
Download

RESEARCH ARTICLE
Isolation and characterization of a broad-spectrum phage SapYZU11 and its potential application for biological control of Staphylococcus aureus
Hua Wen1, Lei Yuan1, Xuan Li1, Jun-hang Ye1, Ya-jie Li1, Zhen-quan Yang1, Wen-yuan Zhou1,2*
1College of Food Science and Engineering, Yangzhou University, Yangzhou, Jiangsu, China;
2College of Veterinary Medicine, Yangzhou University, Yangzhou, Jiangsu, China
Abstract
Staphylococcus aureus, a prominent pathogen, is frequently encountered in clinical and food-processing settings. Given its ability to develop antimicrobial resistance, effective control strategies are required to ensure microbial safety. In this study, four lytic S. aureus phages (SapYZU10, SapYZU11, SapYZU12, and SapYZU13) were isolated from sewage samples in Yangzhou, China. Their biological characteristics and bactericidal effect against S. aureus isolates in vitro and in milk and fresh pork were evaluated. Their activities remained relatively stable under stressful conditions (-80–70°C, pH 3.0–12.0). Notably, SapYZU11 (100%, 53/53) effectively lysed all 53 S. aureus strains, followed by SapYZU12 (90.57%, 48/53), SapYZU13 (79.25%, 42/53), and SapYZU10 (71.70%, 38/53). Among the phages with short latent periods (10–20 min), SapYZU11 had a larger burst size (152.00 plaque forming units [PFU]/mL) and no genes related to antibiotic resistance and virulence. Furthermore, SapYZU11 effectively eradicated S. aureus and its cocktail (YZUsa1, YZUsa4, YZUsa12, YZUsa14, and methicillin-resistance S. aureus (MRSA) JCSC 4744) in Luria–Bertani broth and both food items. Particularly in milk, SapYZU11 with a multiplicity of infection (MOI) of 100 inhibited MRSA JCSC 4744 strain and S. aureus cocktail with maximum reduction levels of 5.03 log (Lg) colony-forming unit (CFU)/mL and 2.80 Lg CFU/mL, respectively. Conversely, contaminated pork treated with three MOIs of SapYZU11 at 25°C and 4°C resulted in reductions of 0.29–1.29 Lg CFU/mL and 0.11–0.32 Lg CFU/mL, respectively. Therefore, SapYZU11 proved as a promising biocontrol agent against S. aureus in different food production settings.
Key words: bacteriophage, biocontrol, food security, milk, Staphylococcus aureus
*Corresponding Author: Dr. Wen-yuan Zhou, No. 196, West Huayang Road, Yangzhou, Jiangsu 225009, China. Email: [email protected]
Received: 16 November 2022; Accepted: 15 January 2023; Published: 1 April 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
In this fast-paced era, animal-derived food products are widely popular among consumers (Mahros et al., 2021). However, foods such as milk and meat, which are rich in proteins and nutrients, promote stubborn bacterial growth, potentially contaminating food products (Iyer et al., 2021). Such frequent and uncontrolled bacterial contamination causes a significant economic burden and poses a threat to public health (Zhou et al., 2018b). Staphylococcus aureus is a Gram-positive pathogen that causes a variety of diseases, including skin infections, respiratory infections, and food poisoning in animals and humans (Su et al., 2020). Moreover, because of antibiotic overuse, S. aureus has gradually evolved as a pathogen, such as methicillin-resistance S. aureus (MRSA), with the ability to cause life-threatening infections spreading rapidly worldwide (Samir et al., 2022). A recent conducted survey of food products, covering most of the provincial capitals of China, revealed that high prevalence of S. aureus was found in meat products as well as pasteurized milk (Zhang et al., 2022a). Furthermore, S. aureus accounts for approximately 11.26% of outbreaks caused by important foodborne pathogens in China (Wu et al., 2018). Considering that the use of antibiotics and chemical agents affects the flavor of food or even has adverse effects (Nikolic et al., 2020; Ravindran and Jaiswal, 2019) and that the development of new antibiotics is difficult, it is necessary to find alternative options to control S. aureus in food applications.
Bacteriophages (phages), which are ubiquitous viruses, can be isolated from river water, soil, farms, and domestic sewage and can effectively lyse bacterial cells (Bhetwal et al., 2017). In recent years, phages have gradually come to be known as natural and effective bactericidal substitutes because of the low efficiency of potent antibiotics in treating bacterial diseases, and hence play an important role in the prevention and control of foodborne pathogens (Pang et al., 2019; Santos and Azeredo, 2019). Some advantages of phages include target specificity protecting the microbiota of the host, capacity to multiply at the site of infection, and low production costs (Monteiro et al., 2019). The safety and efficacy characteristics of phages were proven in several human and animal models with bacterial infections (Miedzybrodzki et al., 2012; Takemura-Uchiyama et al., 2014). Nonetheless, the isolation and screening of phages having a broad-spectrum effect and high lysis ability remains one of the primary paths to control S. aureus in food products.
In this study, highly lytic S. aureus phages were isolated to analyze their biological characteristics. To confirm the potential of phages against S. aureus, their bactericidal effects were evaluated in vitro and food matrices (milk and fresh pork).
Materials and Methods
Strains and culture conditions
This study used 62 bacterial strains of different genera (Table 1). A total of 47 S. aureus strains (YZUsa 1–47) were isolated from 267 nonduplicate samples collected from three swine farms, two slaughter houses, and four markets (Yangzhou, China) from October 2019 to May 2020. The 267 samples included swine stool swabs (n = 30), swine nose swabs (n = 30), dust swabs (n = 30), swine carcass swabs (n = 40), and pork swabs (n = 137). All swab samples were collected using sterile dry swabs (Amies Agar Transport Swabs, Copan, Italy) as described previously and transported to laboratory within 2 h (Zhou et al., 2018a). All samples were screened for S. aureus isolates as described by Li et al. (2015) and confirmed by polymerase chain reaction (PCR) sequencing of products targeting the Staphylococcus-specific 16S ribosomal RNA (rRNA). Following other strains were used in this study: seven standard isolates, namely, S. aureus American Type Culture Collection (ATCC) 25923 and ATCC 29213, Staphylococcus epidermidis ATCC 12228, Escherichia coli China Center of Industrial Culture Collection (CICC) 10664, Salmonella E nteritidis CICC 21513, Listeria monocytogenes ATCC 1911, and Vibrio mimicus CICC 21613; four clinical MRSA isolates, namely, N315, MRSA85/2082, JCSC 4744, and WZ153; and three Staphylococcal strains, namely, Staphylococcus caprae SD2 and S. epidermidis YIC and SD1. All strains were obtained from the Key Laboratory of Prevention and Control of Biological Hazard Factors, Yangzhou University (Jiangsu, China).
Table 1. Host range of SapYZU10, SapYZU11, SapYZU12, and SapYZU13.
| No. | Strain | Species | Location | Source | Lysis performance | |||
|---|---|---|---|---|---|---|---|---|
| SapYZU10 | SapYZU11 | SapYZU12 | SapYZU13 | |||||
| 1. | YZUsa1 | S. aureus | Swine farm | Stool | +++ | +++ | +++ | +++ |
| 2. | YZUsa2 | S. aureus | Swine farm | Stool | – | +++ | + | – |
| 3. | YZUsa3 | S. aureus | Swine farm | Stool | +++ | +++ | +++ | +++ |
| 4. | YZUsa4 | S. aureus | Swine farm | Stool | +++ | +++ | +++ | +++ |
| 5. | YZUsa5 | S. aureus | Swine farm | Dust | ++ | +++ | ++ | + |
| 6. | YZUsa6 | S. aureus | Swine farm | Nose | +++ | +++ | +++ | +++ |
| 7. | YZUsa7 | S. aureus | Swine farm | Nose | – | + | +++ | + |
| 8. | YZUsa8 | S. aureus | Slaughter house | Nose | – | +++ | – | – |
| 9. | YZUsa9 | S. aureus | Slaughter house | Nose | – | + | + | – |
| 10. | YZUsa10 | S. aureus | Slaughter house | Nose | – | + | + | – |
| 11. | YZUsa11 | S. aureus | Slaughter house | Nose | – | +++ | ++ | – |
| 12. | YZUsa12 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 13. | YZUsa13 | S. aureus | Slaughter house | Nose | – | + | ++ | + |
| 14. | YZUsa14 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 15. | YZUsa15 | S. aureus | Slaughter house | Nose | – | +++ | + | – |
| 16. | YZUsa16 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 17. | YZUsa17 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 18. | YZUsa18 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 19. | YZUsa19 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 20. | YZUsa20 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 21. | YZUsa21 | S. aureus | Slaughter house | Nose | +++ | +++ | +++ | +++ |
| 22. | YZUsa22 | S. aureus | Slaughter house | Carcass | ++ | +++ | ++ | +++ |
| 23. | YZUsa23 | S. aureus | Slaughter house | Carcass | +++ | +++ | +++ | +++ |
| 24. | YZUsa24 | S. aureus | Market | Pork | +++ | +++ | +++ | +++ |
| 25. | YZUsa25 | S. aureus | Market | Pork | + | +++ | + | + |
| 26. | YZUsa26 | S. aureus | Market | Pork | + | +++ | + | + |
| 27. | YZUsa27 | S. aureus | Market | Pork | – | + | – | + |
| 28. | YZUsa28 | S. aureus | Market | Pork | – | ++ | +++ | – |
| 29. | YZUsa29 | S. aureus | Market | Pork | + | + | ++ | + |
| 30. | YZUsa30 | S. aureus | Market | Pork | – | + | – | – |
| 31. | YZUsa31 | S. aureus | Market | Pork | – | + | ++ | + |
| 32. | YZUsa32 | S. aureus | Market | Pork | +++ | + | +++ | + |
| 33. | YZUsa33 | S. aureus | Market | Pork | +++ | + | +++ | + |
| 34. | YZUsa34 | S. aureus | Market | Pork | ++ | + | +++ | + |
| 35. | YZUsa35 | S. aureus | Market | Pork | +++ | + | ++ | + |
| 36. | YZUsa36 | S. aureus | Market | Pork | +++ | + | ++ | + |
| 37. | YZUsa37 | S. aureus | Market | Pork | +++ | + | +++ | + |
| 38. | YZUsa38 | S. aureus | Market | Pork | +++ | +++ | +++ | +++ |
| 39. | YZUsa39 | S. aureus | Market | Pork | – | + | – | – |
| 40. | YZUsa40 | S. aureus | Market | Pork | +++ | +++ | +++ | +++ |
| 41. | YZUsa41 | S. aureus | Market | Pork | ++ | +++ | +++ | +++ |
| 42. | YZUsa42 | S. aureus | Market | Pork | +++ | + | +++ | + |
| 43. | YZUsa43 | S. aureus | Market | Pork | + | +++ | +++ | +++ |
| 44. | YZUsa44 | S. aureus | Market | Pork | + | + | +++ | + |
| 45. | YZUsa45 | S. aureus | Market | Pork | +++ | ++ | +++ | ++ |
| 46. | YZUsa46 | S. aureus | Market | Pork | ++ | +++ | + | + |
| 47. | YZUsa47 | S. aureus | Market | Pork | + | ++ | +++ | +++ |
| 48. | MRSA N315 | S. aureus | Hospital | Human | +++ | ++ | ++ | ++ |
| 49. | MRSA 85/2082 | S. aureus | Hospital | Human | – | ++ | ++ | – |
| 50. | MRSA JCSC 4744 | S. aureus | Hospital | Human | + | +++ | ++ | ++ |
| 51. | MRSA WZ153 | S. aureus | Hospital | Human | + | ++ | ++ | + |
| 52. | ATCC 25923 | S. aureus | - | - | – | + | – | – |
| 53. | ATCC 29213 | S. aureus | - | - | + | +++ | + | + |
| 54. | ATCC 12228 | S. epidermidis | - | - | – | + | – | – |
| 55. | YIC | S. epidermidis | - | Pickle | – | – | – | – |
| 56. | SD1 | S. epidermidis | - | Pickle | – | – | – | – |
| 57. | SD2 | Staphylococcus caprae | - | Pickle | – | – | – | – |
| 58. | CICC 10664 | Escherichia coli | - | CICC | – | – | – | – |
| 59. | Eh-YZU05 | Enterobacter hormaechei | - | Pork | – | – | – | – |
| 60. | CICC 21513 | Salmonella | - | CICC | – | – | – | – |
| 61. | ATCC 1911 | Listeriamonocytogenes | - | ATCC | – | – | – | – |
| 62. | CICC 21613 | Vibrio mimicus | - | CICC | – | – | – | – |
“+++” indicates clear and translucent plaque.
