
Download



RESEARCH ARTICLE
Biofilm formation, antibiotic resistance, and genome sequencing of a unique isolate Salmonella Typhimurium M3
Lei Yuan1,2, Yang Liu1, Luyao Fan1, Caowei Chen1, Zhenquan Yang1*, Xin-an Jiao2*
1College of Food Science and Engineering, Yangzhou University, Yangzhou, PR China;
2Jiangsu Key Laboratory of Zoonoses, Yangzhou, PR China
Abstract
Salmonella Typhimurium is a zoonotic bacterium that can cause salmonellosis, and the major concerns of S. Typhimurium for the food industry are its ability to obtain multidrug resistance and form biofilms on food--contact surfaces. In the current study, the antimicrobial resistance of a strong biofilm former S. Typhimurium M3 was assessed by the diffusion method. Genome sequencing was also applied to obtain the genes related to antibiotic resistance, and biofilm formation of S. Typhimurium M3. Biofilm-forming capacity of S. Typhimurium M3 was found to be strain dependent, and a high number of isolates were strong biofilm formers. The high biofilm--forming isolate S. Typhimurium M3 was resistant to oxacillin, lincomycin, rifampicin, tetracycline, and clindamycin, with the MIC values of 512 μg/mL, 32 μg/mL, 16 μg/mL, 64 μg/mL, and 64 μg/mL, respectively. Genomic annotation of S. Typhimurium M3 showed the presence of genes involved in cellulose biosynthesis, curli production, fimbriae biosynthesis, flagellar assemble, quorum sensing, chemotaxis, and some transcriptional regulators. Antibiotic efflux conferring antibiotic resistance genes, antibiotic inactivation genes, and antibiotic target alteration genes were also identified. The results expand scientific understanding on how Salmonella isolates with high biofilm-forming capacity and multidrug resistance survive in stressful conditions in the industry.
Key words: biofilm, genome sequencing, multidrug resistance, Salmonella Typhimurium
*Corresponding authors: Zhenquan Yang, Yangzhou University, 196 Huayang West Road, Yangzhou, Jiangsu 225127, PR China. Email: [email protected]; Xinan Jiao, Jiangsu Key Laboratory of Zoonoses, Yangzhou, Jiangsu, 225009, PR China. Email: [email protected]
Received: 19 October 2022; Accepted: 4 January 2023; Published: 13 January 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Salmonella Typhimurium is one of the major zoonotic bacteria that can cause serious gastroenteritis, typhoid fever, diarrhea, and sepsis (Taylor and Winter, 2020). This bacterium is usually transmitted to humans via the consumption of poultry products, or sometimes by contaminated surfaces (Shen et al., 2022). Although Chinese official data for Salmonella contamination in foods is not available, many studies have indicated that the prevalence of Salmonella was found to be 10.7–71.8%, with S. Typhimurium being one of the most predominant serovars (Zhou et al., 2018). In addition, studies have declared that the major concerns of S. Typhimurium in the food industry are the ability to acquire antimicrobial resistance and form biofilms on surfaces (Merino et al., 2019; Wang et al., 2022).
Antibiotics are used to both control the animal infections caused by Salmonella, and alleviate the widespread of salmonellosis among humans. Unfortunately, with the overuse of antibiotics in agriculture, S. Typhimurium is becoming highly resistant to a variety of antibiotics including lincomycin, tetracycline, quinolones, sulfamethoxazole, and oxytetracycline (Ingle et al., 2021; Tian et al., 2021). The occurrence of multidrug-resistant S. Typhimurium could cause treatment failure in animal husbandry and constitute huge threats to public health.
Biofilm formation has been proved to be a key factor that contributes to the ineffectiveness of antibiotics, which allows for the persistence of S. Typhimurium in the food industry (Yuan et al., 2021). Defined as the assemblage of microbes within the self-produced extracellular polymeric substances (EPS), biofilms are a regular mode for bacterial growth in nature, which account for around 80% of bacterial infections globally (Yuan et al., 2020). Many studies have indicated the occurrence of S. Typhimurium on meat products, glasses, plastics, stainless steel, and rubber in poultry processing environments, and the -presence in the above environs is strongly linked to its biofilm-forming capacity (Lee et al., 2020). Once formed, S. Typhimurium biofilms are believed to show much higher resistance to antibiotics compared to their free-living counterparts, leading to the continuous transmission of foodborne diseases in the food industry (Yuan et al., 2020).
In this study, the biofilm-forming capacity and antibiotic resistance profiles of S. Typhimurium isolates from a swine slaughterhouse were evaluated. In addition, genome sequencing was performed to determine the genes of a unique S. Typhimurium isolate with multi-characteristics of high biofilm-forming ability and multidrug resistance. The results could provide new insights into the persistence of S. Typhimurium in the food industry, which would be valuable to effectively control Salmonella contamination in the food industry.
