
Download






RESEARCH ARTICLE
Effects of wine processed Polygonatum polysaccharides on immunomodulatory effects and intestinal microecology in mice
Xiao-yan Xiao1#, Zhi-jun Guo2#, Xiaoman Li1, Peng Chen1, Yu Li1, Jiu-ba Zhang1, Chun-qin Mao1, De Ji1, Lian-lin Su1*, Bo Gao2*, Tu-lin Lu1*
1School of Pharmacy, Nanjing University of Chinese Medicine, Nanjing, Jiangsu, China;
2China Resources Sanjiu Pharmaceutical Co., Ltd, Shenzhen, China
#These authors have contributed equally to this work.
Abstract
Polygonatum sibiricum is a traditional Chinese medicinal and food homologous substance, usually used after processing with yellow wine. However, its main active ingredient, polysaccharide, has been less studied after wine processing. This study aimed to investigate the effect of Polygonatum polysaccharides after wine processing on the function of immunosuppressed mice and its related mechanisms. The yellow wine processed Polygonatum (YWPP) polysaccharides were extracted by the water extraction alcohol precipitation method. BALB/c mice were used to establish the immunosuppressive animal model with cyclophosphamide (CTX). The immunomodulatory effect of the YWPP polysaccharides Interleukin 2, interferon γ, immunoglobulin A, Immunoglobulin M, T lymphocyte subsets, and other indexes were detected by enzyme-linked immunosorbent assay, flow cytometry, and other technologies. To study the mechanism of immunomodulation of polysaccharides, 16s rDNA sequencing and Gas Chromatography/Mass Spectrometry (GC/MS) were used to analyze the changes of intestinal microbiota composition, diversity, and the content of short-chain fatty acids (SCFAs). The Spearman method was used to analyze the correlation between gut microbiota and various immune indexes and SCFAs. The results showed that the YWPP polysaccharides could significantly restore the body weight of immunosuppressed mice, improve the spleen and thymus, and regulate peripheral blood cell values. The YWPP polysaccharides can also effectively promote Interleukin 2, interferon γ, immunoglobulin A, and immunoglobulin M in immunosuppressed mice and regulate the expression of immune-related indexes. The YWPP polysaccharides promoted the production of SCFAs, significantly regulated the relative abundance of norank_f__norank_o__RF39, Rikenellaceae_RC9_gut_group, norank_f__UCG-010, and norank_f__norank_o__Clostridia_UCG-014; reversed CTX-induced metabolic abnormalities of mouse gut microbiota; and normalized them. Spearman correlation analysis showed that the relative abundance of gut microbiota was significantly correlated with various immune indices and SCFAs. In conclusion, the YWPP polysaccharides can improve the immune function of CTX-induced immunocompromised mice, promote cytokine upregulation and the relative abundance of immune-related beneficial bacteria in mice, and regulate the gut microbiota, thereby regulating host immunity. The results of this study will provide a theoretical basis for the practical application of the YWPP polysaccharides, health product development, and diversified product development.
Key words: gut microbiota, immunomodulatory, Polygonatum polysaccharides, polysaccharides, short-chain fatty acids, wine processed
*Corresponding Authors: Lian-lin Su, Nanjing University of Chinese Medicine, 138 Xianlin Avenue, Nanjing 210023, China. Email: [email protected]; Bo Gao, China Resources Sanjiu Pharmaceutical Co., Ltd, No. 1, Guanqing Road, Shenzhen 518000, China. Email: [email protected]; Tu-lin Lu, Nanjing University of Chinese Medicine, 138 Xianlin Avenue, Nanjing 210023, China. Email: [email protected]
Received: 24 October 2022; Accepted: 22 February 2023; Published: 10 April 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Polygonatum sibiricum is the dried rhizome of Polygonatum kingianum Coll. et Hemsl., Polygonatum sibiricum Red, or Polygonatum cyrtonema Hua (Chinese Pharmacopoeia, 2020). As Polygonatum irritates the throat, it must be processed before daily consumption and clinical use. At present, yellow wine processed Polygonatum (YWPP) sibiricum is commonly used in clinical practice and has been listed in the current edition of the “Chinese Pharmacopoeia” (Cheng et al., 2022). As the main active substances in Polygonatum sibiricum, Polygonatum polysaccharides are also quality evaluation indicators in the Chinese Pharmacopoeia (Wang et al., 2022). Polygonatum polysaccharides have proven to have functions of regulating immunity (Sun et al., 2020), anti-oxidation (Huang et al., 2021), anti-fatigue (Shen et al., 2021), and other functions. They can be extracted and added to drinks, wine, and food to prepare a series of health products.
In recent years, Polygonatum polysaccharides have made great progress in terms of structural characteristics and biological activity. The water extraction alcohol precipitation method is a mature method for extracting crude polysaccharides from Polygonatum (Liu et al., 2018). The separation and purification of polysaccharides were usually performed by gel column chromatography. The structure of polysaccharides is usually characterized by relative molecular mass determination, monosaccharide composition analysis, and sugar chain structure analysis. Previous studies have shown that compared with raw Polygonatum polysaccharides, the molecular weight of the wine processed polysaccharides decreased, the composition ratio of monosaccharides changed, and the proportion of galactose increased significantly (Chen et al., 2022). In FI-IR analysis, polysaccharides absorption peaks were similar, except that the wine processed polysaccharides produced absorption peaks of protein complexes. The wine processed Polygonatum polysaccharides can significantly increase immunomodulatory activity (Sun et al., 2020). However, the mechanism of action of the Y immunomodulation has not been studied in depth.
