implied that AIPSs benefited the treatment or alleviation of the inflammatory symptoms caused by LIH, and the effect of the middle dose of AIPSs was even better than that of FOS.
Figure 3. Impact of AIPSs on the inflammatory factors of mice. Each data group is shown as the mean ± standard deviation and explored by one-way ANOVA. Compared with the blank control group, * represents that the diversity is great (P<0.05), and ** represents that the diversity is extremely great (P<0.01). By comparing with the model group, # represents that the diversity is great (P<0.05), and ## represents that the diversity is extremely great (P<0.01) in all groups, n=7.
Effect of AIPSs on the Intestinal Microbiota of Mice with Diarrhea
α-Diversity and β-diversity analyses of intestinal microbiota
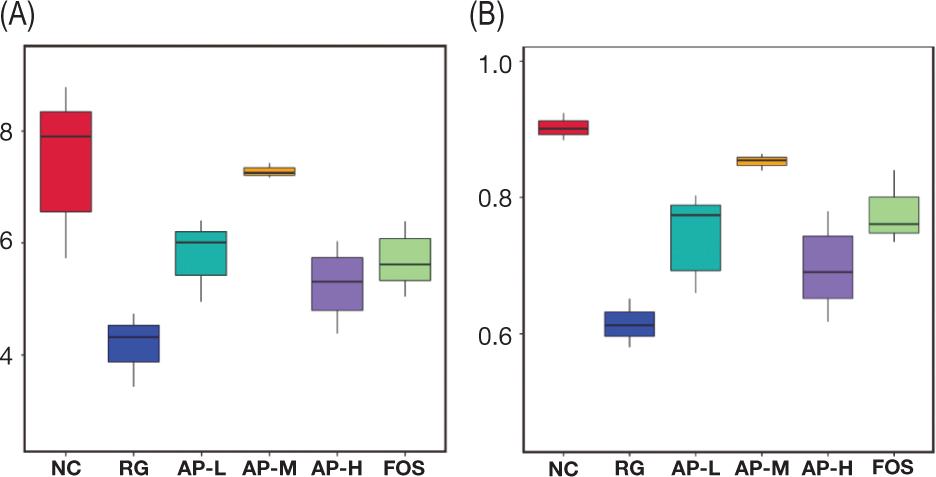
To understand the richness and uniformity of species in the intestinal floras of different groups, α-diversity analyses were performed (Figure 4). Results showed that the difference in the intestinal flora of the RG group was significantly lower than that of the NC group. Thus, LIH seriously destroyed the balance in mouse intestinal flora and inhibited or killed a substantial amount of microorganisms. Intragastric administration of AIPSs and FOS helped regulate and restore the difference in mouse intestinal flora, and the effect of the AP-M group was significantly better than that of the FOS group. Shannon index (Figure 4A) and Simpson index (Figure 4B) revealed that after the intragastric administration of AIPSs, the diversity of mouse intestinal flora was well restored. However, the diversity in the intestinal flora of mice fed with FOS cannot be restored to the level of the NC group.
Figure 4. Shannon (A) and Simpson (B) indices of the α-diversity analysis of intestinal microbiota.
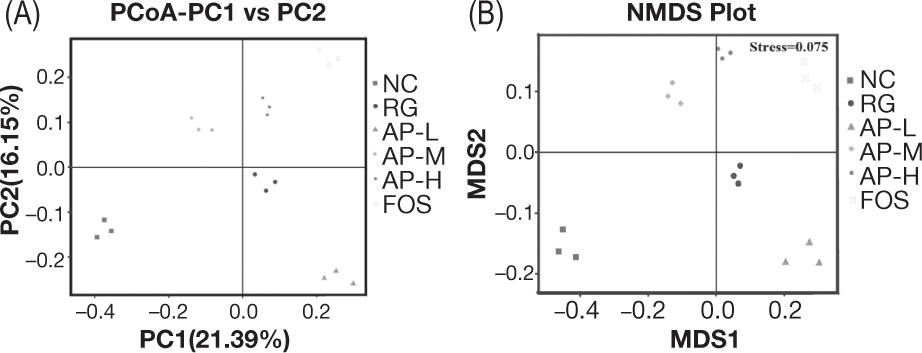
To compare the differences in structures of intestinal floras among the different treatment groups, β-diversity analyses were performed (Figure 5). Results showed significant differences in the structures of intestinal floras among the treatment groups. Thus, the intragastric administration of LIH, AIPSs, and FOS played significant roles in the intestinal floras in mice. AIPSs also played an obvious regulatory role in the diversity of mouse intestinal flora. Repeated samples of each group were also relatively concentrated, indicating that all samples had good repeatability.
Figure 5. β-Diversity analyses of intestinal microbiota. (A) principal coordinate analysis (PCoA); (B) non-metric multidimensional scaling (NMDS) analysis.