Materials and Methods
Bacterial strain and culture conditions
In this study, 39 S. Typhimurium isolates were isolated from a swine slaughterhouse in Jiangsu, China, from October 2016 to April 2017 (Zhou et al., 2018). The cultures of S. Typhimurium were stored at -80°C in tryptic soy broth (TSB, Difco, USA), which contained 20% (v/v) glycerol.
Biofilm formation by S. Typhimurium
Biofilm formation by S. Typhimurium was determined according to the crystal violet staining assay described by Yuan et al. (2018). In brief, aliquots of 200 μL bacterial culture at a concentration of 104 CFU/mL in TSB were added into six wells of a 96-well plate (Costar, Corning, USA). Six negative controls with only TSB were also included. After incubation for 24 h at 37°C, the medium was gently poured, and each well was washed with sterile phosphate buffer saline (PBS), followed by fixing with methanol. Each well was stained by 200 μL of 0.05% (w/v) crystal violet solution (Sigma, USA) for 10 min, and washed with PBS. Adhered crystal violet in each well was then solubilized in 200 μL of 33% (v/v) acetic acid. The microplate reader (Thermo Fisher, USA) was used to measure optical density (OD) of the solution in wells at 594 nm. Biofilm-forming ability of each S. Typhimurium isolate was expressed by cut-off OD (ODc), which was defined as the mean OD of negative control plus three standard deviations (SD). Biofilm-forming capacity of each S. Typhimurium isolate was classified as the following groups: strong biofilm former (OD > 4ODc), moderate biofilm former (4ODc ≥ OD > 2ODc), weak biofilm former (2ODc ≥ OD > ODc), and no biofilm former (ODc ≥ OD) (Diaz et al., 2016). All results are expressed as mean ± SD obtained from assays with three replicates.
Biofilm formation of S. Typhimurium isolates on stainless steel surface was measured by following the assay as described by Yuan et al. (2018). Stainless steel coupons (with a square size of 1 × 1 cm) were immersed in acetone, washed with 75% (v/v) ethanol, rinsed with water, and then autoclaved before use. The overnight culture of each S. Typhimurium isolate was inoculated in 100 mL TSB in a flask containing coupons to obtain the final bacterial concentration of 104 CFU/mL. After the incubation at 37°C for 24 h, stainless steel coupons were removed, washed with sterile PBS (pH 7), and transferred to a tube containing 10 g of glass beads and 10 mL of sterile PBS (pH 7). Adhered cells were detached from stainless steel surface by vortex mixing for 120 s. Each bacterial suspension was decimally diluted and plated onto TSA plates, followed by the incubation for 48 h at 28°C. All results are expressed as mean ± SD obtained from assays with three replicates.
Antimicrobial resistance of the high-biofilm former S. Typhimurium M3
Antibiotic resistance of the high-biofilm former S. Typhimurium M3 was assessed based on the diffusion method of Clinical Laboratory Standards Institute (CLSI) guidelines (2016). Commercial antibiotic discs (HiMedia, India) contain 22 antibiotics (Table 1). In brief, 100 μL of overnight S. Typhimurium M3 culture was spread onto TSA plates. Then, antibiotic discs were placed on plates, followed by the incubation for 20 h at 37°C. Inhibition zones (mm) were measured and categorized as susceptible (S), intermediate (IR), and resistant (R) according to each zone diameter of ≥20, 15–19, and ≤14 mm, respectively.
Table 1. Antibiotic susceptibility of Salmonella Typhimurium M3.
| Antibiotics | Antimicrobial resistance | Antibiotics | Antimicrobial resistance |
|---|---|---|---|
| Oxacillin (1 μg) | R | Doxycycline (30 μg) | I |
| Amoxicillin (20 μg) | S | Enrofloxacin (10 μg) | S |
| Gentamicin (10 μg) | I | Azithromycin (15 μg) | I |
| Streptomycin (10 μg) | I | Norfloxacin (10 μg) | S |
| Penicillin (10 μg) | S | Lomefloxacin (10 μg) | S |
| Cefotaxime (30 μg) | S | Nitrofurantoin (300 μg) | S |
| Cotrimoxazole (1.25 μg) | S | Ampicillin (10 μg) | S |
| Polymyxin B (10 μg) | I | Rifampicin (5 μg) | R |
| Amikacin (30 μg) | S | Clindamycin (10 μg) | R |
| Novobiocin (10 μg) | I | Erythromycin (15 μg) | I |
| Lincomycin (10 μg) | R | Tetracycline (30 μg) | R |
R, Resistant (≤14 mm); IR, Intermediate resistant (15–19 mm); S, Susceptible (≥20 mm).
Defined as the lowest concentration of antibiotics that can inhibit bacterial growth, the MIC of antibiotics in S. Typhimurium M3 was assessed by the broth microdilution method (Dawan and Ahn, 2020). In brief, different dilutions of antibiotics and S. Typhimurium M3 were inoculated in 96-well plates at 37°C for 24 h.