Recent studies have shown that Polygonatum polysaccharides can significantly stimulate serum immunoglobulins, improve the proliferation of peripheral blood T lymphocytes, and upregulate the expression of IL-2, IL-6, and γ-interferon genes, showing significant immunomodulatory activity (Shu et al., 2021). Further studies confirmed that Polygonatum polysaccharides could exert immunomodulatory effects by activating the toll-like receptor 4-live mitogen-activated protein kinase/nuclear factor signaling pathway (Long et al., 2019). On the other hand, the gut microbiota is also involved in the host’s immunity and metabolism. Under normal circumstances, the gut microbiota and immune system are in a state of dynamic balance, maintaining the stability of the intestinal microecology. When the number or species of bacteria changes significantly, the microecological balance is disrupted, the body’s metabolism and immune disorders are disturbed, and inflammation and related metabolic and immune diseases are promoted (Jordan et al., 2019). Based on recent studies, the immunomodulatory pharmacological effects of polysaccharides are closely related to the composition, metabolism, and metabolites of the gut microbiota (Xie et al., 2022). The gut microbiota has many metabolic pathways that are not possessed by the human body. The gut microbiota can ferment polysaccharides that cannot be absorbed and utilized by the human body to produce short-chain fatty acids (SCFAs), such as propionic acid and butyric acid, which have immunomodulatory effects on the human body; regulate the intestinal environment; improve the balance of flora; and play an immunomodulatory role (Liang et al., 2022; Ying et al., 2020). However, at present, few studies have used gut microbiota as the starting point to elucidate the immunomodulatory activity mechanism of polysaccharides.
Based on the research conclusions of predecessors on the structure of polysaccharides, the immunosuppressive animal model was established by cyclophosphamide (CTX), and the immunomodulatory effect of YWPP was evaluated by measuring the changes of mouse body weight, immune organ index, white blood cells (WBC), and red blood cells (RBC) in peripheral blood. The secretion levels of IL-2 and IFN-γ and the expression levels of immunoglobulins, IgM, and IgA in serum were detected by enzyme-linked immunosorbent assay (ELISA). The detection of T-lymphocyte subsets was performed using flow cytometry. The effect of YWPP on immune function and intestinal microbiota in immunosuppressed mice was explored through the Illumina MiSeq high-throughput sequencing platform and GC/MS analysis of SCFAs. The Spearman method was used to analyze the correlation between gut microbiota and various immune indexes and SCFAs. Polygonatum sibiricum has high nutritional value and broad application prospects. However, there is a lack of research on whether the immune effects of polysaccharide components commonly used in YWPP are related to their ability to regulate gut microbiota imbalance. Based on the findings of this study, the beneficial effects of YWPP on immune regulation and intestinal microecology were elucidated, and the development and application of YWPP series products were promoted.
Materials and Methods
Materials
Animals
BALB/c mice, male, SPF grade, body mass 18–20 g, purchased from the Laboratory Animal Management Department of Shanghai Institute of Family Planning Science, certificate number: 20180006030913. This study was carried out in accordance with the principles of the Basel Declaration and recommendations of all applicable international, national, and institutional guidelines for the care and use of animals. The protocol was approved by the Experimental animal center of Nanjing University of Chinese Medicine (license No. SYXK (Su) 2018-0049). Experimental environment: room temperature (22 ± 2)°C, humidity 50 ± 10%, 12 h light/12 h dark circadian rhythm-free diet.
Chemicals and solvents
Dried Rhizomes of Polygonatum sibiricum were purchased from China Resources Sanjiu Medical & Pharmaceutical Co. Ltd. (Anhui, China). The auxiliary yellow wine used for processing was purchased from Zhejiang Guyuelongshan Shaoxing Wine Co. Ltd. (Zhejiang, China); Lentinan (Hubei, China); Cyclophosphamide (CTX, DC Chemicals); IL-2, IFN-γ, IgM, IgA, and ELISA kits (Beijing, China); and antibodies against CD3e, CD4, and CD8a (eBioscience). Other chemicals used in this study were of analytical reagent grade.
Preparation of YWPP
According to the “Chinese Pharmacopoeia” 2020 edition, the wine processed Polygonatum pieces were prepared. The polysaccharides were extracted by water extraction and alcohol precipitation method. The yellow wine steamed Polygonatum was extracted twice with distilled water (1:10, w/v) at 100°C for 30 min. The combined filtrate was concentrated under reduced pressure. After cooling, absolute ethanol was added to the concentrate (1:4, v/v), which finally made the solution contain 80% alcohol. After standing for 12 h, the suspension was centrifuged at high speed. After discarding the supernatant, the crude polysaccharides were obtained and freeze-dried for preservation.
Immune regulation function of YWPP
Grouping and dosing
After adaptive feeding for 1 week, 40 BALB/c mice were randomly divided into normal control (NC), model (CTX), positive drug (LNT), and drug (YWPP) groups, with 10 mice per group. Except for the NC group, the immunosuppressive model was induced via the administration of intraperitoneal injection of 80 mg•kg−1 CT) to mice in the other groups for 3 days (Zhang et al., 2021). After modeling, mice in the treatment groups were intragastrically administered 0.6 g•kg−1 per day of their respective treatments (Dosage conversion according to the Chinese Pharmacopoeia), and mice in the positive group were intragastrically administered 60 mg•kg−1 of a lentinan suspension. The blank and model groups were administered the same volume of normal saline once per day for seven consecutive days.