Structural analysis of intestinal microbial flora in mice
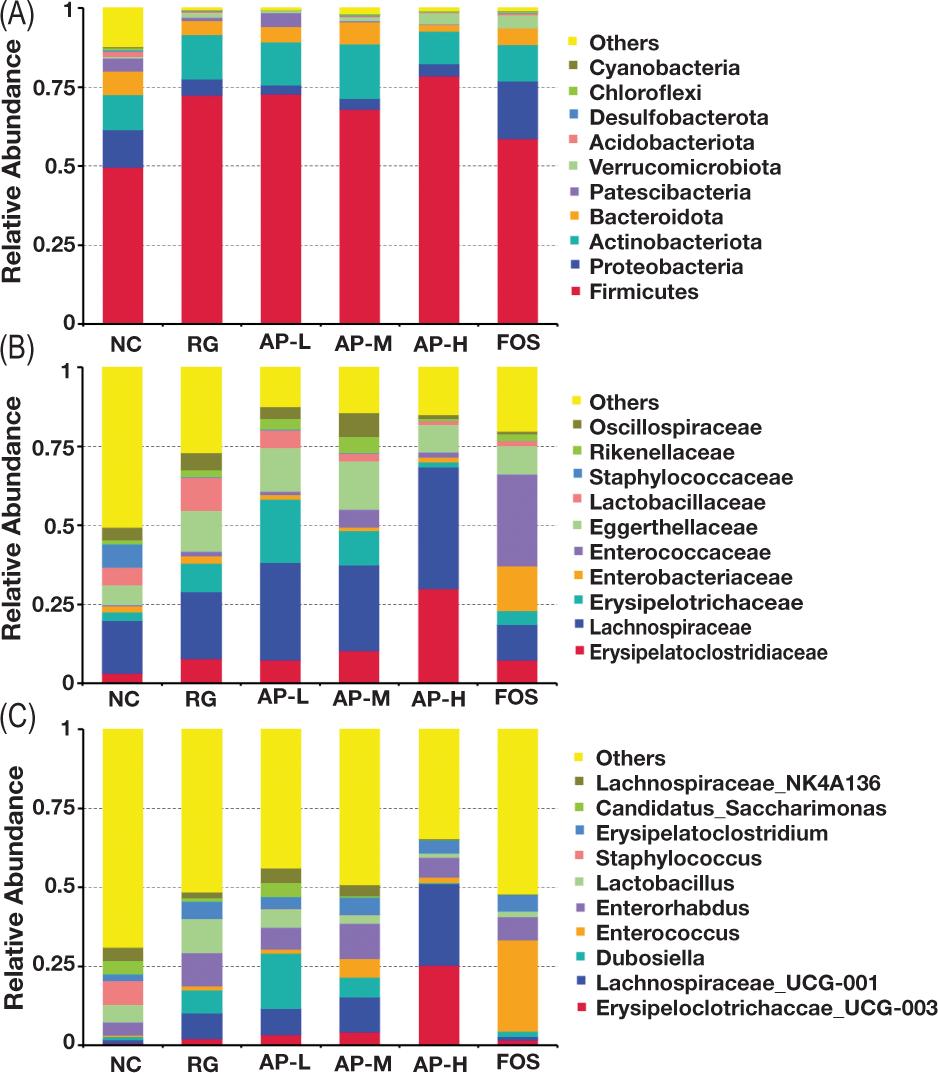
To intuitively observe species with high relative abundance at various classification levels and their proportions in intestinal floras of the different groups, column diagrams of microorganisms with the highest abundance ranking (top 10) at the phylum level (Figure 6A), family level (Figure 6B), and genus level (Figure 6C) were constructed based on the species annotation and abundance information of every sample at the genus level. The outcomes displayed that at the phylum level, Firmicutes, Proteobacteria, and Actinobacteriota were the main floras. Their proportions were approximately 90% of the total flora. Bacteroidota accounted for 5% of the total flora. At the family and genus levels, Erysipelatoclostridiaceae, Lachnospiraceae, Erysipelotrichaceae, Enterobacteriaceae, and Enterococcaceae were the main dominant floras. Their proportions were approximately 55% of the total flora. The relative abundance of Erysipelotrichaceae_UCG-003, a beneficial bacterium, was also significantly higher in the intestinal flora of the AP-H group than that of the NC group. In addition, the relative abundance of the probiotic Lachnospiraceae_UCG-001 in the intestinal flora of the AP-M and AP-H groups was also significantly higher than that of the RG group.
Figure 6. Column diagram of the relative abundance of species in intestinal flora of mice in various groups at the (A) phylum, (B) family, and (C) genus levels.
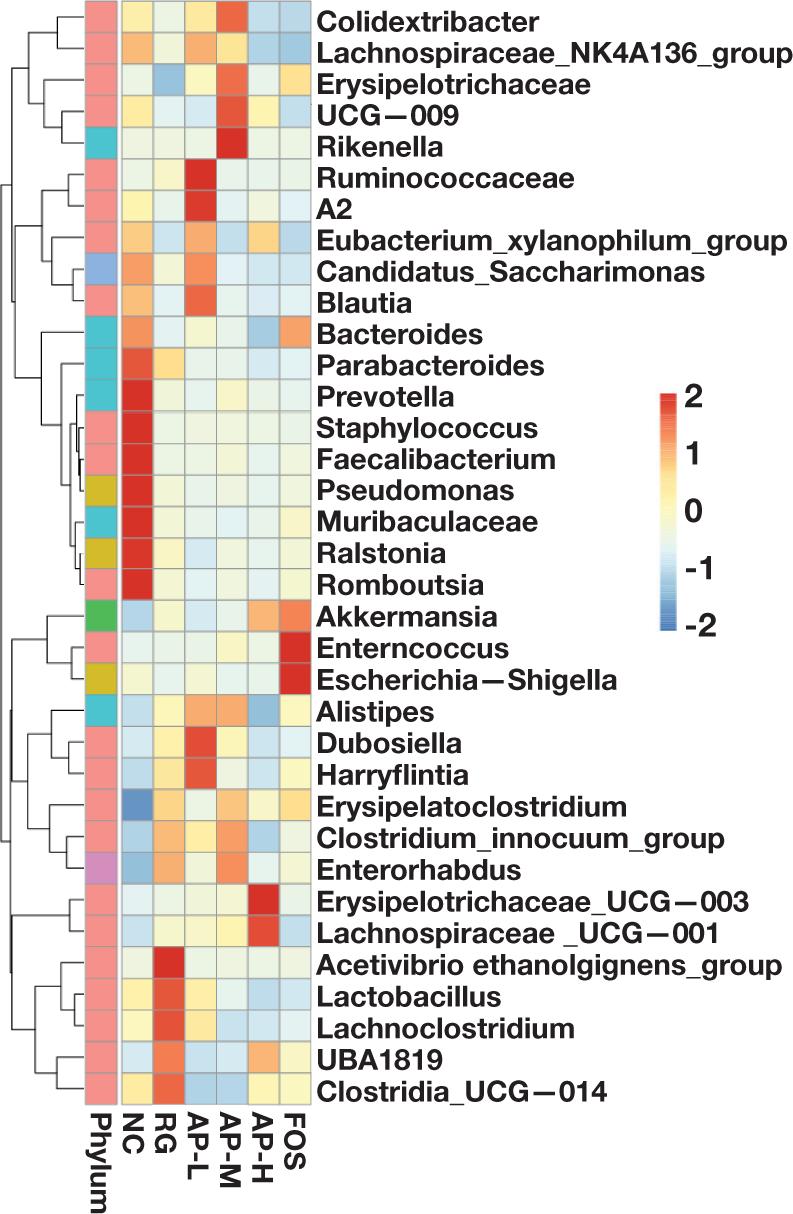
To further analyze the microbial species with great diversities in relative abundance in intestinal floras among the different groups, the genera with the relative abundance ranking (top 35) were selected for the clustering from two dimensions (species and samples) to draw a heat map based on their abundance information in the different groups (Figure 7). Results showed that by comparing with the NC and RG groups, the intragastric administration of AIPSs reversed the decrease or increase in some bacteria caused by LIH. For example, the proportions of Staphylococcus, Prevotella, Lachnospiraceae_UCG-001, Erysipelotrichaceae_UCG-003, and Dubosiella were obviously restored after the intragastric administration of AIPSs. The relative abundance of Lachnospiraceae_UCG-001 and Dubosiella in the AP-M group even significantly improved compared with those in FOS groups. It means that the role of AIPSs in improving the relative abundance of Lachnospiraceae_UCG-001 and Dubosiella was significantly better than that of FOS. Furthermore, the abundance of Acetivibrio_ethanolgignens, which easily led to inflammation or infection, significantly increased after the intragastric administration of antibiotics. Interestingly, AIPSs significantly inhibited the proliferation of Acetivibrio_ethanolgignens. As a result, the relative abundance of Bacteroides in AP-L, AP-M, and AP-H groups was even significantly lower than that in the RG group.
Figure 7. Heat map of the relative abundance of species in the intestinal flora of mice in all groups at the genus level.
Species significant-difference analysis
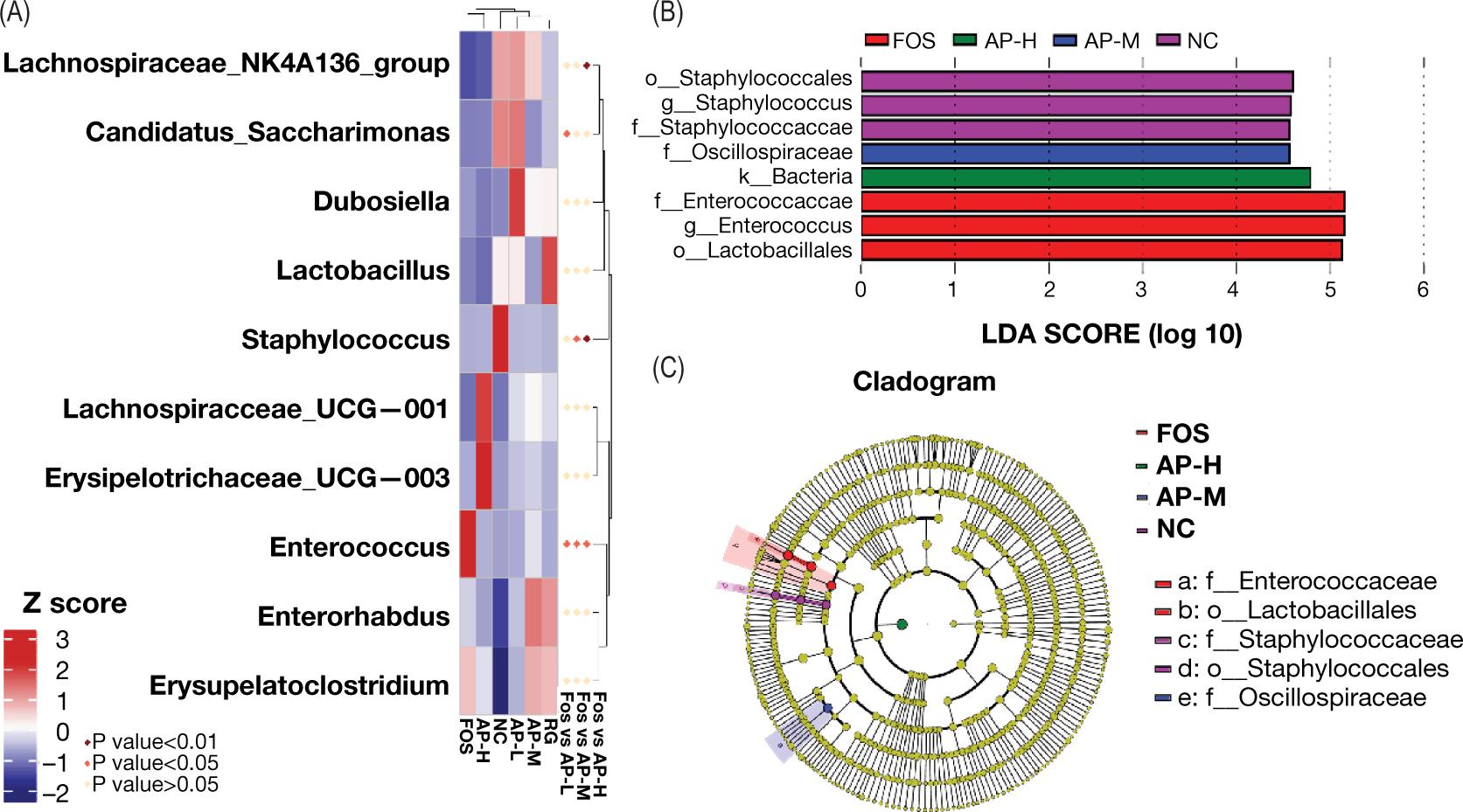