Genome sequencing of S. Typhimurium M3
Genomic DNA of S. Typhimurium M3 was extracted by E.Z.N.A.® Bacterial DNA Kit (Omega, USA) according to the instructions. DNA quality was assessed by NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, USA). Preparation for library was performed based on Illumina’s TruSeq Nano DNA Sample Prep Kit. Whole genome of S. Typhimurium M3 was sequenced by Illumina novaseq 6000 (Illumina Inc, San Diego, USA). Raw paired-end reads were trimmed and filtered using Trimmomatic, and sequences were assembled by ABySS. Genes in the genome of S. Typhimurium M3 were predicted with GeneMarkS. All gene models were blastp against nonredundant (NR) database, Kyoto Encyclopedia of Genes and Genomes (KEGG), Gene Ontology (GO), and Cluster of Orthologous Group (COG) for functional annotation. A circular map of S. Typhimurium M3 genome was drawn using Circos. Antibiotic resistance genes in the S. Typhimurium M3 genome were predicted by Resistance Gene Identifier against the Comprehensive Antibiotic Resistance Database (CARD).
Results and Discussion
Biofilm formation by S. Typhimurium isolates
Biofilm formation by S. Typhimurium in food industry may cause huge economic losses and serious safety issues globally. In this study, the biofilm--forming capacity of each S. Typhimurium isolate was quantified by the crystal violet staining assay. The OD594 values of S. Typhimurium isolates ranged from 0.017 to 1.098, and the highest OD594 value was obtained from S. Typhimurium M3. Biofilm-forming ability was proved to be in a strain--dependent way, as 9, 12, and 18 isolates were classified as weak, moderate, and strong biofilm formers, respectively (Figure 1A). High biofilm-forming ability of S. Typhimurium was also observed in previous studies, providing them survival strategies against antibiotics (Lee et al., 2020). Characteristics of different food-contact surfaces could influence bacterial attachment and biofilm formation of microorganisms. In this work, isolates belonging to S. Typhimurium were also measured for their biofilm--forming capacity on stainless steel surface to simulate the real situation in the food industry. Cell counts recovered from biofilms of all S. Typhimurium isolates were in a range from 2.96 to 8.06 log CFU/cm2 (Figure 1B). High biofilm-forming capacity (7.95 log CFU/cm2) of S. Typhimurium M3 on stainless steel coupon was also found. A previous study proved that Salmonella could be transferred from biofilms formed on stainless steel coupons to poultry products, posing safety issues to consumers (Wang et al., 2015). Therefore, the high biofilm-forming ability of S. Typhimurium observed in this study indicates the need to optimize the cleaning and disinfection procedures to avoid Salmonella biofilm contamination.
Figure 1. Biofilm formation of 39 Salmonella Typhimurium isolates on both polystyrene (A) and stainless steel coupons (B).
Antibiotic resistance of S. Typhimurium M3
The wide spread of multidrug-resistant Salmonella has made it a threat to public health. The high biofilm--forming isolate S. Typhimurium M3 was selected to test its susceptibility to 22 antibiotics by the diffusion method (Table 1). S. Typhimurium M3 was resistant to oxacillin, lincomycin, rifampicin, tetracycline, and clindamycin, and intermediate resistant to gentamicin, streptomycin, polymyxin B, novobiocin, doxycycline, azithromycin, and erythromycin. Furthermore, the MICs of oxacillin, lincomycin, rifampicin, tetracycline, and clindamycin were 512, 32, 16, 64, and 64 ug/mL, respectively. Occurrence of resistance to these antibiotics of S. Typhimurium have already been proved in different countries (Dawan and Ahn, 2020; Harb et al., 2018). It is speculated that the resistance to multiple antibiotics may be caused by several mechanisms, including altering the cell membrane permeability, modifying the site of drug action, interfering with DNA synthesis, and affecting the structure of cell wall (Enrique et al., 2020).