“++” indicates a slightly turbid plaque.
“+” indicates plaque turbidity.
“–” indicates no plaque.
ATCC: American Type Culture Collection; CICC: China Center of Industrial Culture Collection.
Isolation and propagation of phages
S. aureus phages were isolated from sewage samples collected between December 2019 and May 2020 from Yangzhou, China, using the previously described method (Zhou et al., 2021). In brief, 50 mL of the collected sewage was separated at 8,000 × g for 10 min, and the supernatant liquid was collected through 0.45- and 0.22-µm membranes (Sangon Biotech Co. Ltd., Shanghai, China). A 3-mL phage filtrate and a 100-µL host bacteria with stable growth were suspended in a double Luria–Bertani (LB) broth (2 ×LB; Hope Bio-Technology Co. Ltd., Qingdao, China), and incubated at 120 revolutions per minute (rpm) and 37°C for 24 h. The presence of phage was confirmed using the spot assay, and the high-titer phage suspensions of at least three enrichment processes were purified. Phages were diluted at different concentrations using saline magnesium (SM) dilution buffer. The host bacteria were mixed with phages and added to 5 mL of soft-top agar (LB containing 0.4% [w/v] agar), followed by pouring of the mixture into solid nutritional agar plates (LB containing 1.5% [w/v] agar) for overnight incubation. Large and round transparent patches were mixed in the buffer and a double layer was created; a pure phage solution was obtained after several cycles. The phage titers were determined in plaque forming units (PFU/mL) as described by Zhou et al. (2021).
Transmission electron microscopy analysis of phages
Transmission electron microscopy (TEM; Tecnai 12; Tecnai, Eindhoven, Netherlands) was used to examine the morphology of phages (Zhou et al., 2021). The phages were concentrated by isokinetic centrifugation using the cesium chloride density gradient method and incubated in a copper mesh. After 5 min, 2% phosphotungstic acid (Sigma-Aldrich Trading Co. Ltd., Shanghai, China) was added and allowed to dry naturally.
Multiplicity of infection of phages
As described by Luo et al. (2021), the phage solution diluted into different multiples was dispersed in host bacterial suspension (multiplicity of infection [MOI] = 100, 1, 0.1, 0.01, and 0.001), and the phage dilution ratio with a relatively high titer was used as the best MOI for the phage.
One-step growth curve
The experimental method for phage growth in host bacteria is based on previous studies with some adjustments (Luo et al., 2021). Phages stock dilutions (100 μL, 107 PFU/mL) were dispersed onto the pre-prepared S. aureus ATCC 29213 suspension (100 μL, 108 colony-forming unit [CFU]/mL) and were allowed to rest for 10 min. The adsorbent was centrifuged using the same procedure, and the precipitate was resuspended in 5-mL LB broth. The cultures were aspirated at 0 min, and then at 10, 20, 30, 40, 50, 60, 75, 90, 105, 120, 135, 150, 165, 180, 200, 220, and 240 min to determine their titers. The burst size was calculated by dividing the titer of the stably growing phage by the average of the original titer.
Thermal and pH stabilities
The phages were studied for their ability to survive at different temperatures and pH values. To measure thermal stability, 100 μL of phage suspension (109 PFU/mL) was transferred into 900-μL fresh LB broth, incubated at –80, –40, –20, 40, 50, 60, and 70°C and collected after 20 min, 40 min, and 1 h.
For estimating pH stability, the phage suspension was added to fresh LB broth with different pH values (pH was adjusted from 2.0 to 12.0 using HCl or NaOH) to a final concentration of 108 PFU/mL. Subsequently, the culture was incubated at 37°C for 1 h. After respective temperature and pH experiments, the phage titer was also determined using the double-layer agar method.
Lytic spectra of phages
Bacterial strains (n = 62) were dispersed in a double-layer dish, solidified to allow each phage solution (108 PFU/mL, 10 μL) to be dropped vertically onto the surface. Sodium chloride, Magnesium sulphate, and gelatine (SM) buffer was selected as a blank control and incubated at 37°C. The resulting plaque brightness was divided into four different grades (Li et al., 2021b) as shown in Table 1: clear and translucent (+++), slightly turbid (++), turbidity (+), and no plaque (–).
Bactericidal activity of phage against S. aureus in LB broth
Phage activity in the bacterial challenge test was determined by the 96-well plate culture method (Alves et al., 2014). The concentration of each of the S. aureus strain suspension (ATCC 29213, YZUsa1, YZUsa4, YZUsa12, YZUsa14, and MRSA JCSC 4744) was adjusted to 106 CFU/mL, and five S. aureus strains (YZUsa1, YZUsa4, YZUsa12, YZUsa14, and MRSA JCSC 4744) were selected to be mixed in equal volumes to prepare S. aureus cocktail. Thereafter, S. aureus or S. aureus cocktail was thoroughly mixed with phage (MOI = 0.01, 1, and 100) in equal proportions to make phages and bacterial mixtures. A 96-well plate containing 198-μL LB broth was supplemented with 2 μL of phages and bacterial mixtures, and statically cultured at 37°C for 24 h; the absorbance was measured at 600 nm every 3 h.
Genomic analysis of phage SapYZU11
The extracted phage genomic DNA was sequenced on an Illumina sequencer (Illumina, San Diego, CA, USA), and the sequencing results were processed according to the method described by (Zhou et al. (2021). Open reading frames (ORFs) were predicted using software packages (FGENESB, Glimmer, and GeneMarkS) and annotated based on the “NCBI Prokaryotic Genome Automatic Annotation Pipeline” and “eggNOG” functions. Transfer RNA (tRNA)-encoding genes were screened using tRNA Scan-SE. The completed phage genome sequence of SapYZU 11 was deposited in GenBank (accession number MW864250).
Bactericidal activity of phage against S. aureus in skimmed milk
The effect of purified phage on the growth of S. aureus in skimmed milk was measured according to previously described methods with certain modifications (Li et al., 2021a). Ultra-high temperature (UHT) skimmed milk was used in the current study to evaluate the effect of phage on S. aureus. Skimmed milk, 10 mL, was inoculated with 100-μL MRSA JCSC 4744 or S. aureus cocktail (106 CFU/mL), followed by treatment with 100 μL of phage solution (MOI = 0.01, 1, and 100) or SM buffer. This was then incubated with a static culture at 25°C or 4°C. Cultures (1 mL) were removed for bacterial counting at 0, 3, 6, 9, 12, 24, and 48 h (25°C) or on daily basis (7 days, 4°C).
Bactericidal activity of phage against S. aureus in fresh pork
The method as described by Li et al. (2021b) with minor modifications was used to measure the effect of phages on the growth of S. aureus in fresh pork. In order to prevent the interference of microbiota, fresh pork samples of 1 × 1 × 1 cm size were washed with 70% ethanol and treated with ultraviolet radiation (UV) for 1 h. The S. aureus cocktail (106 CFU/mL, 100 μL) was evenly distributed on the surface of each meat piece, followed by a stable attachment and addition of the same amount of phage solution (MOI = 0.01, 1 and, 100) in the same position. Finally, the samples were cultured in constant temperature incubators at 25°C and 4°C. A standard plate count was performed at 0, 3, 6, 9, 12, 24, and 48 h (25°C) or on daily basis (7 days, 4°C) to quantify the surviving cells.
Statistical analysis
All tests in this study were performed in triplicate. Results are presented as mean values, and variances were determined by standard deviation from mean values (Zhou et al., 2021). The SPSS software with two-way analysis of variance (IBM-Armonk, New York, USA) was used to calculate the significance of data, and performed to determine differences between control and treatment groups (thermal and pH stability, inhibition assays in vitro, and food matrix). P < 0.05 was considered statistically significant.
Results
Isolation and morphological analysis of phages
Four phages (SapYZU10, SapYZU11, SapYZU12, and SapYZU13) infecting the host S. aureus ATCC 29213 were obtained from the sewage samples of Yangzhou, China. TEM results (Figures 1A–1D) indicated that the head diameters of SapYZU10 and SapYZU11 phages measured 41.61 ± 3.39 nm and 26.75 ± 2.69 nm, respectively. SapYZU10 and SapYZU11 phages had similar short tails (13.56 ± 1.96 nm and 28.55 ± 2.03 nm, respectively), and belonged to the Podoviridae family. Phages SapYZU12 and SapYZU13 had an icosahedral head (75.47 ± 2.36 nm and 74.77 ± 5.32 nm, respectively) attached to a contracted tail (182.33 ± 11.26 nm and 104.05 ± 9.23 nm, respectively). These morphological characteristics indicated that phages SapYZU12 and SapYZU13 belonged to the Myoviridae family. Additionally, phage patches formed by the four mentioned phages were different, their diameters were more than 1 mm, and their sizes were uniform (Figures 1E–1H).