The heat map (Figure 8A) showed that the relative abundance of some bacteria were significantly different between AP-L, AP-M, AP-H, and FOS groups, such as Staphylococcus, Lachnospiraceae_NK4A136_group, Candidatus_Saccharimonas and Enterococcus. Among them, Enterococcus was reported as a potentially harmful bacterium, and the high concentration of Enterococcus has a positive correlation with rat colitis (Rose et al., 2010; Onderdonk et al., 1998). Interestingly, the relative abundance of Enterococcus in the AP-L, AP-M, and AP-H groups was even significantly lower than that in the FOS group (Figure 8A). Based on the LEfSe analysis of the AP-M, AP-H, FOS, and NC groups, eight microbial species were found with >4 LDA score, such as Staphylococcales, Oscillospiraceae, Enterococcus, and Lactobacillales (Figure 8B). Among them, Oscillospiraceae was reported to have a positive role in the extra-intestinal pain (Hollister et al., 2020). Moreover, it was reported that the inflammatory symptoms of colitis mice were reduced after taking the Bacillus cereus strain, and the abundance of Oscillospiraceae simultaneously increased significantly, which indicated that Oscillospiraceae was positively correlated with the remission of colitis and the inhibition of inflammation (Sheng et al., 2021). Additionally, LIH destroyed the balance of intestinal flora, which was primarily manifested as the reduced abundance of some beneficial bacteria and increased abundance of some pathogenic bacteria. From the evolutionary branching diagram (Figure 8C), the relative abundancs of Lactobacillales and Enterococcus in the FOS group was increased compared to the NC group. After the intragastric administration of AIPSs, the imbalance of intestinal flora structure significantly improved, further proving that AIPSs had a significant regulatory and restorative effect on the imbalance of the LIH-induced mouse intestinal flora.
Figure 8. Analysis of significant differences in intestinal flora structure among treatment groups. (A) Analysis of MetaStat heatmap. Z score represents the species aggregation degree, and the P value represents the level of significant differences. P<0.05 represents that the difference is significant, and P < 0.01 represents that the difference is extremely significant. (B) An LDA distribution histogram was used to analyze the microbial species with the significant difference among the AP-M, AP-H, FOS, and NC groups. (C) An evolutionary branching diagram to compare the AP-M, AP-H, FOS, and NC groups. The large circles from the interior to the exterior refer to classifying species from the phylum to the genus/species. The size of each node circle refers to the relative abundance of species. If the circle is larger, the relative abundance is higher.
Discussion
The intestinal flora plays an important role in maintaining body health and metabolism. However, the long-term use of broad-spectrum antibiotics has become the most common clinical cause of disorders in the intestinal flora, accompanied by varying degrees of diarrhea, depression, and inflammatory symptoms (Lv et al., 2017). Moreover, the destruction of intestinal flora mediated by antibiotics reduces the therapeutic effect of anticancer drugs (Sivan et al., 2015). The administration of prebiotics can regulate the balance of intestinal flora, enhance the gut barrier of the intestine, and reduce the risk of immune and metabolic diseases caused by taking excessive antibiotics (Varankovich et al., 2015).