Genome properties of S. Typhimurium M3
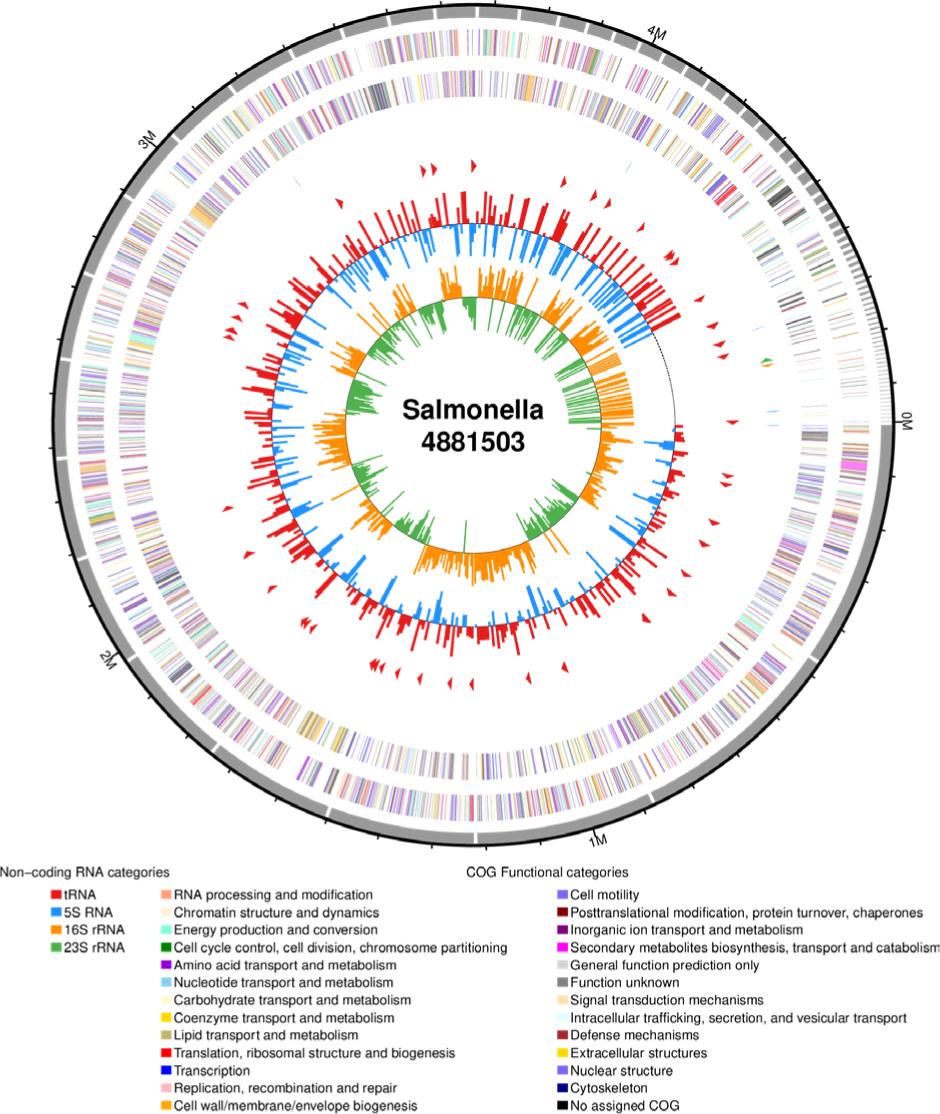
This work also aims to determine the genome properties of S. Typhimurium M3 with unique multi--characteristics of high biofilm-forming and multidrug resistance. The draft genome sequence of S. Typhimurium M3 had a length of 4881503 bp, a GC content of 52.17%, which contained 4579 coding sequences, 81 tRNAs, 8 rRNAs (6 of 5S rRNA, 1 of 16S rRNA, and 1 of 23S rRNA) (Table 2). The circular representation of S. Typhimurium M3 draft genome is shown in Figure 2.
Table 2. Genome properties of Salmonella Typhimurium M3.
| Attributes | Salmonella Typhimurium M3 |
|---|---|
| Genome size | 4881503 bp |
| Protein coding genes | 4579 |
| G-C content | 52.17% |
| rRNA encoding genes | 8 |
| tRNA encoding genes | 81 |
| Total scaffolds | 80 |
| Scaffold N50 | 208949 bp |
| Scaffold N90 | 47339 bp |
Figure 2. Genomic circle map of Salmonella Typhimurium M3.
The proteins with functional assignments of S. Typhimurium M3 included 2276 with GO assignments, 3882 with eggNOG, 138 with CARD, and 3171 with KEGG pathway mapping. The COG protein database was generated by the comparison of predicted and known proteins in completely sequenced microbial genomes to infer sets of orthologs (Figure 3). In this study, the COG annotation showed that transcription, carbohydrate transport and metabolism, energy production and conversion, and amino acid transport and metabolism were the most abundant categories. Furthermore, KEGG annotation showed that 2003, 485, 263, and 245 genes were participated in overview maps, carbohydrate metabolism, amino acid metabolism, and membrane transport, respectively (Figure 4).
Figure 3. Cluster of orthologous group annotation of genes in Salmonella Typhimurium M3.
Figure 4. Kyoto encyclopedia of genes and genomes pathway enrichment results of genes in Salmonella Typhimurium M3.