Figure 1. (A) Electron microscopy and (B) plaque morphology of S. aureus phages (SapYZU10, SapYZU11, SapYZU12, and SapYZU13). Images of phage plaques are magnified three times and shown with red squares.
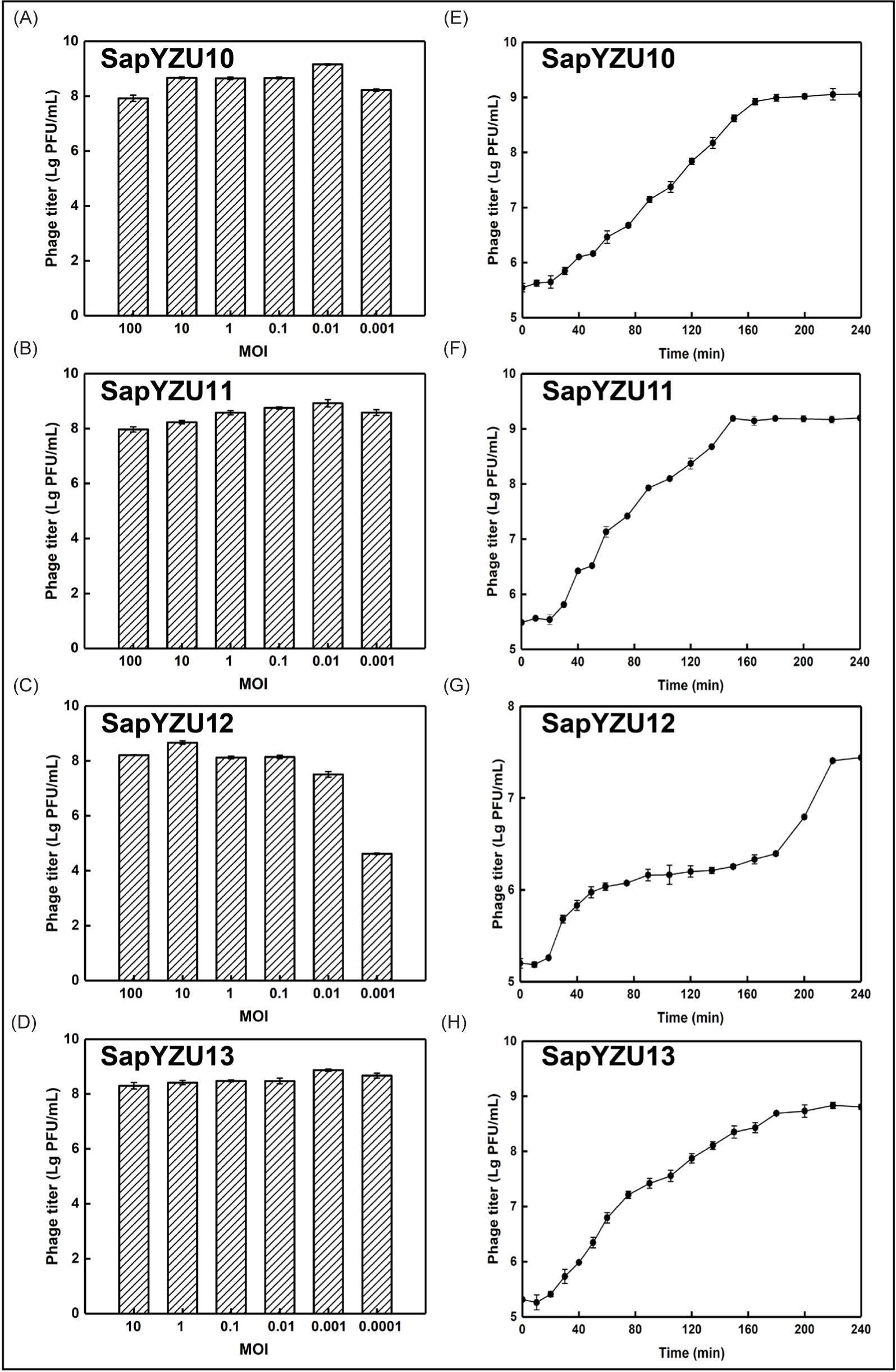
Optimal MOI and one-step growth of phages
When the four phages were in an optimal MOI, the titers produced were above 108 PFU/mL, especially for SapYZU11, in which titer was as high as 1.6 × 109 PFU/mL (Figures 2A–2D). As shown in Figures 2E–2H, the incubation periods of phages SapYZU10, SapYZU11, and SapYZU12 were 20 min, but their burst sizes were different. Compared to SapYZU10 (63.01 PFU/infected cell) and SapYZU12 (61.23 PFU/infected cell), SapYZU11 had the highest burst size of 152.00 PFU/infected cell. Although SapYZU13 had a short incubation period (10 min), its burst size (59.42 PFU/cell) was lowest of the four phages.
Figure 2. (A) Titer change of S. aureus phage under six multiplicity of infection (MOIs). (B) One-step growth curve of phages with an MOI of 0.1 with S. aureus ATCC 29213 as host bacterium.
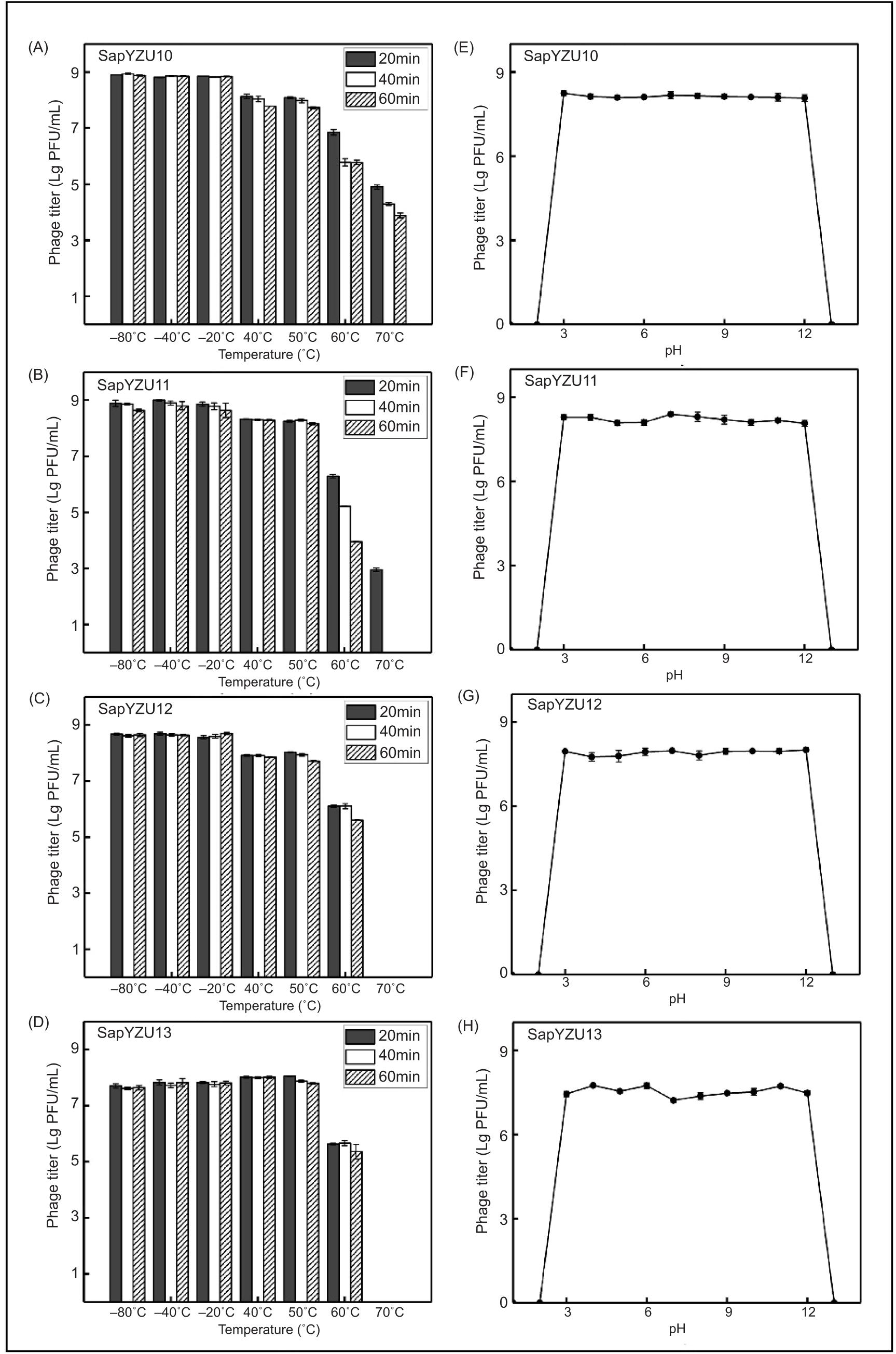
Thermal and pH stability of phages
The tolerance to distinct temperatures and pH conditions of phages are shown in Figure 3. Results showed no reduction in titers when the four phages were incubated between –80°C and –20°C, remaining at 8.85, 8.92, 8.64, and 7.79 log (Lg) PFU/mL. The titers of phages SapYZU10, SapYZU11, and SapYZU12 decreased by 0.74, 0.62, and 0.67 Lg PFU/mL, respectively, when incubated between 40°C and 50°C (P< 0.05) whereas the titers of SapYZU13 stabilized at 7.7 Lg PFU/mL. With the temperature rising to 60°C, activity of the four phages continually reduced by 1.26, 1.99, 1.86, and 2.40 Lg PFU/mL, respectively (P < 0.05). The thermal inactivation kinetics of SapYZU11 and SapYZU13 were similar when incubated at 70°C for 20 min and their titers decreased to 4.90 Lg PFU/mL and 2.95 Lg PFU/mL, respectively, whereas phages SapYZU12 and SapYZU13 were inactivated at 70°C. However, SapYZU11 was inactivated after incubation at 70°C for 40 min and longer, and only SapYZU10 retained a fraction of its activity after 1-h incubation, with a titer of 3.89 Lg PFU/mL (Figures 3A–3D). In addition, the results of acid–base solution on the growth of four phages showed that they retained a stable titer (8.13, 8.20, 7.91, and 7.52 Lg PFU/mL) from pH 3.0 to pH 12.0 for 1 h (Figures 3E–3H).