Body mass and organ index measurement
The body weight of mice was measured daily to observe the changes in body weight after the establishment of the immunosuppressive model. After the experimental process, all groups of mice were anesthetized with pentobarbital sodium (1%, 50 mg/kg, I.P). Blood was collected from eye sockets of mice and finally they were killed by dislocation of the cervical vertebra. The thymus and spleen of mice were removed and weighed accurately after cleaning the blood and hair attached to the organs. The thymus and spleen indexes were calculated. The formula was as follows: organ index = organ weight (mg)/body weight (g).
Peripheral RBC and WBC count
Twenty microliters of orbital blood were mixed with a diluent. After that, WBC and RBC were counted within 12 h using a whole blood cell analyzer.
Measuring serum inflammatory factor levels by ELISA
Blood samples were collected from the medial canthus of mice and centrifuged at 3500 r•min−1 for 10 min to separate the upper serum. After that, 50 μL serum was retrieved, and serum IL-2 and IFN-γ levels were measured according to the ELISA kit instructions.
Immunoglobulin levels measuring by ELISA
The IgA and IgM expression levels were determined according to the same scheme under section “2.3.4” (Khan et al., 2022).
T lymphocyte testing
Blood samples were collected from the medial canthus of mice. After that, 0.8 mL of heparin sodium anticoagulant was added to RBC lysate precooled at 4°C, fully mixed, and centrifuged. Finally, the resulting supernatant was discarded. The above steps were repeated three times to obtain milky white lymphocytes, and 600 μL of PBS was added to prepare the cell suspension. Add mixed fluorescent antibodies CD3, CD4, and CD8 to the cell suspension. Incubate at 4°C for 30 min and rinse with PBS. Flow cytometry was used to detect and calculate the CD4+ and CD8+ content (Li et al., 2022).
Regulation function of YWPP in the gut microbiota of immunosuppressive mice
Fresh rat feces were collected in sterile tubes after the final treatment for 16s rDNA sequencing analysis. The Illumina HiSeq sequencing platform was used to construct small fragment libraries using the double-end sequencing method. Alpha, beta, and dominant species diversity analyses were carried out through read splicing filtering, Operational Taxonomic Unit (OTU) clustering, species annotation, and abundance analysis.
Effect of YWPP on SCFAs metabolism in immunosuppressed mice
The feces of mice in each group were collected in a grinding tube, and ultrapure water (Phosphoric acid (0.5%)) was added to prepare the solution. The supernatant was centrifuged for 10 min and extracted with 0.2 mL n-butanol solvent (2-ethylbutyric acid as an internal standard), mixed, sonicated at low temperature for 10 min, and centrifuged. The supernatant was then collected for GC-MS analysis. The chromatographic conditions were as follows: HP FFAP capillary column (30 m × 0.25 mm × 0.25 μm, Agilent J & W Scientific, Folsom, CA, USA); carrier gas, high-purity helium (purity not less than 99.999%); flow rate, 1.0 mL/min; injection port temperature, 180°C; injection volume, 1 μL; split injection, split ratio 10:1; and solvent delay, 2.5 min. The temperature program was as follows: initial temperature of the column oven, 80°C; increased to 120°C at 20°C/min and 160°C at 5°C/min, and maintained at 220°C for 3 min. The chromatographic conditions were as follows: electron bombardment ion source (EI); ion source temperature, 230°C; four-bar temperature, 150°C; transmission line temperature, 230°C; and electron energy, 70 eV. Ion scanning mode (SIM) was selected as the scanning mode.
Statistical analysis
GraphPad Prism 8 software was utilized for statistical analysis. Results were shown as means ± standard deviation. Each assay was conducted in three replicates. One-way ANOVA and Student’s t-test were used for statistical comparisons. P < 0.05 was considered to be statistically significant.
The Spearman method was used to analyze the correlation among gut microbiota, immune indexes, and SCFAs, using GraphPad Prism 8 software.