The genome analysis of S. Typhimurium M3 showed several genes related to antibiotic resistance by antibiotic efflux, antibiotic target alteration, and antibiotic inactivation (Table 3). Antimicrobial resistance genes were predicted based on the CARD database, and S. Typhimurium M3 contained 18 genes connected with resistance to tetracycline (tetA), fluoroquinolone (gyrB, parC, parE), triclosan (gyrA), sulfonamide (folP), nitrofuran (nfsA), elfamycin (tuf), aminoglycoside (AAC6), rifamycin (rpoB), fosfomycin (uhpA, uhpT, ptsI, murA, cyaA, glpT), cephalosporin (ampH), and isoniazid (fabI). Remarkably, in this study, 36 genes belonged to several gene families, including ATP-binding cassette (ABC) antibiotic efflux pumps (soxS, soxR, phoP, msbA, tolC, yojI), resistance-nodulation–cell division (RND) antibiotic efflux pumps (acrA, acrE, acrR, baeR, baeS, cpxA, golS, marR, mdsA, mdsB, mdsC, sdiA, crp, marA, acrB, acrD, acrF, mdtA, mdtB, mdtC, ramA), major facilitator superfamily (emrA, emrB, emrR, hns, mdtG, mdtH, mdtM), multidrug and toxic compound extrusion (MATE) transporters (mdtK), and KdpDE (kdpE) identified in the genome of S. Typhimurium M3. The presence of efflux pumps makes S. Typhimurium isolates insensitive to tetracycline, oxacillin, lincomycin, rifampicin, streptomycin, clindamycin, kanamycin, ciprofloxacin, norfloxacin (Ge et al., 2022; Zwama and Nishino, 2021).
Table 3. Genes essential for antibiotic resistance in genome of Salmonella Typhimurium M3.
| Gene identified | General function | Resistance mechanism |
|---|---|---|
| aac6-I | Cryptic aminoglycoside N-acetyltransferase AAC(6')-Iy/Iaa | Antibiotic inactivation |
| acrA | Multidrug efflux RND transporter periplasmic adaptor subunit AcrA | Antibiotic efflux |
| acrB | Multidrug efflux pump subunit AcrB | Antibiotic efflux |
| acrD | Multidrug efflux RND transporter permease AcrD | Antibiotic efflux |
| acrE | Efflux RND transporter periplasmic adaptor subunit | Antibiotic efflux |
| acrF | Multidrug efflux RND transporter permease subunit | Antibiotic efflux |
| acrR | Multidrug efflux transporter transcriptional repressor AcrR | Antibiotic efflux |
| ampH | D-alanyl-D-alanine-carboxypeptidase/ endopeptidase AmpH | Antibiotic inactivation |
| baeR | Two-component system response regulator BaeR | Antibiotic efflux |
| baeS | Two-component system sensor histidine kinase BaeS | Antibiotic efflux |
| cpxA | Envelope stress sensor histidine kinase CpxA | Antibiotic efflux |
| crp | cAMP-activated global transcriptional regulator CRP | Antibiotic efflux |
| cyaA | Adenylate cyclase | Antibiotic target alteration |
| emrA | Multidrug efflux MFS transporter periplasmic adaptor subunit EmrA | Antibiotic efflux |
| emrB | Inner membrane component of tripartite multidrug resistance system | Antibiotic efflux |
| emrR | Transcriptional repressor MprA | Antibiotic efflux |
| fabI | Enoyl-ACP reductase FabI | Antibiotic target alteration |
| folP | Dihydropteroate synthase | Antibiotic target alteration |
| glpT | Glycerol-3-phosphate transporter | Antibiotic target alteration |
| golS | MerR family transcriptional regulator | Antibiotic efflux |
| gyrA | DNA gyrase subunit A | Antibiotic target alteration |
| gyrB | DNA gyrase subunit B | Antibiotic target alteration |
| hns | DNA-binding transcriptional regulator H-NS | Antibiotic efflux |
| kdpE | Two-component system response regulator KdpE | Antibiotic efflux |
| marA | MDR efflux pump AcrAB transcriptional activator MarA | Antibiotic efflux |
| marR | Multiple antibiotic resistance protein MarR | Antibiotic efflux |
| mdtA | Multidrug resistance protein MdtA | Antibiotic efflux |
| mdtB | Multidrug resistance protein MdtB | Antibiotic efflux |
| mdtC | Multidrug efflux RND transporter permease subunit MdtC | Antibiotic efflux |
| mdtG | Multidrug efflux MFS transporter MdtG | Antibiotic efflux |
| mdtH | Multidrug efflux MFS transporter MdtH | Antibiotic efflux |
| mdtK | Multidrug efflux MATE transporter MdtK | Antibiotic efflux |
| mdtM | Multidrug resistance protein MdtM | Antibiotic efflux |
| mdsA | Multidrug efflux RND transporter periplasmic adaptor subunit MdsA | Antibiotic efflux |
| mdsB | Multidrug efflux RND transporter permease subunit MdsB | Antibiotic efflux |
| mdsC | Efflux pump outer membrane protein | Antibiotic efflux |