Some fungal polysaccharides that cannot be digested are considered to have a good prebiotic effect. They can benefit the host by stimulating the growth or activity of specific intestinal flora (Zhang et al., 201a8). For example, Zhang et al. (Zhang et al., 2020) found that polysaccharides from Flammulina velutipes can improve colitis symptoms caused by dextran sodium sulfate (DSS) by regulating the intestinal flora to downregulate the TLR4\NF-κB inflammatory signal pathway. Ren et al. (Ren et al., 2018) found that polysaccharides from Hericium could downregulate oxidative stress and inflammatory factor markers, reverse the intestinal flora disorder of DSS inflammatory C57BL/6 in mice, and maintain the stability of the intestinal barrier. Liu et al. (Liu et al., 2020) showed that polysaccharides from Phellinus igniarius could maintain intestinal homeostasis and reduce lipopolysaccharide content in the blood by increasing the abundance of bacteria producing short-chain fatty acids (SCFAs) to reduce systemic inflammation and reverse insulin resistance. Thus, this polysaccharide has potential application as a prebiotic to regulate the intestinal flora of patients with diabetes mellitus type 2. Tian et al. (Tian et al., 2018) demonstrated that ingesting Agaricus bisporus induced a change in intestinal microflora and bacterial metabolism, and this species played the role of prebiotics and improved the development of metabolic health and diabetes. A. cinnamomea is a valuable fungus for medicine and food, and its polysaccharides have many biological activities, such as antioxidation, anti-inflammatory, antivirus, and immune-regulation activities. This species also has great research value and development potential as a prebiotic (Zhang et al., 2015; De Oliveira et al., 2017). In this study, after the intragastric administration of LIH, the body weight of mice significantly decreased, and the immune organs were damaged to varying degrees, resulting in inflammatory symptoms. However, the intragastric administration of AIPSs and FOS alleviated the weight loss and inflammatory symptoms of mice and restored the immune--organ index. This result was consistent with the report of Liu et al. (Liu et al., 2018), in which lentinan reduces the inflammatory response by reducing the production of proinflammatory factors. It was found that oligosaccharides from Antrodia cinnamomea can significantly suppress the inflammation in lung tissues of LPS-injected C57BL/6 mice (Zheng et al., 2018). Besides, Firmicutes in the intestinal bacteria participate in the nutrition and metabolism of the host by synthesizing SCFAs and exert a good curative effect on alleviating inflammatory bowel diseases (Stojanov et al., 2020). Herein, at the phylum level of microorganisms (Figure 6A), the intragastric administration of AIPSs and FOS improved the relative abundance of Firmicutes in the intestine. The column diagram of the relative abundance in genus level (Figure 6B) revealed that the middle dose of AIPSs ingestion significantly improved the proportion and relative abundance of Erysipelotrichaceae and Lachnospiraceae in the mouse intestine. Erysipelotrichaceae and Lachnospiraceae were found as the main members of butyric acid production and played an extremely important role in butyric acid production when the host participates in useful exercise (Estaki et al., 2016). However, a high abundance of Lachnospiraceae may be an antecedent of type 1 diabetes (Kameyama and Itoh, 2014). Therefore, although a high dose of AIPSs treatment also can reduce intestinal inflammation symptoms, it is obvious that the middle dose of AIPSs treatment is better for restoring the intestinal flora structure to a normal state in mice.