Figure 3. (A) Stability of S. aureus phages were maintained under different temperatures (–80°C, –40°C, –20°C, 40°C, 50°C, 60°C, and 70°C) for 2 h. (B) Stability of S. aureus phages was maintained under acid–base solution conditions (2.0, 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, 11.0, and 12.0) for 2 h.
Lytic spectrum of phages
Upon testing the phages against 62 common bacterial strains, including 53 S. aureus (Table 1), the results showed that SapYZU11 had the widest lytic spectrum, lysing 100% of the tested S. aureus strains and one S. epidermidis strain. Notably, 29 S. aureus (+++) strains were sensitive to SapYZU11, six S. aureus strains (++) showed slightly turbid plaques, and the remaining phage plaques were very cloudy (+). Compared to phage SapYZU12 with a lysis rate of 90.57% (48/53), SapYZU13 and SapYZU10 lysed only 79.25% (42/53) and 71.70% (38/53) of S. aureus strains, respectively. However, all four phages were unable to lyse bacteria other than S. aureus and S. epidermidis.
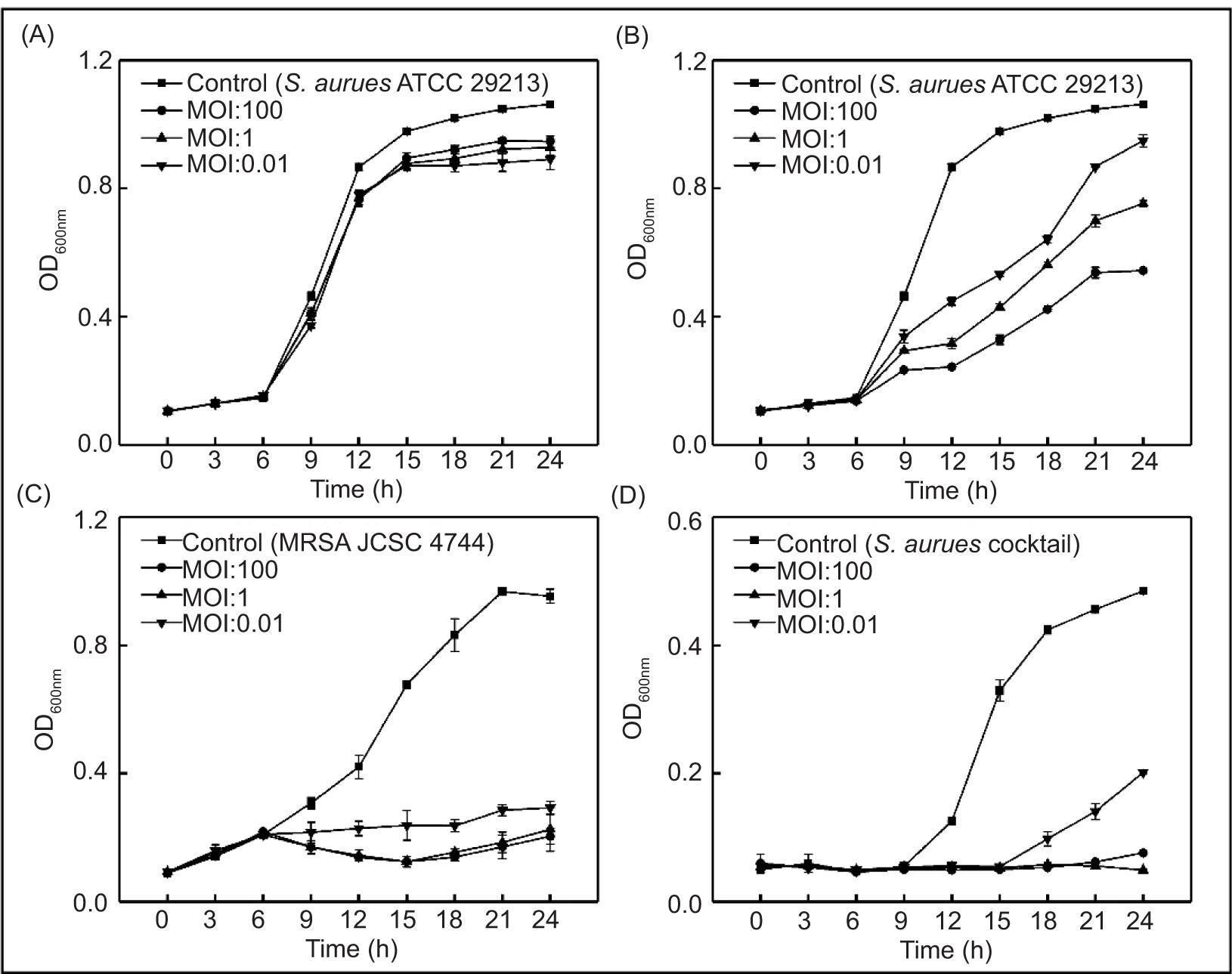
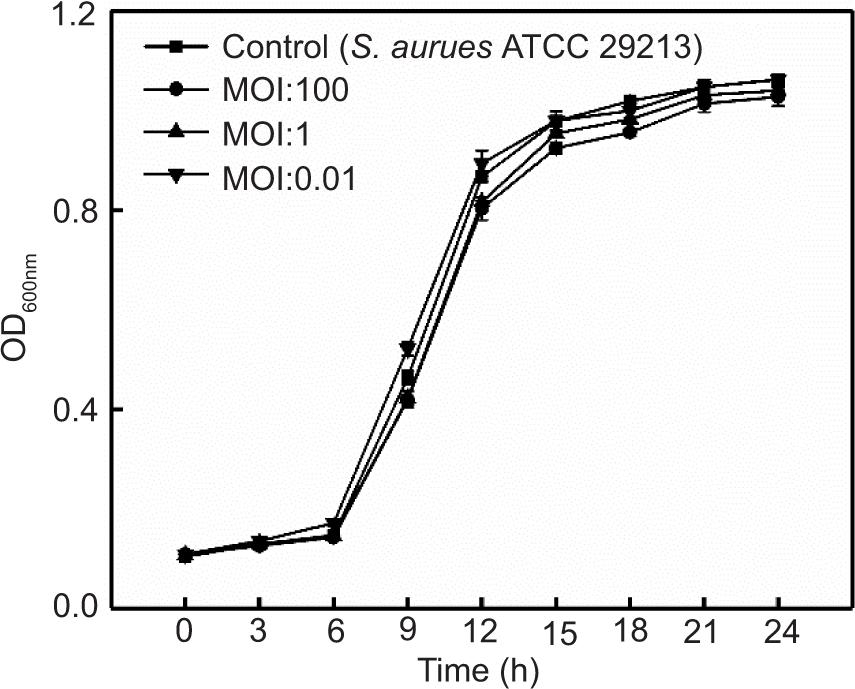
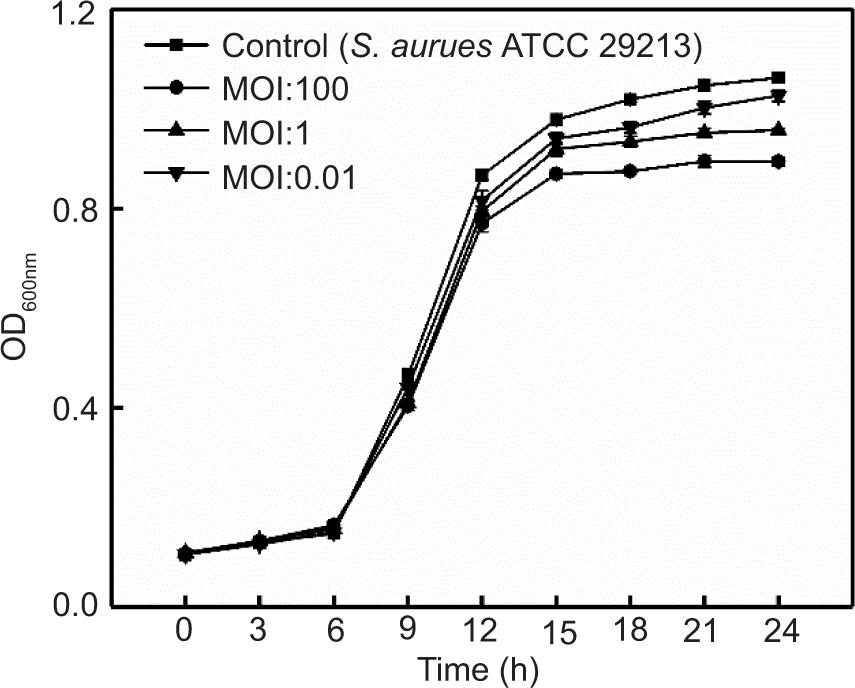
Phage SapYZU11 treatment in LB broth
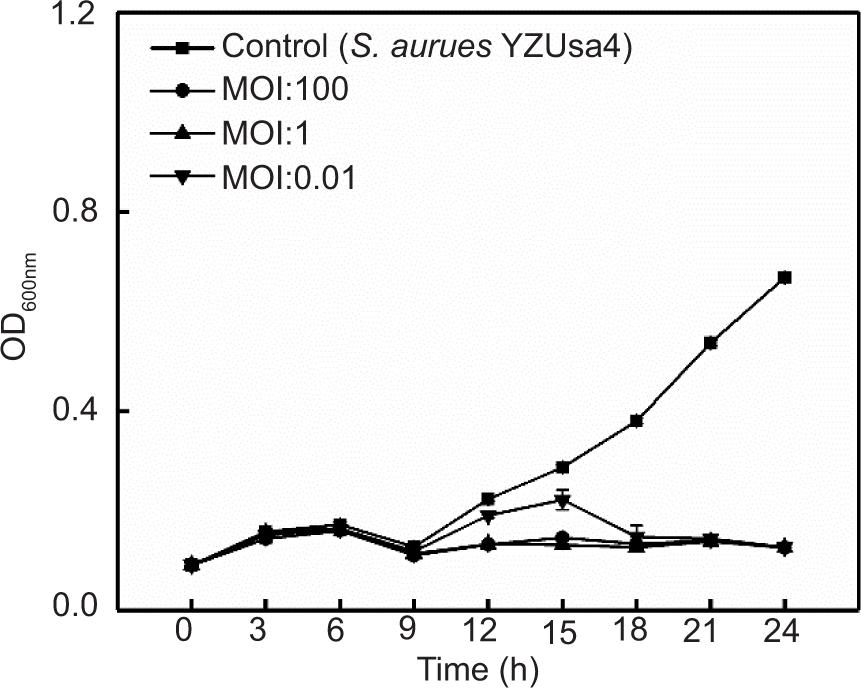
The effects of four phages on S. aureus ATCC 29213 in LB broth are shown in Figures 4A, 4B, and Supplementary Figures S1 and S2. The optical density (OD600nm) value of S. aureus ATCC 29213 without phage treatment was stable at 0.10 in the first 6 h, increased sharply to 0.91 at 6−15 h, and finally rose slowly at 15−24 h before stabilizing at 1.04. When the MOIs were 1 and 0.01, SapYZU11 reduced the OD600nm value of S. aureus to 0.45 (at 15 h, P < 0.05), which was significantly lower than that of the control group (0.98). However, SapYZU11 with an MOI of 100 showed better inhibitory effects, and the maximum reduction was 0.65 (at 15 h, P < 0.001). Compared to SapYZU11, phages SapYZU10, SapYZU12, and SapYZU13 showed maximum reduction in OD600nm values of S. aureus when the MOI was 100, which were 0.17 (at 24 h), 0.17 (at 24 h), and 0.06 (at 12 h), respectively (P < 0.05).