Results and Discussion
Effect of YWPP on immunomodulatory activity
Body mass and immune organ index
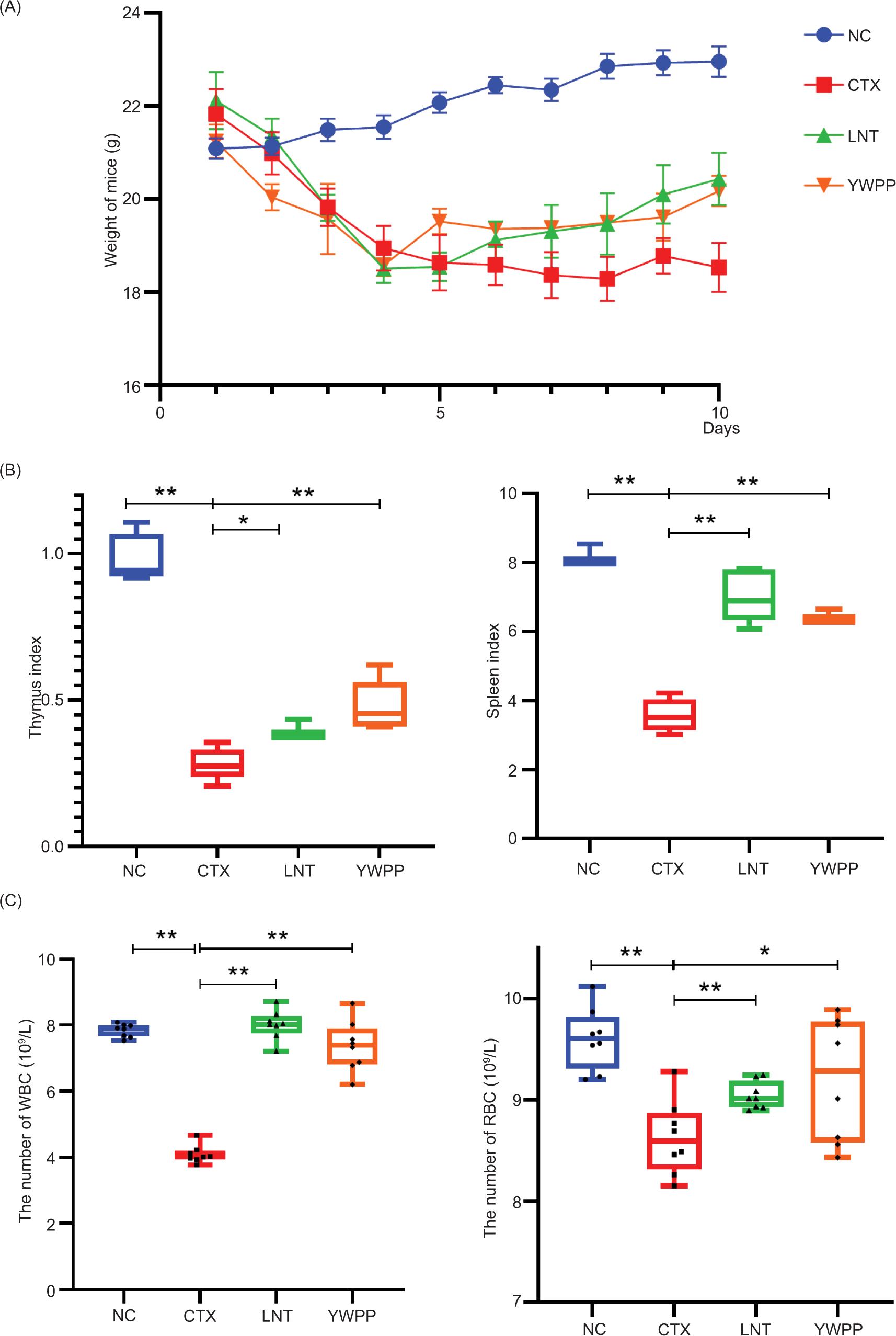
The average weight of mice in the CTX group was lower than that of mice in the NC group (P < 0.01). Compared to the CTX group, the average body weight of mice in the YWPP group increased significantly (P < 0.01) (Figure 1A).
Figure 1. Effect of yellow wine processed Polygonatum (YWPP) on the immunomodulatory activity. (A) & (B) Body mass and immune organ index; (C) white blood cells (WBC) and red blood cells (RBC) counts in peripheral blood of mice. Data are expressed as mean ± S.D. *0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01.
Compared with mice in the NC group, the spleen and thymus indexes of mice in the CTX group decreased significantly (P < 0.01), indicating that the model was successfully established. Compared to the CTX group, the thymus and spleen indexes in the YWPP group were significantly increased (P < 0.01) (Figure 1B).
WBC and RBC counts in peripheral blood of mice
The WBC and RBC counts of the CTX group decreased significantly (P < 0.01) compared to those of the NC group. The WBC counts of the YWPP groups increased significantly (P < 0.01). Compared with the CTX group, the RBC count of the YWPP group increased significantly (P < 0.05) (Figure 1C).
Expression level of IL-2 and IFN-γ
The expression levels of IL-2 and IFN-γ in the CTX group significantly decreased (P < 0.01, 0.05) compared to levels in the NC group. The expression levels of IL-2 and IFN-γ were significantly increased (P < 0.05) in the YWPP group (Figure 2A).
Figure 2. Effect of yellow wine processed Polygonatum (YWPP) on IL-2, IFN-γ, IgA, IgM expression, and T lymphocyte Subset activity. Data are expressed as mean ± S.D. *0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01.
Expression of IgA and IgM
Compared to the NC group, the expression levels of the IgA and IgM proteins in the CTX group decreased significantly (P < 0.01). Compared to the CTX group, the expression levels of IgA and IgM in the YWPP group increased significantly (P < 0.01) (Figure 2B).
Detection of T lymphocyte subset
Compared to the NC group, the CD4+/CD8+T lymphocytes in the peripheral blood of the CTX group decreased. Compared to the CTX group, CD4+/CD8+T lymphocytes were increased in the YWPP group (Figure 2C).