Compared with the FOS group, the AP-L, AP-M, and AP-H groups significantly reduced the relative abundance of Enterococcus. The immunohistochemical and serological analysis of fecal infections in pancreatic cancer patientsa and chronic pancreatitis mice revealed that high abundance of Enterococcus could cause pancreatic infection through bile (Maekawa et al., 2018). Besides, the Enterococcus also can be colonized in patients and lead to some wound infections, such as bacteremia, peritonitis, endocarditis, and urinary tract (Sava et al., 2010; O’Driscoll and Crank, 2015). It was found that Long term high-fat diet mice can reverse colonic adverse reactions after receiving a short-term balanced diet (Wu et al., 2022). A balanced diet could reduce the weight gain of mice, at the same time, the relative abundance of Lachnospiraceae_NK4A136_group also returns to the normal level, which indicates that Lachnospiraceae_NK4A136_group has the potential to become probiotics (Wu et al., 2022). Additionally, it was found that fed with fucosylated chondroitin sulfate from Acaudina molpadioides can reduce the content of serum proinflammatory cytokines (IL-6, TNF-α) in high-fat diet mice and increase the relative abundance of Lachnospiraceae_NK4A136_group in the intestinal flora (Hu et al., 2019). It is considered that the selective aggregation of probiotics, such as Lachnospiraceae_NK4A136_group and direct inhibition of bacterial pathogens lead to the reduction of inflammatory symptoms in high-fat mice. The Lachnospiraceae_NK4A136_group is beneficial to gut health and is regarded as an anti-inflammatory factor due to its production of short-chain fatty acids (Wang et al., 2019). In this study, the relative abundance of Lachnospiraceae_NK4A136_group increased significantly (P<0.01) in AP-M group compared with the FOS group (Figure 8A). Thus, we believe that the middle dose of AIPSs is well suitable for Lachnospiraceae_NK4A136_group survived in the intestinal tract of mice were fed intragastrically with lincomycin hydrochloride and finally alleviated the inflammatory symptoms. This result is consistent with the report, which proved that degraded polysaccharides from Sargassum fusiforme (PSF-T2) could reduce inflammatory factors IL-6 and TNF-α and increased the abundance of Lachnospiraceae_NK4A136_group and Oscillospiraceae in intestines, then protected the intestinal barrier and improved the colitis induced by dextran sodium sulfate in mice (Chen et al., 2021). In addition, it was found that compared with normal calf feces, the abundance of Osllospiraceae in the feces of calves with watery or hemorrhagic morphology was significantly reduced, which can be used as a potential indicator to judge the status of calf feces (Fan et al., 2021). What is more, the abundance of Osllospiraceae in normal calf feces was found significantly higher than that in watery or hemorrhagic feces (Zeineldin et al., 2018).
Therefore, we believe that the intragastric administration of AIPSs could significantly increase the relative abundance of some beneficial microorganisms in mouse intestinal flora. Such treatment could also significantly reduce the relative abundance of harmful microorganisms in mouse intestinal flora to restore its balance and even adjust and optimize its structure. These activities ease the damage and harm inflicted by antibiotics, further alleviating and treating the adverse symptoms caused by LIH, such as diarrhea, inflammation, and weight loss.
Conclusions
A. cinnamomea intracellular polysaccharides (AIPSs) were extracted from the mycelium in submerged fermentation, and the contents of neutral polysaccharides and protein in the AIPSs were 82.74%±0.63% and 1.32%±0.11%, respectively. In vivo experiments in the mice showed that the intragastric administration of AIPSs with the dose of 0.25 g/kg (bodyweight of mice) can significantly increase the relative abundance of some beneficial microorganisms in mouse intestinal flora (such as Lachnospiraceae_NK4A136_group, Osllospiraceae, Lachnospiraceae) and significantly reduced the relative abundance of some harmful microorganisms (such as Enterococcus). Consequently, the structure and balance of mouse intestinal flora were regulated, and the harm caused by antibiotics was alleviated. These actions further eased the adverse symptoms caused by LIH, such as diarrhea and inflammation. Overall, this study provided a new concept and laid a practical foundation for developing new multifunctional prebiotics.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (Grant numbers: 32001661 and 32102100), the Natural Science Foundation of Jiangsu Province, China (Grant number: BK20190890), and the Universities Natural Science Research Project of Jiangsu Province(Grant number: 19KJA430016).