Figure 4. In vitro lytic activity of (A) phage SapYZU10, and (B) SapYZU11 against S. aureus ATCC 29213 in LB broth. In vitro lytic activity of phage SapYZU11 against (C) MRSA JCSC 4744 and (D) S. aureus cocktail (YZUsa1, YZUsa4, YZUsa12, YZUsa14, and MRSA JCSC 4744) in LB broth.
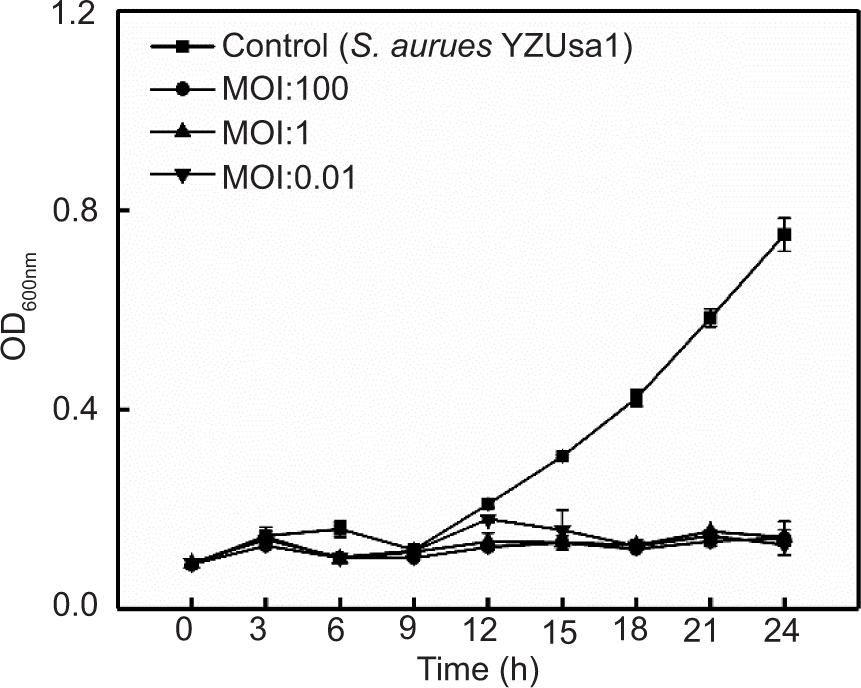
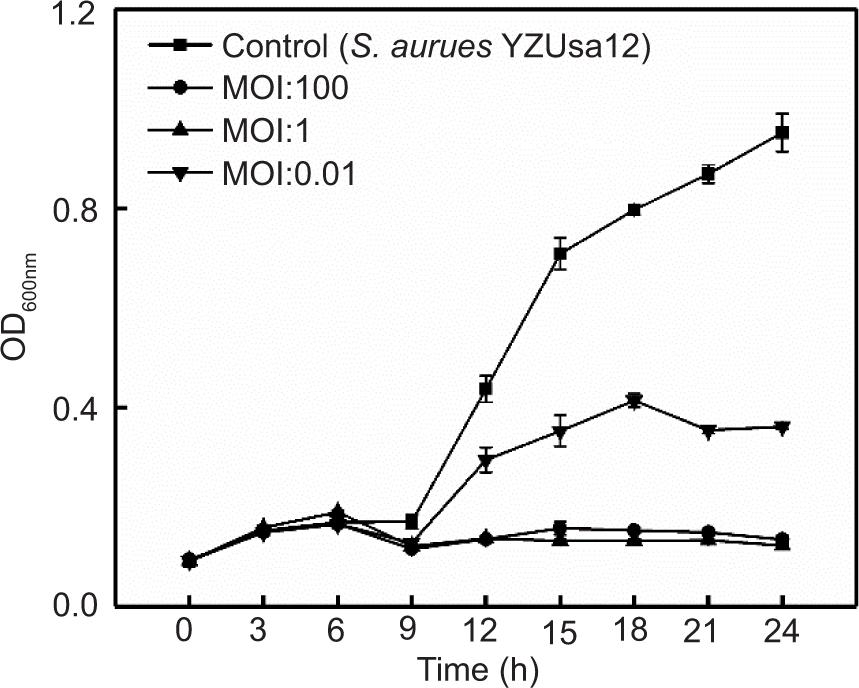
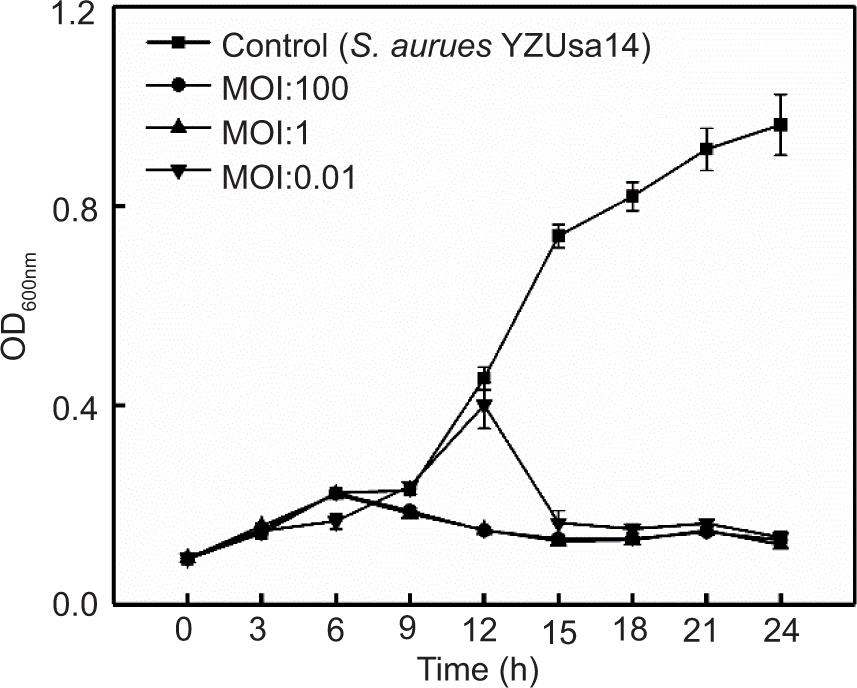
In addition, consistent with the expected results, following treatment with phage SapYZU11 at different MOI values (1 and 100) within 24 h, the OD600 nm values of five S. aureus strains, i.e., YZUsa1, YZUsa4, YZUsa12, YZUsa14, and MRSA JCSC 4744, were stabilized at 0.14, 0.12, 0.14, 0.15, and 0.16, respectively. By contrast, when MOI was 0.01, SapYZU11 only decreased the growth inhibition of S. aureus YZUsa12, and the OD600nm value was 0.36 at 24 h (P < 0.05; Figures 4C and S3–S6). Furthermore, when the MOI values were 1 and 100, SapYZU11 inhibited the growth of S. aureus cocktail for 24 h, and reduced their OD600nm values at 24 h by 0.41 and 0.43, respectively. However, the control effect of SapYZU11 with an MOI of 0.01 on S. aureus cocktail decreased slightly, and the best effect was achieved at 18 h, with the OD600nm value decreased by 0.33 (P < 0.05).