CTX as a chemotherapy drug with a broad spectrum of antitumors can significantly inhibit the function of immune organs and cause an imbalance of serum immunoglobulins and cytokines (Yoo et al., 2020). The selection of immune indicators was derived from cellular immunity and humoral immunity. They are two important immune mechanisms in the human body. Humoral immunity is an important body immune response, mainly through the release of antibodies by plasma cells to produce an immune response. Therefore, serum immunoglobulin level is an important indicator of humoral immune function detection. T cells play an important role in the immune system. The balance of CD4+ and CD8+ in the T lymphocyte subpopulation is essential for maintaining immune function (Jie et al., 2020). The results showed that YWPP had a significant immune enhancement effect. The body weight, thymus and spleen index, WBC and RBC number, and CD4+/CD8+T lymphocytes of immunosuppressed mice increased. The expression levels of IL-2, IFN-γ, IgA, and IgM increased.
Analysis of the composition and diversity of the gut microbiota
α and β diversity analysis of the gut microbiota
The intestinal microbial diversity of mice was analyzed using the Illumina MiSeq system. The rarefaction curves and estimators are shown in Figure 3A. The rarefaction curves indicated that the sequencing depth of the gut microbiota in each sample was fully captured and could be used for further analysis. The classification and community richness of the species were calculated using Simpson, Heip, and Shannon indexes (Figure 3B). The Simpson index in the YWPP group was significantly lower than that in the CTX group (P < 0.01); however, the Heip and Shannon richness indexes of the YWPP group were higher than those of the other groups. These results indicate that immunosuppressed mice’s microbial community richness and diversity were markedly affected, and YWPP had a good regulation and improvement effect on gut microbes.
Figure 3. α and β diversity analysis of the gut microbiota. (A) Rarefaction curves. (B) Simpson, Heip and Shannon’s index. (C) PLS-DA score plot. Values are expressed as mean ± S.D. Graph bars marked with different letters represent statistically significant results (P < 0.05) based on ANOVA with Duncan’s range tests, whereas bars marked with identical letters represent no statistically significant differences.
Partial least squares discriminant analysis (PLS-DA) was used to evaluate the clustering of the gut microbiota in each group (Figure 3C). A plot of the PLS-DA revealed that the CTX group had a significant shift along COMP1 (21.79%) at the phylum level compared with the NC and YWPP groups. The YWPP group had a significant shift along COMP1 (14.03%) at the genus level compared to the NC and CTX groups. Overall, each group showed good separation at the phylum and genus levels, indicating differences in the composition and diversity of the gut microbiota of mice in each group. PLS-DA indicated a significant difference in the bacterial community between the YWPP and CTX groups.
Composition analysis of the gut microbiota
The composition of the gut microbiota of the different groups was further investigated. The relative abundance of the gut microbiota classification units is presented as a stacking histogram (Figure 4A). All groups were mainly composed of Firmicutes, Bacteroidota, Desulfobacterota, Patescibacteria, and Actinobacteriota at the phylum level, and Lachnospiraceae_NK4A136_group, Lactobacillus, norank_f__Muribaculaceae, Desulfovibrio, and norank_f__norank_o__Clostridia_UCG-014 at the genus level. At the phylum and genus levels, we also counted the species abundance of each sample. We studied the community composition intuitively using the visualization method of the heatmap (Figure 4B). The YWPP and NC groups could be clustered well at the phylum and genus levels. They could be significantly separated from the CTX groups. Thus, the gut microbiota composition of mice in the YWPP group was similar to that of the NC group.
Figure 4. Composition of the gut microbiota. (A) Community bar-plot analysis at the phylum and genus levels. (B) Community heatmap analysis at the phylum and genus levels.
Changes in the relative abundance of each phylum and genus among the groups were compared (Figure 5). At the genus level, a decreased abundance of norank_f__norank_o__RF39, Colidextribacter, norank_f__norank_o__Clostridia_UCG-014, and Candidatus_Saccharimonas was found in the CTX group compared with the NC group (Figure 5A). At the phylum level, Bacteroides abundance was significantly lower in the CTX group. The relative abundance of Bacteroides in immunosuppressed mice increased significantly when YWPP was administered (Figure 5B). In addition, the Firmicutes/Bacteroidetes (F:B) ratio in the CTX group was significantly higher than that in the NC group, and significantly reduced in the YWPP group compared with the CTX group (Figure 5C). Notably, YWPP significantly increased the abundances of norank_f__norank_o__RF39, Rikenellaceae_RC9_gut_group, and norank_f__norank_o__Clostridia_UCG-014. Notably, YWPP can significantly decrease the relative abundance of Alistipes and Candidatus_Saccharimonas. Gut microbiota plays an important role in regulating the intestinal mucosal immune barrier, promoting the development and regulation of the immune system (Tatiana et al., 2017). Bacteroidetes are rich in carbohydrate-active enzymes (CAzyme), which can generate SCFAs and regulate the body’s energy balance (Cabral et al., 2022). Beneficial gut microbiota such as norank_f__norank_o__RF39 can activate the host’s immune system and improve the body’s immune defense function. Harmful gut microbiota such as Alistipes competes with the host for nutrients in the gut, secreting toxic compounds and inducing an inflammatory response. Therefore, YWPP modulated immunity by increasing the abundance of beneficial bacteria such as norank_f__norank_o__RF39 and decreasing the abundance of harmful bacteria such as Alistipes.
Figure 5. Comparative analysis of the differences between gut microbiota. (A) At the genus level, the comparative analysis of the relative abundance of seven intestinal microorganisms in the intestinal tract of mice in each group. (B) At the phylum level, the comparative analysis of the relative abundance of Bacteroidota in the intestinal tract of mice in each group. (C) The comparative analysis of the Firmicutes/Bacteroidetes (F:B) ratio in each group. *0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01, ***P ≤ 0.001.