REFERENCES
Chang C.J., Lu CC., Lin C.S., Martel J., Ko Y.F., Ojcius D.M., Wu T.R., Tsai Y.H., Yeh T.S., and Lu J.J,, 2018. Antrodia cinnamomea reduces obesity and modulates the gut microbiota in high-fat diet-fed mice. International Journal of Obesity, 42(2): 231–243. 10.1038/ijo.2017.149
Chen F and Huang GL, 2018. Preparation and immunological activity of polysaccharides and their derivatives. International Journal of Biological Macromolecules, 112: 211–216. 10.1016/j.ijbiomac.2018.01.169
Chen Q.L., Tang H.L., Zha Z.Q., Yin H.P., Wang Y., Wang Y.F., Li H.T., and Yue L., 2017. β-d-glucan from Antrodia Camphorata ameliorates LPS-induced inflammation and ROS production in human hepatocytes. International Journal of Biological Macromolecules, 104: 768–777. 10.1016/j.ijbiomac.2017.05.191
Chen X.Y., Li X., Sun-Waterhouse D.X., Zhu B.Y., You L.J., and Hileuskaya K., 2021. Polysaccharides from Sargassum fusiforme after UV/H2O2 degradation effectively ameliorate dextran sulfate sodium-induced colitis. Food & Function, 12(23): 11747–11759. 10.1039/D1FO02708E
De Oliveira A.F., Do Nascimento G.E., Lacomini M., Cordeiro L.M.C., and Cipriani T.R., 2017. Chemical structure and anti-inflammatory effect of polysaccharides obtained from infusion of Sedum dendroideum leaves. International Journal of Biological Macromolecules, 105: 940–946. 10.1016/j.ijbiomac.2017.07.122
Estaki M., Pither J., Baumeister P., Little J.P., Gill, S.K., Ghosh S., Ahmadi-Vand Z., Marsden K.R., and Gibson D.L., 2016. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome, 4(1): 1–13. 10.1186/s40168-016-0189-7
Everard A., Belzer C., Geurts L., Ouwerkerk J.P., Druart C., Bindels L.B., Guiot Y., Derrien M., Muccioli G.G., and Delzenne N.M., 2013. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proceedings of the National Academy of Sciences, 110(22): 9066–9071. 10.1073/pnas.1219451110
Fan P.X., Kim M., Liu G., Zhai Y.T., Liu T., Driver J.D., and Jeong K.C., 2021. The gut microbiota of newborn calves and influence of potential probiotics on reducing diarrheic disease by inhibition of pathogen colonization. Frontiers in Microbiology, 12: 772863. 10.3389/fmicb.2021.772863
Ganesan N., Baskaran R., Velmurugan B.K., and Thanh N.C., 2019. Antrodia cinnamomea—An updated minireview of its bioactive components and biological activity. Journal of Food Biochemistry, 43(8): e12936. 10.1111/jfbc.12936
Gao P.F., Ma C., Sun Z., Wang L.F., Huang S., Su X.Q., Xu J., and Zhang H.P., 2017. Feed-additive probiotics accelerate yet antibiotics delay intestinal microbiota maturation in broiler chicken. Microbiome, 5(1): 1–14. 10.1186/s40168-017-0315-1
Geethangili M. and Tzeng Y.M., 2011. Review of pharmacological effects of Antrodia camphorata and its bioactive compounds. Evidence-Based Complementary and Alternative Medicine, 2011. 10.1093/ecam/nep108
Hollister E.B., Cain K.C., Shulman R.J., Jarrett M.E., Burr R.L., Ko C., Zia J., Han C.J., and Heitkemper M.M., 2020. Relationships of microbiome markers with extra-intestinal, psychological distress and gastrointestinal symptoms, and quality of life in women with irritable bowel syndrome. Journal of Clinical Gastroenterology, 54(2): 175. 10.1097/MCG.0000000000001107
Hooper L.V. and Macpherson A.J., 2010. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nature Reviews Immunology, 10(3): 159–169. 10.1038/nri2710
Hu S.W., Wang J.H., Xu Y.L., Yang H.C., Wang J.F., Xue C.H., Yan X.J., and Su L.J., 2019. Anti-inflammation effects of fucosylated chondroitin sulphate from Acaudina molpadioides by altering gut microbiota in obese mice. Food & Function, 10(3): 1736–1746. 10.1039/c8fo02364f
Kameyama K. and Itoh K., 2014. Intestinal colonization by a Lachnospiraceae bacterium contributes to the development of diabetes in obese mice. Microbes and Environments, 29(4): 427–430. 10.1264/jsme2.ME14054
Ker Y.B., Peng C.C., Chang W.L., Chyau C.C., and Peng R.Y., 2014. Hepatoprotective bioactivity of the glycoprotein, antrodan, isolated from Antrodia cinnamomea mycelia. PLoS One, 9(4): e93191. 10.1371/journal.pone.0093191
Li HX, Lu Z.M., Geng Y., Gong J.S., Zhang X.J., Shi J.S., Xu Z.H., and Ma Y.H., 2015. Efficient production of bioactive metabolites from Antrodia camphorata ATCC 200183 by asexual reproduction-based repeated batch fermentation. Bioresource Technology, 194: 334–343. 10.1016/j.biortech.2015.06.144
Liang J.J., Zhang M.N., Wang X.N., Ren Y.C., Yue T.L., Wang Z.L., and Gao Z.P., 2021. Edible fungal polysaccharides, the gut microbiota, and host health. Carbohydrate Polymers, 273: 118558. 10.1016/j.carbpol.2021.118558