| msbA | Lipid A ABC transporter ATP-binding protein/permease MsbA | Antibiotic efflux |
| murA | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | Antibiotic target alteration |
| nfsA | Nitroreductase A | Antibiotic target alteration |
| parC | DNA topoisomerase IV subunit A | Antibiotic target alteration |
| parE | DNA topoisomerase IV subunit B | Antibiotic target alteration |
| phoP | Two-component system response regulator PhoP | Antibiotic target alteration |
| ptsI | Phosphoenolpyruvate-protein phosphotransferase PtsI | Antibiotic target alteration |
| ramA | RamA family antibiotic efflux transcriptional regulator | Antibiotic efflux |
| sdiA | Transcriptional regulator SdiA | Antibiotic efflux |
| rpoB | DNA-directed RNA polymerase subunit beta | Antibiotic target alteration |
| soxS | Superoxide response transcriptional regulator SoxS | Antibiotic target alteration |
| soxR | Redox-sensitive transcriptional activator SoxR | Antibiotic target alteration |
| tetA | Tetracycline resistance protein | Antibiotic efflux |
| tolC | Outer membrane channel protein TolC | Antibiotic efflux |
| tuf | Elongation factor Tu | Antibiotic target alteration |
| uhpA | DNA-binding transcriptional activator UhpA | Antibiotic target alteration |
| uhpT | Hexose-6-phosphate:phosphate antiporter | Antibiotic target alteration |
| yojI | Multidrug ABC transporter permease/ATP-binding protein | Antibiotic efflux |
In this study, key genes encoding for chemotaxis, motility, surface attachment, quorum sensing (QS), matrix protein-encoding genes, biofilm transcriptional regulators, and matrix polysaccharide synthesis genes were also identified (Table 4). Some of the adhesion factors, such as flagella and pili, are vital for biofilm formation (Yuan et al., 2020). Genes related to flagellar (flgA, flgB, flgC, flgD, flgE, flgF, flgG, flgH, flgI, flgJ, flgK, flgL, flgM, flgN, flgO, flgP) identified in this study are essential for attachment but can serve different roles during Salmonella biofilm formation. Gene coding for -chemotaxis (cheV) in this study is involved in the transmission of sensory signals from the chemoreceptors to the flagellar motors. QS is the bacterial cell-to-cell communication signaling mechanism, which can regulate bacterial motility and biofilm formation (Yuan et al., 2018). The luxS gene detected in this study is responsible for the production of QS signaling molecule by converting S-ribosyl-L-homocysteine into homocysteine and DPD. EPS of S. Typhimurium biofilms is composed of cellulose, curli, and biofilm--associated protein, and can support the adherence and surface attachment of S. Typhimurium onto different surfaces (Maruzani et al., 2019). In this study, the genes for cellulose biosynthesis (bcsA, bcsB, bcsC, bcsE, bcsF, bcsG, bcsQ), curli production (csgA, csgB, csgE, csgF, csgG), fimbriae biogenesis (fimA, fimC, fimF, fimH, fimW, fimY, lpfB, yhcD, yhcA, mrkB) were identified. Biofilm formation by Salmonella has proved to be regulated by a complex genetic network involving interactions between regulators, and the major regulators including csgD, adrA, and flk were identified in this study. CsgD encoded by csgD is the biofilm control center to regulate expressions of major Salmonella biofilm constituents, control the transition between planktonic and biofilm cells, and even maintain the 3-D structure of biofilms (Chen et al., 2021). AdrA regulates genes encoding cellulose biosynthesis of Salmonella by changing cellular levels of c-di-GMP during biofilm formation (Chen et al., 2021). The large protein BapA (encoded by bapA) of S. Typhimurium was shown to be necessary for bacterial aggregation and pellicle formation by interconnecting individual cells.
Table 4. Genes essential for biofilm formation in the genome of Salmonella Typhimurium M3.
| Gene identified | General function | Biofilm regulation mechanism |
|---|---|---|
| cheV | Involved in the transmission of sensory signals from the chemo-receptors to the flagellar motors | Chemotaxis |
| luxS | S-ribosylhomocysteine lyase | Quorum sensing |
| flgA | Flagellar basal body P-ring formation protein FlgA | Flagellar |
| flgB | Flagellar basal body rod protein FlgB | Flagellar |
| flgC | Flagellar basal body rod protein FlgC | Flagellar |
| flgD | Flagellar hook assembly protein FlgD | Flagellar |
| flgE | Flagellar hook protein FlgE | Flagellar |