Correlation analysis between gut microbiota and immune-related indicators
Probiotics play an immune adjuvant role in the collection of lymphoid tissues with antigen recognition sites in the intestine, activating the relevant lymphoid tissues in the intestinal mucosa, improving immune recognition, and inducing T and B lymphocytes and macrophages to produce cytokines. In this study, the relationship between immune-related indexes and gut microbiota was explored through Spearman analysis (Figure 6A). The significantly increased gut microbiota in the YWPP group compared with the CTX group was positively correlated with immune indexes (P < 0.01). For example, norank_f__norank_o__Clostridia_UCG-014 was significantly positively correlated with IgM and CD4+/CD8+ T lymphocytes (P < 0.01). Alistipes was a significantly reduced gut microbiota in the YWPP group, which was negatively correlated with thymus and spleen index, IgM, CD4+/CD8+ T lymphocytes, and RBC (P < 0.05). Bifidobacteria can activate the host intestinal mucosal immune system, and promote the secretion of IgA (Ivaylo and Honda, 2012; Rimoldi and Rescigno, 2005). Researchers (Cervantes-Barragan et al., 2010) found that Lactobacillus reuteri, together with a tryptophan-rich diet, could reprogram intraepithelial CD4+ T cells to immune regulatory T cells. Relevant studies have shown that the components in Polygonatum have the ability to restore the gut microflora interfered by CTX, which provided a reference for the conclusion of this study (He et al., 2022). In summary, YWPP may improve immunity by regulating the abundance of microflora.
Figure 6. The correlation among gut microbiota, immune indexes, and short-chain fatty acids (SCFAs) by Spearman’s correlation analysis. (A) Correlation analysis between gut microbiota and immune-related indicators. (B) Correlation analysis between gut microbiota and SCFAs. (C) Correlation analysis between SCFAs and immune-related indicators. *0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01, ***P ≤ 0.001.
Analysis of SCFAs metabolism
Effect of YWPP on the content of SCFAs
The contents of acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, isovaleric acid, hexanoic acid, and isohexanoic acid were investigated to determine the effects of YWPP on gut microbiota metabolites (Figure 7). The contents of all eight SCFAs significantly decreased in the colon of mice in the CTX group compared with those in the NC group. The SCFA contents of mice in the LNT and YWPP groups recovered to different levels compared with those in the CTX group. In conclusion, YWPP can improve metabolic disorders caused by immunosuppression by increasing SCFA content in feces.
Figure 7. Concentrations of short-chain fatty acids (SCFAs) in the colonic contents of mice. Data are expressed as mean ± S.D. *0.01 < P ≤ 0.05, **0.001 < P ≤ 0.01, ***P ≤ 0.001.
Correlation analysis between gut microbiota and SCFAs
The activity of CAZymes in gut microbiota increases SCFA production, reduces the expression of inflammatory factors, and increases the expression of tight junction proteins (El Kaoutari et al., 2013; Gao et al., 2018; Zhang et al., 2018). Therefore, correlation analysis was used to study the relationship between gut microbiota and SCFA content (Figure 6B). Bacteroidota was positively correlated with SCFAs (P < 0.01, 0.05). norank_f__norank_o__Clostridia_UCG-014, Alistipes and Rikenellaceae_RC9_gut_group were significantly positively correlated with Propanoic acid, Butanoic acid, Isovaleric acid, Valeric acid, Isohexanoic acid, Hexanoic acid, and Acetic acid (P < 0.01, 0.05). A recent study has shown that raw Polygonatum polysaccharides can increase beneficial bacteria in the gut microbiota, and the levels of acetic acid, propionic acid, and butyric acid in serum are significantly increased (Luo et al., 2022). In conclusion, YWPP may be zymolysis by Bacteroidota, norank_f__norank_o__Clostridia_UCG-014, Alistipes, and Rikenellaceae_RC9_gut_group to generate SCFAs.
Analyzation of the correlation between immune indexes and SCFAs
Based on the significant increase in SCFAs in the YWPP group, the association between SCFAs and immune indicators was analyzed (Figure 6C). IgM and CD4+/CD8+ T lymphocytes were significantly positively correlated with SCFAs (P < 0.01, 0.05). Acetic acid, Hexanoic acid, Butanoic acid, Isovaleric acid, and Isobutyric acid were significantly positively correlated with immunoassays other than RBC (P < 0.01, 0.05). Therefore, the increase in the content of SCFAs may be related to the recovery of metabolism by immune enhancement.
Conclusions
Polysaccharides in traditional Chinese medicine have various biological activities, such as anti-inflammatory, anti-oxidation, immune regulation, and antitumor activities. Furthermore, their mechanisms of action for the treatment or improvement of diseases are multifaceted. The gut microbiota and SCFAs play important roles in this process. The structure of Polygonatum polysaccharides changed after wine processing, but their immunomodulatory effect and mechanism of action lacked in-depth research. In this study, ELISA, flow cytometry, 16S rRNA sequencing, GC-MS, and other technologies were used to study the immunomodulatory effect of YWPP on immunosuppressed animals and the gut microbiota. The mechanism of YWPP enhancing immunity was discussed from the perspective of gut microbiota. The results of this study showed that YWPP can interact with the gut microbiota and co-metabolize to produce flora metabolites, thereby intervening in diseases. Gut microbiota is closely related to intestinal immunity, and polysaccharides can indirectly affect the immune regulatory function of the body through the gut microbiota, thereby treating diseases. It is hoped that the results of the study can provide a reference for the comprehensive development of YWPP as health adjuvants. Owing to the continuous in-depth study of polysaccharides in the regulation of gut microbiota, researchers have gradually realized the importance of polysaccharides in the treatment of diseases. Our research group has found that the structure of YWPP changes with the change of steaming time, and we will analyze the structure of polysaccharides at different steaming times and analyze their relationship with gut microbiota in the future.