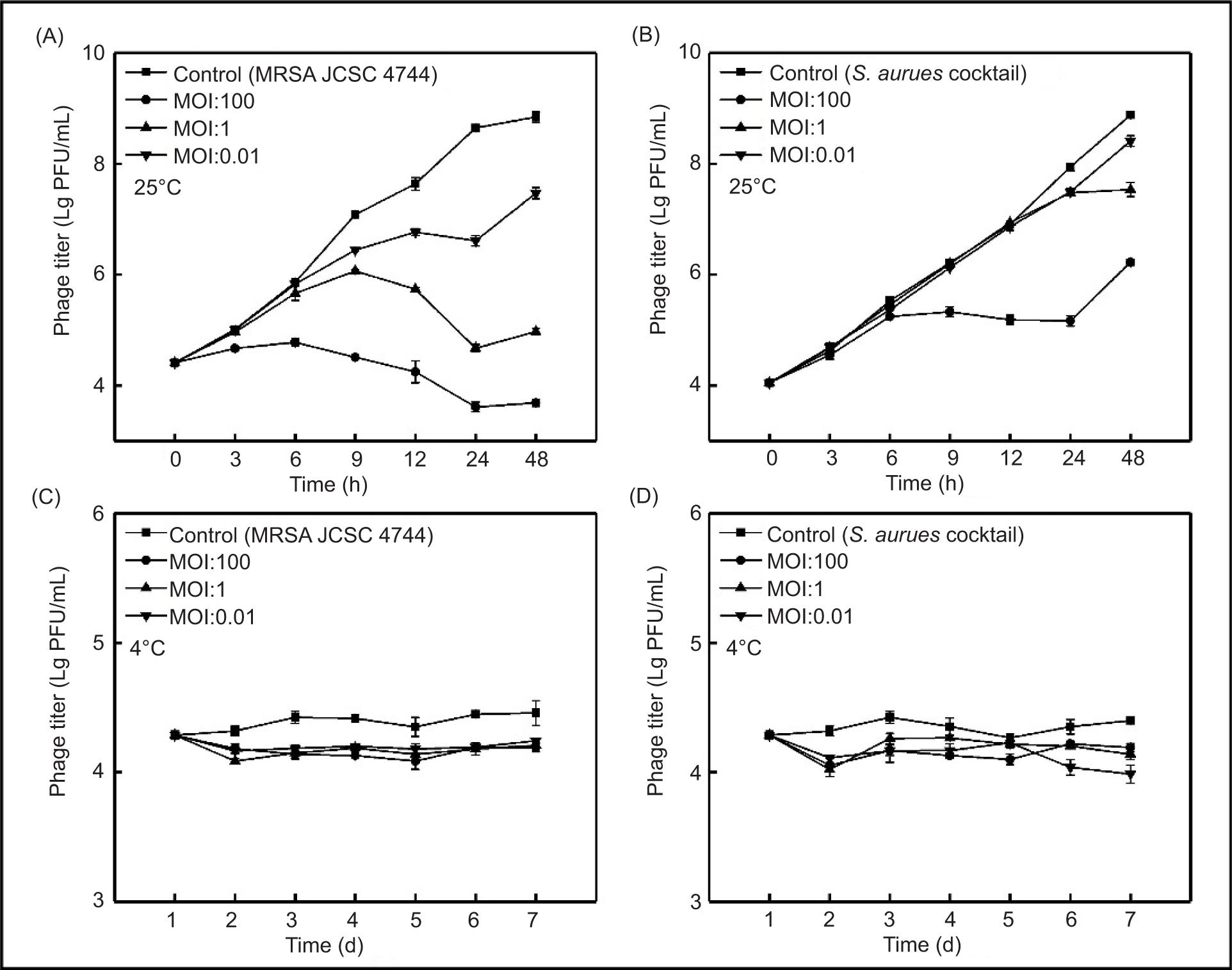
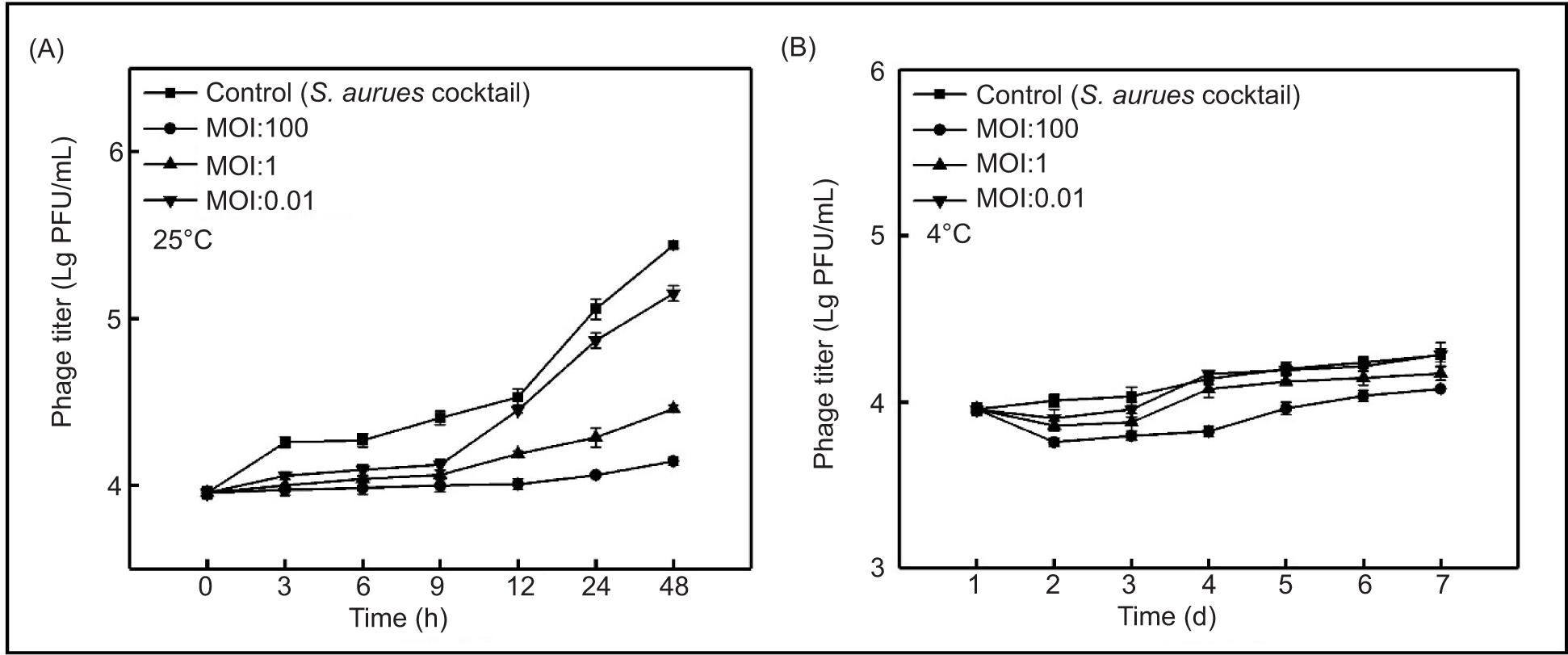
Phage SapYZU11 treatment in milk
The effect of SapYZU11 on the viability of S. aureus in milk was examined at 25°C and 4°C (Figures 5A and 5B). At 25°C, SapYZU11 with an MOI of 100 effectively inhibited the growth of MRSA JCSC 4744 and S. aureus cocktail for up to 24 h, thus reducing their counts by 5.03 Lg CFU/mL and 2.80 Lg CFU/mL (compared to the untreated milk sample), respectively (P < 0.001). Moreover, when the MOI values were 1 and 0.01, the growth of MRSA JCSC 4744 was significantly inhibited, and its count decreased by 3.97 Lg CFU/mL and 2.03 Lg CFU/mL, respectively (P < 0.001). At the same MOI values, the S. aureus cocktail counts were 7.50 Lg CFU/mL and 8.41 Lg CFU/mL at 48 h, respectively, whereas the count was 8.90 Lg CFU/mL for the untreated milk sample.
Figure 5. Effectiveness of phage SapYZU11 in reducing (A and C) MRSA JCSC 4744, and (B and D) S. aureus cocktail (106 CFU/mL) in milk at 4°C and 25°C. Phage SapYZU11 with three MOIs (0.01, 1, and 100) was mixed with S. aureus in milk, and the number of bacteria was determined at the indicated time points (4°C: 1, 2, 3, 4, 5, 6, and 7 day; 25°C: 0, 3, 6, 9, 12, 24, and 48 h).
The results of SapYZU11 on S. aureus in milk at 4°C are shown in Figures 5C and 5D. SapYZU11 at an MOI of 100 resulted in maximum reductions of MRSA JCSC 4744 and S. aureus cocktail at 4 days to 0.33 Lg CFU/mL and 0.29 Lg CFU/mL, respectively (P < 0.05). After phage treatment at MOIs of 0.01 and 1, the counts of MRSA JCSC 4744 decreased by 0.25 Lg CFU/mL (at 6 days) and 0.27 Lg CFU/mL (at 3 days), respectively, whereas the S. aureus cocktail counts at MOIs of 0.01 and 1 decreased by 0.11 Lg CFU/mL (at 2 days) and 0.16 Lg CFU/mL (at 3 d), respectively.
Phage SapYZU11 treatment in fresh pork
In order to test the bactericidal potential of SapYZU11 in meat products, fresh pork was artificially inoculated with S. aureus cocktail. When administered at MOIs of 1 and 100, SapYZU11 reduced the counts of viable S. aureus at 25°C by 0.98 Lg CFU/mL (at 48 h) and 1.29 Lg CFU/mL (at 48 h), respectively (P < 0.05; Figure 6A). On the other hand, at MOIs of 1 and 100, S. aureus decreased by 0.16 Lg CFU/mL (at 3 days) and 0.32 Lg CFU/mL (at 4 days) at 4°C, respectively (P < 0.05; Figure 6B). However, when using SapYZU11 at an MOI of 0.01 at 25°C, the maximum reduction of S. aureus was demonstrated to be 0.30 Lg CFU/mL at 48 h (P < 0.05). By contrast, the count of S. aureus was reduced by 0.11 Lg CFU/mL (at 2 days) following phage SapYZU11 treatment at 4°C.
Figure 6. Effectiveness of phage SapYZU11 in reducing S. aureus cocktail in fresh pork at (A) 4°C and (B) 25°C. S. aureus (106 CFU/mL) was divided into several parts and added to fresh pork and phage SapYZU11 at three MOIs (0.01, 1, and 100) at the same position after drying. The number of bacteria was determined at the indicated time points (4°C: 1, 2, 3, 4, 5, 6, and 7 day; 25°C: 0, 3, 6, 9, 12, 24, and 48 h).
Discussion
In the food industry, phages with specific characteristics are considered natural antimicrobials and potential tools against multi-drug-resistant bacteria (Lewis and Hill, 2020). In this study, four virulent S. aureus phages were isolated, and their inhibition against S. aureus in LB broth and food was analyzed. It is reported that latent period and burst size of phages are indicators of potential antibacterial capabilities (Yang et al., 2020). A phage with a larger burst size can have a practical advantage in therapy, because the phage population increases therapy’s initial dose by several hundred folds within a short time period (Amarillas et al., 2017). Previously, studies reported that the latent period of phages pSal-SNUABM-04 (Kwon et al., 2020) and SaGU1 (Shimamori et al., 2021) infecting S. aureus was 60 min and 40 min, respectively. In comparison, our data from the one-step growth curve showed that SapYZU11 and three other phages had shorter latent periods. Despite exhibiting a shorter latent period, SapYZU13 was not considered the best candidate to be a biocontrol agent because of its lower burst size. Another study reported that phages ME18 and ME126, which infect S. aureus, displayed a burst size of 114.00 PFU/cell and 140.00 PFU/cell, respectively (Gharieb et al., 2020). In contrast, SapYZU11 has a larger burst size, which is critical of efficacy as a biological antibacterial agent. These results indicated that SapYZU11 could be used as an effective biocontrol agent against S. aureus contamination in the food industry.