| flgF | Flagellar basal body rod protein FlgF | Flagellar |
| flgG | Flagellar basal body rod protein FlgG | Flagellar |
| flgH | Flagellar basal body L-ring protein FlgH | Flagellar |
| flgI | Flagellar basal body P-ring protein FlgI | Flagellar |
| flgJ | Flagellar assembly peptidoglycan hydrolase FlgJ | Flagellar |
| flgK | Flagellar hook-associated protein FlgK | Flagellar |
| flgL | Flagellar basal body–associated protein FliL | Flagellar |
| flgM | Flagellar motor switch protein FliM | Flagellar |
| flgN | Flagellar motor switch protein FliN | Flagellar |
| flgO | Flagellar type III secretion system protein FliO | Flagellar |
| flgP | Flagellar type III secretion system pore protein FliP | Flagellar |
| flgQ | Flagellar biosynthesis protein FliQ | Flagellar |
| bcsA | UDP-forming cellulose synthase catalytic subunit | Cellulose biosynthesis |
| bcsB | Cellulose biosynthesis cyclic di-GMP-binding regulatory protein BcsB | Cellulose biosynthesis |
| bcsC | Cellulose biosynthesis protein BcsC | Cellulose biosynthesis |
| bcsE | Cellulose biosynthesis protein BcsE | Cellulose biosynthesis |
| bcsF | Cellulose biosynthesis protein BcsF | Cellulose biosynthesis |
| bcsG | Cellulose biosynthesis protein BcsG | Cellulose biosynthesis |
| bcsQ | Cellulose biosynthesis protein BcsQ | Cellulose biosynthesis |
| csgA | Major curlin subunit | Curli production |
| csgB | Minor curlin subunit | Curli production |
| csgE | Curli production assembly/transport protein CsgE | Curli production |
| csgF | Curli production assembly/transport protein CsgF | Curli production |
| csgG | Curli production assembly/transport protein CsgG | Curli production |
| fimA | Type-1 fimbrial protein subunit | Biogenesis of fimbriae |
| fimC | Fimbrial chaperone protein FimC | Biogenesis of fimbriae |
| fimF | Fimbrial-like protein FimF | Biogenesis of fimbriae |
| fimH | Fimbrial protein FimH | Biogenesis of fimbriae |
| fimW | Fimbria biosynthesis transcriptional regulator FimW | Biogenesis of fimbriae |
| fimY | Fimbria biosynthesis regulator FimY | Biogenesis of fimbriae |
| lpfB | Fimbrial assembly chaperone | Biogenesis of fimbriae |
| yhcD | Fimbrial biogenesis outer membrane usher protein | Biogenesis of fimbriae |
| yhcA | Fimbrial chaperone protein | Biogenesis of fimbriae |
| mrkB | Fimbrial assembly chaperone | Biogenesis of fimbriae |
| csgD | Transcriptional regulator CsgD | Transcriptional regulator |
| adrA | Diguanylate cyclase AdrA | Transcriptional regulator |
| sdiA | transcriptional regulator SdiA | Transcriptional regulator |
| flk | Flagella biosynthesis regulator Flk | Flagellar regulator |
| bapA | Biofilm-associated protein BapA | Cell-surface protein |
| bssS | Biofilm formation regulatory protein BssS | Biofilm regulator |
| bsmA | Lipoprotein BsmA | Biofilm stress and motility protein |
| bdcA | SDR family oxidoreductase | Increases biofilm dispersal |
| yehA | Putative fimbrial-like adhesin protein | Contributes to adhesion to various surfaces in specific environmental niches |
Conclusion
In summary, the high biofilm-forming capacity and multi-drug resistance of S. Typhimurium from swine slaughterhouse evidenced by in vitro and genomic sequencing tests confirmed their persistence in the food industry. The results have significant implications for the food safety and public health globally, highlighting the importance of good hygienic practices and appropriate use of antibiotics in the industry. Furthermore, the development of more appropriate and effective approaches for control of Salmonella contamination control in the food industry are also critical.
Acknowledgments
This research was financially supported by the Natural Science Fund for Colleges and Universities in Jiangsu Province (21KJB550007), the Natural Science Foundation of Jiangsu Province (BK20210814), and China Postdoctoral Science Foundation (2021TQ0274, 2022M720120).
REFERENCES
Chen, S., Feng, Z., Sun, H., Zhang, R., Qin, T. and Peng, D., 2021. Biofilm-formation-related genes csgD and bcsA promote the vertical transmission of Salmonella Enteritidis in Chicken. Frontiers in Veterinary Science 7: 625049. 10.3389/fvets.2020.625049
CLSI (Clinical and Laboratory Standard Institute), 2016. Performance standards for antimicrobial susceptibility testing. Twenty-Second Informational Supplement.