Author Contribution
Xiaoyan Xiao and Zhijun Guo were involved in investigation, methodology, data curation, writing original draft, and review and editing. Xiaoman Li was concerned with investigation and formal analysis. Peng Chen was involved in investigation and methodology. Yu Li was responsible for methodology and sample collection. Jiuba Zhang took care of methodology and image processing. Chunqin Mao and De Ji did investigation and data curation. Tulin Lu, Lianlin Su, and Bo Gao were responsible for conceptualization, supervision, investigation, methodology, project administration, data curation, formal analysis, review and editing, and funding acquisition.
Acknowledgments
We are grateful for financial support from the National Key Research and Development Plan (No. 2018YFC1707000), Research of Quality Assurance System of Chinese Herbal Pieces (201507002-2), and the National Natural Science Foundation of China (No. 81373971). The gut microbiota and short-chain fatty acid metabolism analysis were performed using the online platform of Majorbio Cloud Platform (www.majorbio.com).
REFERENCES
Cabral, L., Persinoti, G.F., Paixão, D.A.A., Martins, M.P., Morais, M.A.B., Chinaglia, M., et al. 2022. Gut microbiome of the largest living rodent harbors unprecedented enzymatic systems to degrade plant polysaccharides. Nature Communications. 13(1): 629. 10.1038/s41467-022-28310-y
Cervantes-Barragan, L., Züst, R., Maier, R., Sierro, S., Janda, J., Levy, F., et al. 2010. Dendritic cell-specific antigen delivery by coronavirus vaccine vectors induces long-lasting protective antiviral and antitumor immunity. mBio. 1(4): e00171–10. 10.1128/mBio.00171-10
Chen, Z.R., Zhu, B.J., Chen, Z.X., Cao, W., Wang, J.Q., Li, S.P., et al. 2022. Effects of steam on polysaccharides from Polygonatum cyrtonema based on saccharide mapping analysis and pharmacological activity assays. Chinese Medicine. 17(1): 97. 10.1186/s13020-022-00650-3
Cheng, X., Ji, H., Cheng, C., Sun, Y., Cheng, H., Wang, D., et al. 2022. Comprehensive determination of the processing level of rhizome of Polygonatum sibiricum by macroscopic, micromorphological, and microscopic characterizations. Microscopy research and technique 85(7): 2669–2678. 10.1002/jemt.24121
Chinese Pharmacopoeia Commission, 2020. Chinese Pharmacopoeia (S). Part I. China medicine science and technology press 327–328.
El Kaoutari, A., Armougom, F., Gordon, J.I., Raoult, D. and Henrissat, B., 2013. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nature Reviews Microbiology 1(7): 497–504. 10.1038/nrmicro3050
Gao, H., Zhang, W.C., Wang, B.S., Hui, A.L., Du, B., Wang, T.T., et al. 2018. Purification, characterization and anti-fatigue activity of polysaccharide fractions from okra (Abelmoschus esculentus (L.) Moench). Food & function, 9(2), 1088–1101. 10.1039/c7fo01821e
He, Y.F., Huang, L., Jiang, P., Xu, G.P. and Sun, T.T., 2022. Immunological regulation of the active fraction from Polygonatum sibiricum F. Delaroche based on improvement of intestinal microflora and activation of RAW264.7 cells. Journal of Ethnopharmacology. 293: 115240. 10.1016/j.jep.2022.115240
Huang, S., Yuan, H.Y., Li, W.Q., Liu, X.Y., Zhang, X.J., Xiang, D.X., et al. 2021. Polygonatum sibiricum polysaccharides protect against MPP-induced neurotoxicity via the Akt/mTOR and Nrf2 pathways. Oxidative medicine and cellular longevity. 2021: 8843899. 10.1155/2021/8843899
Ivaylo, I.I. and Honda, K., 2012. Intestinal commensal microbes as immune modulators. Cell Host and Microbe 12(4): 496–508. 10.1016/j.chom.2012.09.009
Jie, D., Gao, T.T., Shan, Z.S., Song, J.Y., Zhang, M., Kurskaya, O., et al. 2020. Immunostimulating effect of polysaccharides isolated from Ma-Nuo-Xi decoction in cyclophosphamide-immunosuppressed mice. International journal of biological macromolecules. 146: 45–52. 10.1016/j.ijbiomac.2019.12.042
Jordan, S., Tung, N., Casanova-Acebes, M., Chang, C., Cantoni, C., Zhang, D., et al. 2019. Dietary intake regulates the circulating inflammatory monocyte pool. Cell 178(5): 1102–1114.e17. 10.1016/j.cell.2019.07.050
Khan, A.I., Rehman, A.U., Farooqui, N.A., Siddiqui, N.Z., Ayub, Q., Ramzan, M.N., et al. 2022. Effects of shrimp peptide hydrolysate on intestinal microbiota restoration and immune modulation in cyclophosphamide-treated mice. Molecules 27(5): 1720. 10.3390/molecules27051720