Liu K.J., Leu S.J., Su C.H., Chiang B.L., Chen Y.L., and Lee Y.L., 2010. Administration of polysaccharides from Antrodia camphorata modulates dendritic cell function and alleviates allergen-induced T helper type 2 responses in a mouse model of asthma. Immunology, 129(3): 351-362. 10.1111/j.1365-2567.2009.03175.x
Liu L., Rohdin P., and Moshfegh B., 2018. Investigating cost--optimal refurbishment strategies for the medieval district of Visby in Sweden. Energy and Buildings, 158: 750–760. 10.1016/j.enbuild.2017.10.002
Liu Y.G., Li L.Z., An S.S., Zhang Y.Z., Feng S.W., Zhao L., Teng L.R., and Wang D., 2017a. Antifatigue effects of Antrodia cinnamomea cultured mycelium via modulation of oxidative stress signaling in a mouse model. BioMed Research International, 2017: 9374026. 10.1155/2017/9374026
Liu Y.G., Wang J., Li L.Z., Hu W.J., Qu Y.D., Ding Y.P., Meng L.N., Teng L.R., and Wang D., 2017b. Hepatoprotective effects of Antrodia cinnamomea: The modulation of oxidative stress signaling in a mouse model of alcohol-induced acute liver injury. Oxidative Medicine and Cellular Longevity, 2017: 7841823. 10.1155/2017/7841823
Liu Y.Y., Wang C.R., Li J.S., Li T.T., Zhang Y., Liang Y.X., and Mei Y.X., 2020. Phellinus linteus polysaccharide extract improves insulin resistance by regulating gut microbiota composition. The FASEB Journal, 34(1): 1065–1078. 10.1096/fj.201901943RR
Lu M.C., El Shazly M., Wu T.Y., Du Y.C., Chang T.T., Chen C.F., Hsu Y.M., Lai K.H., Chiu C.P., and Chang F.R., 2013. Recent research and development of Antrodia cinnamomea. Pharmacology & Therapeutics, 139(2): 124–156. 10.1016/j.pharmthera.2013.04.001
Lv W.J., Liu C., Ye C.X., Sun J.Q., Tan X.W., Zhang C., Qu Q., Shi D.Y., and Guo S.N., 2017. Structural modulation of gut microbiota during alleviation of antibiotic-associated diarrhea with herbal formula. International Journal of Biological Macromolecules, 105(3): 1622–1629. 10.1016/j.ijbiomac.2017.02.060
Maekawa T., Fukaya R., Takamatsu S., Itoyama S., Fukuoka T., Yamada M., Hata T., Nagaoka S., Kawamoto K., and Eguchi H., 2018. Possible involvement of Enterococcus infection in the pathogenesis of chronic pancreatitis and cancer. Biochemical and Biophysical Research Communications, 506(4): 962–969. 10.1016/j.bbrc.2018.10.169
Masuko T., Minami A., Iwasaki N., Majima T., Nishimura S.I., and Lee Y.C., 2005. Carbohydrate analysis by a phenol–sulfuric acid method in microplate format. Analytical Biochemistry, 339(1): 69–72. 10.1016/j.ab.2004.12.001
O’Driscoll T. and Crank C.W., 2015. Vancomycin-resistant enterococcal infections: epidemiology, clinical manifestations, and optimal management. Infection and Drug Resistance, 8: 217–230. 10.2147/IDR.S54125
Onderdonk A.B., Richardson J.A., Hammer R.E., and Taurog J.D., 1998. Correlation of cecal microflora of HLA-B27 transgenic rats with inflammatory bowel disease. Infection and Immunity, 66(12): 6022–6023. 10.1128/IAI.66.12.6022-6023.1998
Pelaseyed T. and Hansson G.C., 2020. Membrane mucins of the intestine at a glance. Journal of Cell Science, 133(5): jcs240929. 10.1242/jcs.240929
Rajakovich L.J. and Balskus E.P., 2019. Metabolic functions of the human gut microbiota: the role of metalloenzymes. Natural Product Reports, 36(4): 593–625. 10.1039/C8NP00074C
Ren Y.L., Geng Y., Du Y., Li W., Lu Z.M., Xu H.Y., Xu G.H., Shi J.S., and Xu Z.H., 2018. Polysaccharide of Hericium erinaceus attenuates colitis in C57BL/6 mice via regulation of oxidative stress, inflammation-related signaling pathways and modulating the composition of the gut microbiota. The Journal of Nutritional Biochemistry, 57: 67–76. 10.1016/j.jnutbio.2018.03.005
Rose D.J., Venema K., Keshavarzian A., and Hamaker B.R., 2010. Starch-entrapped microspheres show a beneficial fermentation profile and decrease in potentially harmful bacteria during in vitro fermentation in faecal microbiota obtained from patients with inflammatory bowel disease. British Journal of Nutrition, 103(10): 1514–1524. 10.1017/S0007114509993515
Sava I.G., Heikens E., and Huebner J., 2010. Pathogenesis and immunity in enterococcal infections. Clinical Microbiology and Infection, 16(6): 533–540. 10.1111/j.1469-0691.2010.03213.x
Sheng K.L., Xu Y.F., Kong X.W., Wang J.M., Zha X.D., and Wang Y.Z., 2021. Probiotic Bacillus cereus Alleviates dextran sulfate sodium-induced colitis in mice through improvement of the intestinal barrier function, anti-inflammation, and gut microbiota modulation. Journal of Agricultural and Food Chemistry, 69(49): 14810–14823. 10.1021/acs.jafc.1c03375