Dawan, J. and Ahn, J., 2020. Assessment of cross-resistance potential to serial antibiotic treatments in antibiotic-resistant Salmonella Typhimurium. Microbial Pathogenesis 148: 104478. 10.1016/j.micpath.2020.104478
Diaz, M., Ladero, V., Rio, B.D., Redruello, B., Fernandez, M., Martin, M.C. and Alvarez, M.A., 2016. Biofilm-forming capacity in biogenic amine-producing bacteria isolated from dairy products. Frontiers in Microbiology 7: 591. 10.3389/fmicb.2016.00591
Enrique Castro-Vargas, R.E., Herrera-Sanchez, M.P., Rodriguez-Hernandez, R. and Rondon-Barragan, I.S., 2020. Antibiotic resistance in Salmonella spp. isolated from poultry: a global overview. Veterinary World 13: 2070–2084. 10.14202/vetworld.2020.2070-2084
Ge, H.W., Wang, Y.Z. and Zhao, X.H., 2022. Research on the drug resistance mechanism of foodborne pathogens. Microbial Pathogenesis 162: 105306. 10.1016/j.micpath.2021.105306
Harb, A., Habib, I., Mezal, E.H., Kareem, H.S., Laird, T., O’Dea, M. and Abraham, S., 2018. Occurrence, antimicrobial resistance and whole-genome sequencing analysis of Salmonella isolates from chicken carcasses imported into Iraq from four different countries. International Journal of Food Microbiology 284: 84–90. 10.1016/j.ijfoodmicro.2018.07.007
Ingle, D.J., Ambrose, R.L., Baines, S.L., Duchene, S., da Silva, A.G., Lee, D.Y.J., Jones, M., Valcanis, M., Taiaroa, G. and Ballard, S.A., 2021. Evolutionary dynamics of multidrug resistant Salmonella enterica serovar 4,[5],12:i:-in Australia. Nature Communications 12: 4786. 10.1038/s41467-021-25073-w
Lee, K.H., Lee, J.Y., Roy, P.K., Mizan, M.F.R., Hossain, M.I., Park, S.H. and Ha, S.D., 2020. Viability of Salmonella Typhimurium biofilms on major food-contact surfaces and eggshell treated during 35 days with and without water storage at room temperature. Poultry Science 99: 4558–4565. 10.1016/j.psj.2020.05.055
Maruzani, R., Sutton, G., Nocerino, P. and Marvasi, M., 2019. Exopolymeric substances (EPS) from Salmonella enterica: polymers, proteins and their interactions with plants and abiotic surfaces. Journal of Microbiology 57: 1–8. 10.1007/s12275-019-8353-y
Merino, L., Procura, F., Trejo, F.M., Bueno, D.J. and Golowczyc, M.A., 2019. Biofilm formation by Salmonella sp. in the poultry industry: detection, control and eradication strategies. Food Research International 119: 530–540. 10.1016/j.foodres.2017.11.024
Shen, W.W., Chen, H., Geng, J.W., Wu, R.A., Wang, X. and Ding, T., 2022. Prevalence, serovar distribution, and antibiotic resistance of Salmonella spp. isolated from pork in China: a systematic review and meta-analysis. International Journal of Food Microbiology 361: 109473. 10.1016/j.ijfoodmicro.2021.109473
Taylor, S.J. and Winter, S.E., 2020. Salmonella finds a way: metabolic versatility of Salmonella enterica serovar Typhimurium in diverse host environments. PLoS Pathogens 16: e1008540. 10.1371/journal.ppat.1008540
Tian, Y., Gu, D., Wang, F., Liu, B., Li, J., Kang, X., Meng, C., Jiao, X.N. and Pan, Z.M., 2021. Prevalence and characteristics of Salmonella spp. from a pig farm in Shanghai, China. Foodborne Pathogens and Disease 18: 477–488. 10.1089/fpd.2021.0018
Wang, H., Zhang, X., Zhang, Q., Ye, K., Xu, X. and Zhou, G., 2015. Comparison of microbial transfer rates from Salmonella spp. biofilm growth on stainless steel to selected processed and raw meat. Food Control 50: 574–580. 10.1016/j.foodcont.2014.09.049
Wang, Y.Z., Ge, H.W., Wei, W.Y. and Zhao, X.H., 2022. Research progress on antibiotic resistance of Salmonella. Food Quality and Safety 6: fyac035. 10.1093/fqsafe/fyac035
Yuan, L., Burmølle, M., Sadiq, F.A., Wang, N. and He, G., 2018a. Interspecies variation in biofilm-forming capacity of-psychrotrophic bacterial isolates from Chinese raw milk. Food Control 91: 47–57. 10.1016/j.foodcont.2018.03.026
Yuan, L., Hansen, M.F., Røder, H.L., Wang, N., Burmølle, M. and He, G., 2020. Mixed-species biofilms in the food industry: current knowledge and novel control strategies. Critical Reviews in Food Science and Nutrition 60: 2277–2293. 10.1080/10408398.2019.1632790
Yuan, L., Sadiq, F.A., Burmølle, M., Liu, T. and He, G., 2018b. Insights into bacterial milk spoilage with particular emphasis on the roles of heat-stable enzymes, biofilms, and quorum sensing. Journal of Food Protection 81: 1651–1660. 10.4315/0362-028X.JFP-18-094
Yuan, L., Sadiq, F.A., Wang, N., Yang, Z. and He, G., 2021. Recent advances in understanding the control of disinfectant--resistant biofilms by hurdle technology in the food industry. Critical Reviews in Food Science and Nutrition 61: 3876–3891. 10.1080/10408398.2020.1809345
Zhou, Z., Jin, X., Zheng, H., Li, J., Meng, C., Yin, K., Xie, X.L., Huang, C.Y., Lei, T.Y., Sun, X.Y., Xia, Z.M., Zeng, Y., Pan, Z.M. and Jiao, X.N., 2018. The prevalence and load of Salmonella, and key risk points of Salmonella contamination in a swine slaughterhouse in Jiangsu province, China. Food Control 87: 153–160. 10.1016/j.foodcont.2017.12.026
Zwama, M. and Nishino, K., 2021. Ever-adapting RND efflux pumps in Gram-negative multidrug-resistant pathogens: a race against time. Antibiotics 10: 774. 10.3390/antibiotics10070774