Li, Q.Y., Zhang, C.J., Xilin, T., Ji, M.Y., Meng, X.X., Zhao, Y.L., et al. 2022. Effects of koumiss on intestinal immune modulation in immunosuppressed rats. Frontiers in nutrition 9: 765499. 10.3389/fnut.2022.765499
Liang, Q.X., Zhao, Q.C., Hao, X.T., Wang, J.M., Ma, C.Y., Xi, X.F., et al. 2022. The effect of Flammulina velutipes polysaccharide on immunization analyzed by intestinal flora and proteomics. Frontiers in nutrition 9: 841230. 10.3389/fnut.2022.841230
Liu, N., Dong, Z.H., Zhu, X.S., Xu, H.Y. and Zhao, Z.X., 2018. Characterization and protective effect of Polygonatum sibiricum polysaccharide against cyclophosphamide-induced immunosuppression in Balb/c mice. International journal of biological macromolecules 107(Pt A): 796–802. 10.1016/j.ijbiomac.2017.09.051
Long, T.T., Liu, Z.J., Shang, J.C., Zhou, X., Yu, S., Tian, H., et al. 2019. Corrigendum to “Polygonatum sibiricum polysaccharides play anti-cancer effect through TLR4-MAPK/NF-κB signaling pathways” [Int. J. Biol. Macromol. 111 (2018): 813–821]. International journal of biological macromolecules 127: 703. 10.1016/j.ijbiomac.2018.08.087
Luo, Y., Fang, Q., Lai, Y., Lei, H., Zhang, D., Niu, H., et al. 2022. Polysaccharides from the leaves of Polygonatum sibiricum Red. regulate the gut microbiota and affect the production of short-chain fatty acids in mice. AMB Express. 12(1): 35. 10.1186/s13568-022-01376-z
Rimoldi, M. and Rescigno, M., 2005. Uptake and presentation of orally administered antigens. Vaccine 23(15): 1793–1796. 10.1016/j.vaccine.2004.11.007
Shen, W.D., Li, X.Y., Deng, Y.Y., Zha, X.Q., Pan, L.H., Li, Q.M., et al. 2021. Polygonatum cyrtonema Hua polysaccharide exhibits anti-fatigue activity via regulating osteocalcin signaling. International journal of biological macromolecules. 175: 235–241. 10.1016/j.ijbiomac.2021.01.200
Shu, G., Xu, D., Zhao, J., Yin, L.Z., Lin, J.C., Fu, H.L., et al. 2021. Protective effect of Polygonatum sibiricum polysaccharide on cyclophosphamide-induced immunosuppression in chickens. Research in veterinary science. 135: 96–105. 10.1016/j.rvsc.2020.12.025
Sun, T.T., Zhang, H., Li, Y., Liu, Y., Dai, W., Fang, J., et al. 2020. Physicochemical properties and immunological activities of polysaccharides from both crude and wine-processed Polygonatum sibiricum. International journal of biological macromolecules. 143: 255–264. 10.1016/j.ijbiomac.2019.11.166
Tatiana, T., Fenero, C.I.M. and Câmara, N.O.S., 2017. Intestinal barrier and gut microbiota: shaping our immune responses throughout life. Tissue Barriers 5(4): e1373208. 10.1080/21688370.2017.1373208
Wang, F.F., Jiang, Y.J., Jin, S.F., Wang, D.K., Wu, K.J., Yang, Q.W., et al. 2022. Structure characterization and bioactivity of neutral polysaccharides from different sources of Polygonatum Mill. Biopolymers 113(6): e23490. 10.1002/bip.23490
Xie, Z.Y., Bai, Y.X., Chen, G.J., Dong, W., Peng, Y.J., Xu, W.Q., et al. 2022. Immunomodulatory activity of polysaccharides from the mycelium of Aspergillus cristatus, isolated from Fuzhuan brick tea, associated with the regulation of intestinal barrier function and gut microbiota. Food research international 152: 110901. 10.1016/j.foodres.2021.110901
Ying, M.X., Yu, Q., Zheng, B., Wang, H., Wang, J.Q., Chen, S.P., et al. 2020. Cultured Cordyceps sinensis polysaccharides modulate intestinal mucosal immunity and gut microbiota in cyclophosphamide-treated mice. Carbohydrate polymers 235: 115957. 10.1016/j.carbpol.2020.115957
Yoo, J.H., Lee, Y.S., Ku, S. and Lee, H.J., 2020. Phellinus baumii enhances the immune response in cyclophosphamide-induced immunosuppressed mice. Nutrition research 75: 15–31. 10.1016/j.nutres.2019.12.005
Zhang, J., Zhou, H.C., He, S.B., Zhang, X.F., Ling, Y.H., Li, X.Y., et al. 2021. The immunoenhancement effects of sea buckthorn pulp oil in cyclophosphamide-induced immunosuppressed mice. Food & Function 2(17): 7954–7963. 10.1039/d1fo01257f
Zhang, T.H., Yang, Y., Liang, Y., Jiao, X. and Zhao, C.H., 2018. Beneficial effect of intestinal fermentation of natural polysaccharides. Nutrients 10(8): 1055. 10.3390/nu10081055