Sivan A., Corrales L., Hubert N., Williams J.B., Aquino Michaels K., Earley Z.M., Benyamin F.W., Lei Y.M., Jabri B., and Alegre M.L., 2015. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science, 350(6264): 1084–1089. 10.1126/science.aac4255
Song T.Y., Hsu S.L., Yeh C.T., and Yen G.C., 2005. Mycelia from Antrodia camphorata in submerged culture induce apoptosis of human hepatoma HepG2 cells possibly through regulation of Fas pathway. Journal of Agricultural and Food Chemistry, 53(14): 5559–5564. 10.1021/jf050329
Stojanov S., Berlec, A., and Štrukelj B., 2020. The influence of probiotics on the Firmicutes/Bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms, 8(11): 1715. 10.3390/microorganisms8111715
Tian Y., Nichols R.G., Roy P., Gui W., Smith P.B., Zhang J.T., Lin Y.D., Weaver V., Cai J.W., and Patterson A.D., 2018. Prebiotic effects of white button mushroom (Agaricus bisporus) feeding on succinate and intestinal gluconeogenesis in C57BL/6 mice. Journal of Functional Foods, 45: 223–232. 10.1016/j.jff.2018.04.008
Varankovich N.V., Nickerson M.T., and Korber D.R., 2015. Probiotic-based strategies for therapeutic and prophylactic use against multiple gastrointestinal diseases. Frontiers in Microbiology, 6: 685. 10.3389/fmicb.2015.00685
Wang C.S., Li W.B., Wang H.Y., Ma, Y.M., Zhao X.H., Zhang X.D., Yang H., Qian J.M., and Li J.N., 2019. Saccharomyces boulardii alleviates ulcerative colitis carcinogenesis in mice by reducing TNF-α and IL-6 levels and functions and by rebalancing intestinal microbiota. BMC Microbiology, 19(1): 1–12. 10.1186/s12866-019-1610-8
Wu M.R., Chou T.S., Huang C.Y., and Hsiao J.K., 2020. A potential probiotic-Lachnospiraceae NK4A136 group: Evidence from the restoration of the dietary pattern from a high-fat diet. Research Square, 2020. 10.21203/rs.3.rs-48913/v1
Wu T.R., Lin C.S., Chang C.J., Lin T.L., Martel J., Ko Y.F., Ojcius D.M., Lu C.C., Young J.D., and Lai H.C., 2019. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut, 68(2): 248–262. 10.1136/gutjnl-2017-315458
Yang C.M., Zhou Y.J, Wang R.J., and Hu M.L., 2009. Anti-angiogenic effects and mechanisms of polysaccharides from Antrodia cinnamomea with different molecular weights. Journal of Ethnopharmacology, 123(3): 407–412. 10.1016/j.jep.2009.03.034
Yang H.L., Lin S.W., Lee C.C., Lin K.Y., Liao C.H., Yang T.Y., Wang H.M., Huang H.C., Wu C.R., and Hseu Y.C., 2015. Induction of Nrf2-mediated genes by Antrodia salmonea inhibits ROS generation and inflammatory effects in-lipopolysaccharide-stimulated RAW264. 7 macrophages. Food & Function, 6(1): 229–240. 10.1039/C4FO00869C
Yen I.C., Yao C.W., Kuo M.T., Chao C.L., Pai C.Y., and Chang W.L., 2015. Anti-cancer agents derived from solid-state fermented Antrodia camphorata mycelium. Fitoterapia, 102: 115–119. 10.1016/j.fitote.2015.02.010
Zeineldin M., Aldridge B., and Lowe J., 2018. Dysbiosis of the fecal microbiota in feedlot cattle with hemorrhagic diarrhea. Microbial Pathogenesis, 115: 123–130. 10.1016/j.micpath.2017.12.059
Zhang R.J., Yuan S.J., Ye J.F., Wang X.D., Zhang X.D., Shen J., Yuan M.M., and Liao W.Z., 2020. Polysaccharide from Flammuliana velutipes improves colitis via regulation of colonic microbial dysbiosis and inflammatory responses. International Journal of Biological Macromolecules, 149: 1252–1261. 10.1016/j.ijbiomac.2020.02.044
Zhang W.X., Song D., Xu D., Wang T.T., Chen L., and Duan J.Y., 2015. Characterization of polysaccharides with antioxidant and immunological activities from Rhizoma Acori Tatarinowii. Carbohydrate Polymers, 133: 154–162. 10.1016/j.carbpol.2015.07.018
Zhang X., Zhang M., Ho C.T., Guo X.J., Wu Z.F., Weng P.F., Yan M.D., and Cao J.X., 2018a. Metagenomics analysis of gut microbiota modulatory effect of green tea polyphenols by high fat diet-induced obesity mice model. Journal of Functional Foods, 46: 268–277. 10.1016/j.jff.2018.05.003
Zhang Y.T., Wang Z., Li D.Y., Zang W.T., Zhu H., Wu P.Y., Mei Y.X., and Liang Y.X., 2018b. A polysaccharide from Antrodia cinnamomea mycelia exerts antitumor activity through blocking of TOP1/TDP1-mediated DNA repair pathway. International Journal of Biological Macromolecules, 120: 1551–1560. 10.1016/j.ijbiomac.2018.09.162
Zheng J.P., Jiao S.M., Li Q.Y., Jia P.Y., Yin H., Zhao X.M., Du Y.G., and Liu H.T., 2018. Antrodia cinnamomea oligosaccharides suppress lipopolysaccharide-induced inflammation through promoting O-GlcNAcylation and repressing p38/Akt phosphorylation. Molecules, 23(1): 51. 10.3390/molecules23010051