Temperature and pH values have significant effects on phage stability, and are considered important characteristics to be applied in the food industry (Cao et al., 2021). Isolated phages sustained for 1 h at temperatures below 50°C exhibited a stable titer. However, at 60°C, the activity of phages decreased, and after incubation at 70°C for 20 min, they gradually became inactive, similar to the S. aureus phage vB_SauS_JS02 (Zhang et al., 2022b). However, the temperature tolerance of SapYZU11 and other three phages was generally higher, compared to the four Staphylococcus phages Stab20–Stab23 isolated in Albania (Oduor et al., 2020). Moreover, SapYZU11 exhibited higher pH tolerance and stability than some previously reported phages, such as phage JD419 (Feng et al., 2021). Notably, SapYZU11 showed the highest activity at pH 12.0 for 2 h upon persistent incubation, allowing for its utilization in distinct food matrices and food production conditions with different pH values.
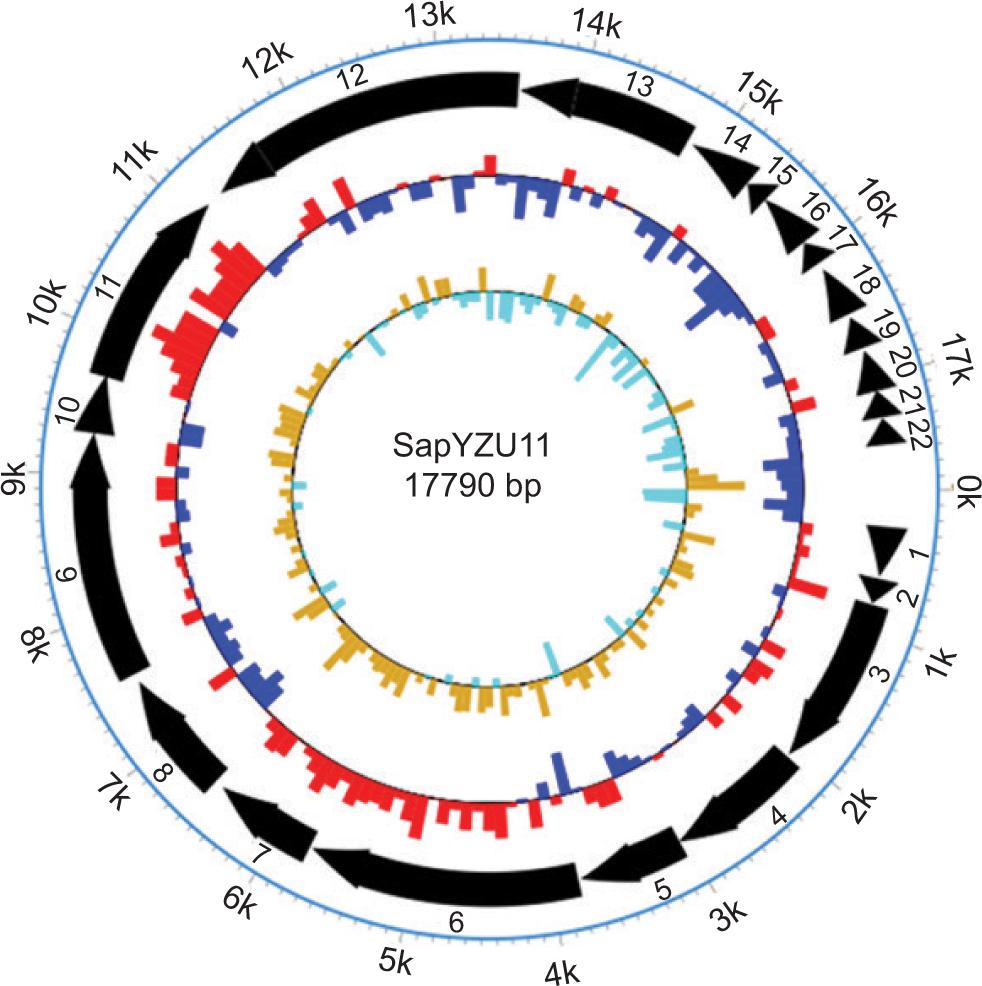
A phage with a wide range of hosts is a primary criterion of application in the food industry to ensure maximal bacterial infection, especially in fermented foods or farm animals (Meaden and Koskella, 2013). In a study involving 21 S. aureus strains as host spectra, phage vB_SauM_CP9 could lyse only 10 strains effectively (Kitamura et al., 2020). In another study involving four phages in the host spectrum, phage Trsa205 infected 37.50% (21/56) of S. aureus strains and lysed 66.07% (37/56) when combined with Trsa207, Trsa220, and Trsa222 phages (Abdurahman et al., 2021). In comparison, the isolated S. aureus phage SapYZU11 showed a broad range of hosts, with ability to infect a full range of tested S. aureus strains. Analysis of complete genome revealed that SapYZU11 harbored two genes encoding tail fiber protein (Figure S7 and Supplementary Table S1), which shared 83.60% and 97.60% amino acid sequence identity with phage CSA13 (Cha et al., 2019). Phage CSA13 had a broad host spectrum and lytic activity against strains, including methicillin-susceptible S. aureus (MSSA), MRSA, local S. aureus isolates, and non-aureus Staphylococci. Therefore, a broad host spectrum of SapYZU11 could be due to the existence of unique tail fiber proteins. Its genome did not encode any gene related to tRNA, drug resistance, lysogenicity, and virulence. These results identified that SapYZU11 had a potential application value as a biocontrol agent in food. However, the future research must elucidate the efficiency of plating (EoP), resistance, and safety of phage SapYZU11 for a more accurate and effective phage therapy in food applications.
In control of S. aureus in LB broth by phages, the ability of SapYZU11 to significantly inhibit the growth of all six S. aureus strains (ATCC 29213, YZUsa1, YZUsa4, YZUsa12, YZUsa14, and MRSA JCSC 4744) was reflected. This lytic activity was similar to the one reported by Ma et al. (2021), where the phage cocktail (LSA2366 and LSA2308) was found to decrease the content of S. aureus BM001 in LB broth within 24 h. Notably, in LB broth with S. aureus cocktail, SapYZU11 with MOIs of 1 and 100 displayed a high level of infective potential similar to phage LPSTLL (Guo et al., 2021). However, the optical density of S. aureus cocktail culture significantly decreased within 15 h of incubation with SapYZU11 at an MOI of 0.01, possibly because of the development of phage resistance or complex interactions between bacteria as reported by Zhou et al. (2021). Further research is required to understand the exact mechanism for developing bacterial resistance against phage SapYZU11.
Phage SapYZU11 had excellent characteristics and biological safety supporting its possible application in the food industry. In the present study, milk and fresh pork were used as representatives of food products frequently contaminated with S. aureus to evaluate the efficacy of SapYZU11 in controlling these pathogens in food matrices. The experimental results of artificial contamination of S. aureus in milk sample showed that when controlling the growth of MRSAJCSC 4744 at 25°C, the inhibitory rate of SapYZU11 at MOIs of 1 and 100 was more than 99.99%. This lytic activity was higher than that reported in a study conducted by Ma et al. (2021), showing an inhibitory rate of 99.97% against S. aureus in milk sample at 24 h following treatment with LSA2308 at an MOI of 10,000. Phage SapYZU11 exhibited greater efficiency than phage LSA2308, with the inhibition rate for S. aureus reaching 99.05% with SapYZU11 at an MOI of 0.01 in milk. In addition, SapYZU11 displayed a high level of infective potential against S. aureus cocktail at 25°C, and exhibited lytic activity in fresh pork samples. Their inhibitory effect was more apparent with growing numbers of MOIs at 25°C, as reported by Zhou et al. (2021). Similar results were also reported by Zhou et al. (2022), suggesting that addition of a high dosage of SapYZU11 could remove bacterial contamination from food products with greater efficiency.
At 4°C, concentration in S. aureus and S. aureus cocktail after 7-day SapYZU11 treatment was closely compared to the initial bacterial concentration (3.92–4.15 Lg CFU/mL) in milk and fresh pork samples whereas concentration in the control group was 4.38–4.51 Lg CFU/mL, which is consistent with the lytic activity of phages SPHG1 and SPHG3 (Esmael et al., 2021). In fact, phages require a stable host to reproduce and propagate; therefore, in low-temperature conditions both bacterial growth and phage activity are slow, consequently diminishing the outcome of phage treatment (Abhisingha et al., 2020). It was reported that some phages showed similar lytic activity at 4°C; however, the bactericidal mechanisms of phages at low temperature is still unclear (Witte et al., 2022). Accordingly, the temperature must be considered prior to the treatment with phage. Additionally, the data showed that inhibition of S. aureus by phage SapYZU11 in milk had a more significant effect than that in fresh pork, regardless of whether the phage was applied at 25°C or 4°C. These results revealed that aside from phage type and concentration, the food item could play a vital role in determining the efficacy of a bacteriophage intervention. This could have possibly resulted from the inability of phage particles to move and reach their host on the surface of a solid food or/and the emergence of phage resistance (Duc et al., 2020). Greater inhibition of S. aureus in milk could be attributed to the mobility of liquid milk, as the free movement offered in a liquid system would aid phage–bacteria engagement, increasing interactions between them.
Although SapYZU11 was proved to be effective in removing pathogen contamination from milk and fresh pork, one limitation of the current study was that the viable concentration of host bacteria could only be reduced to the initial level at the most but not below the detection limit (<1 Lg CFU/mL). This limitation also suggests how it was not be possible for one phage to control all pathogenic S. aureus. Therefore, further research is required wherein multiple combinations of phages, other food-grade antimicrobials, or higher concentration of phages could be used to improve their antibacterial effectiveness. In addition, in this study, the lysing activity of SapYZU11 was evaluated only in milk and pork. The future studies must include in vitro analysis in more food matrices and could focus on the stability, effectiveness, and safety of phage SapYZU11.
Conclusions
In conclusion, the four phages isolated in this study demonstrated high activity at a wide range of temperatures and pH values, and were able to lyse more than 60% of S. aureus. Notably, SapYZU11 lysed 100% of the tested S. aureus and revealed greater burst size (152.00 PFU/cell), compared to those of the other three phages. Moreover, SapYZU11 was more effective against S. aureus in LB broth, and significantly decreased the content of S. aureus cocktail in milk and fresh pork at 25°C and 4°C. These results suggested that phage SapYZU11 could be used as a novel natural biocontrol agent against S. aureus in food products.
Funding
This research was supported by the National Natural Science Foundation of China (grant numbers: 32102100 and 32001661) and China Postdoctoral Science Foundation (grant number: 2022M712695).
Conflicts of interest
The authors had no competing interests to declare.
Author contributions
Study conception and design: Wen-yuan Zhou; Acquisition of data: Hua Wen, Wen-yuan Zhou, Lei Yuan, Xuan Li, Jun-hang Ye, and Ya-jie Li; Analysis and interpretation of data: Hua Wen, Wen-yuan Zhou; Drafting of manuscript: Hua Wen, Wen-yuan Zhou and Lei Yuan; Critical revision: Hua Wen, Zhen-quan Yang, and Wen-yuan Zhou.
REFERENCES
Abdurahman, M.A., Tosun, I., Durukan, I., Khorshidtalab, M. and Kilic, A.O., 2021. Four temperate bacteriophages from methicillin-resistant Staphylococcus aureus show broad bactericidal and biofilm removal activities. Kafkas Universitesi Veteriner Fakultesi 27: 1. 10.9775/kvfd.2020.24680
Abhisingha, M., Dumnil, J. and Pitaksutheepong, C., 2020. Efficiency of phage cocktail to reduce Salmonella Typhimurium on chicken meat during low temperature storage. Food Science and Technology (LWT) 129: 109580. 10.1016/j.lwt.2020.109580
Alves, D.R., Gaudion, A., Bean, J.E., Perez, E.P., Arnot, T.C., Harper, D.R., et al. 2014. Combined use of bacteriophage K and a novel bacteriophage to reduce Staphylococcus aureus biofilm formation. Applied and Environmental Microbiology 80: 6694–6703. 10.1128/AEM.01789-14
Amarillas, L., Rubi-Rangel, L., Chaidez, C., Gonzalez-Robles, A., Lightbourn-Rojas, L. and Leon-Felix, J., 2017. Isolation and characterization of phiLLS, a novel phage with potential biocontrol agent against multidrug-resistant Escherichia coli. Frontiers in Microbiology 8: 1355. 10.3389/fmicb.2017.01355
Bhetwal, A., Maharjan, A., Shakya, S., Satyal, D., Ghimire, S., Khanal, P.R. et al. 2017. Isolation of potential phages against multidrug-resistant bacterial isolates: promising agents in the rivers of Kathmandu, Nepal. BioMed Research International 2017: 3723254. 10.1155/2017/3723254
Cao, Y., Zhang, Y., Lan, W. and Sun, X., 2021. Characterization of vB_VpaP_MGD2, a newly isolated bacteriophage with biocontrol potential against multidrug-resistant Vibrio parahaemolyticus. Archives of Virology 166: 413–426. 10.1007/s00705-020-04887-x
Cha, Y., Chun, J., Son, B. and Ryu, S., 2019. Characterization and genome analysis of Staphylococcus aureus Podovirus CSA13 and its anti-biofilm capacity. Viruses (Basel) 11(1): 54. 10.3390/v11010054
Duc, H.M., Son, H.M., Yi, H., Sato, J., Ngan, P.H., Masuda, Y., et al. 2020. Isolation, characterization and application of a polyvalent phage capable of controlling Salmonella and Escherichia coli O157:H7 in different food matrices. Food Research International 131: 108977. 10.1016/j.foodres.2020.108977
Esmael, A., Azab, E., Gobouri, A.A., Nasr-Eldin, M.A., Moustafa, M., Mohamed, S.A., e al. 2021. Isolation and characterization of two lytic bacteriophages infecting a multi-drug resistant Salmonella Typhimurium and their efficacy to combat salmonellosis in ready-to-use foods. Microorganisms 9(2): 423. 10.3390/microorganisms9020423
Feng, T., Leptihn, S., Dong, K., Loh, B., Zhang, Y., Stefan, M.I., et al. 2021. JD419, a Staphylococcus aureus phage with a unique morphology and broad host range. Frontiers in Microbiology 12: 602902. 10.3389/fmicb.2021.602902
Gharieb, R.M.A., Saad, M.F., Mohamed, A.S. and Tartor, Y.H., 2020. Characterization of two novel lytic bacteriophages for reducing biofilms of zoonotic multidrug-resistant Staphylococcus aureus and controlling their growth in milk. Food Science and Technology (LWT) 124: 109145. 10.1016/j.lwt.2020.109145
Guo, Y., Li, J., Islam, M.S., Yan, T., Zhou, Y., Liang, L., et al. 2021. Application of a novel phage vB_SalS-LPSTLL for the biological control of Salmonella in foods. Food Research International 147: 110492. 10.1016/j.foodres.2021.110492
Iyer, K.R., Revie, N.M., Fu, C., Robbins, N. and Cowen, L.E., 2021. Treatment strategies for cryptococcal infection: challenges, advances and future outlook. Nature Reviews Microbiology 19: 454–466. 10.1038/s41579-021-00511-0
Kitamura, N., Sasabe, E., Matsuzaki, S., Daibata, M. and Yamamoto, T., 2020. Characterization of two newly isolated Staphylococcus aureus bacteriophages from Japan belonging to the genus Silviavirus. Archives of Virology 165: 2355–2359. 10.1007/s00705-020-04749-6
Kwon, J., Kim, S.G., Kim, H.J., Giri, S.S., Kim, S.W., Lee, S.B., et al. 2020. Isolation and characterization of Salmonella jumbo-phage pSal-SNUABM-04. Viruses (Basel) 13(1): 27. 10.3390/v13010027
Lewis, R. and Hill, C., 2020. Overcoming barriers to phage application in food and feed. Current Opinion in Biotechnology 61: 38–44. 10.1016/j.copbio.2019.09.018
Li, Y., Chen, H., Shu, M., Zhong, C., Bi, Y., Yang, H., et al. 2021b. Isolation, characterization and application of an alkaline resistant virulent bacteriophage JN01 against Escherichia coli O157:H7 in milk and beef. Food Science and Technology (LWT) 144: 111266. 10.1016/j.lwt.2021.111266
Li, G., Wu, C., Wang, X. and Meng, J., 2015. Prevalence and characterization of methicillin susceptible Staphylococcus aureus ST398 isolates from retail foods. International Journal of Food Microbiology 196: 94–97. 10.1016/j.ijfoodmicro.2014.12.002
Li, H., Yang, X., Zhu, X., Gao, L., Rao, S., Yuan, L., et al. 2021a. Isolation and characterization of broad host-range of bacteriophages infecting Cronobacter sakazakii and its biocontrol potential in dairy products. Quality Assurance and Safety of Crops & Foods 13: 21–44. 10.15586/QAS.V13I3.890
Luo, D., Li, C., Wu, Q., Ding, Y., Yang, M., Hu, Y., et al. 2021. Isolation and characterization of new phage vB_CtuP_A24 and application to control Cronobacter spp. in infant milk formula and lettuce. Food Research International 141: 110109. 10.1016/j.foodres.2021.110109
Ma, F., Ning, Y., Wan, Q., Zou, L., Liu, Y., Chen, S., et al. 2021. Bacteriophages LSA2308 and LSA2366 infecting drug-resistant Staphylococcus aureus: isolation, characterization and potential application for milk safety. Food Science and Technology (LWT) 152: 112298. 10.1016/j.lwt.2021.112298
Mahros, M.A., Abd-Elghany, S.M. and Sallam, K.I., 2021. Multi-drug-, methicillin-, and vancomycin-resistant Staphylococcus aureus isolated from ready-to-eat meat sandwiches: an ongoing food and public health concern. International Journal of Food Microbiology 346: 109165. 10.1016/j.ijfoodmicro.2021.109165
Meaden, S. and Koskella, B., 2013. Exploring the risks of phage application in the environment. Frontiers in Microbiology 4: 358. 10.3389/fmicb.2013.00358
Miedzybrodzki, R., Borysowski, J., Weber-Dabrowska, B., Fortuna, W., Letkiewicz, S., Szufnarowski, K., et al. 2012. Clinical aspects of phage therapy. Advances in Virus Research 83: 73–121. 10.1016/B978-0-12-394438-2.00003-7
Monteiro, R., Pires, D.P., Costa, A.R. and Azeredo, J., 2019. Phage therapy: going temperate? Trends in Microbiology 27: 368–378. 10.1016/j.tim.2018.10.008
Nikolic, P., Mudgil, P. and Whitehall, J., 2020. The in vitro antibacterial effect of permethrin and formaldehyde on Staphylococcus aureus. MicrobiologyOpen 9: e1054. 10.1002/mbo3.1054
Oduor, J., Kadija, E., Nyachieo, A., Mureithi, M.W. and Skurnik, M., 2020. Bioprospecting Staphylococcus phages with therapeutic and bio-control potential. Viruses (Basel) 12(2): 133. 10.3390/v12020133
Pang, Z., Raudonis, R., Glick, B.R., Lin, T.J. and Cheng, Z., 2019. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnology Advances 37: 177–192. 10.1016/j.biotechadv.2018.11.013
Ravindran, R. and Jaiswal, A.K., 2019. Wholesomeness and safety aspects of irradiated foods. Food Chemistry 285: 363–368. 10.1016/j.foodchem.2019.02.002
Samir, S., El-Far, A., Okasha, H., Mahdy, R., Samir, F. and Nasr, S., 2022. Isolation and characterization of lytic bacteriophages from sewage at an egyptian tertiary care hospital against methicillin-resistant Staphylococcus aureus clinical isolates. Saudi Journal of Biological Science 29: 3097–3106. 10.1016/j.sjbs.2022.03.019
Santos, S.B. and Azeredo, J., 2019. Bacteriophage-based biotechnological applications. Viruses (Basel) 11(8): 737. 10.3390/v11080737
Shimamori, Y., Pramono, A.K., Kitao, T., Suzuki, T., Aizawa, S.I., Kubori, T., et al. 2021. Isolation and characterization of a novel phage SaGU1 that infects Staphylococcus aureus clinical isolates from patients with atopic dermatitis. Current Microbiology 78: 1267–1276. 10.1007/s00284-021-02395-y
Su, Y.C., Hung, W.W., Lin, J.M., Chang, C.C., Chen, Y.H., Lai, Y.L., et al. 2020. Tracking the evolution of the two successful CC59 methicillin-resistant Staphylococcus aureus clones in Taiwan: the divergence time of the two clades is estimated to be the 1980s. International Journal of Antimicrobial Agents 56: 106047. 10.1016/j.ijantimicag.2020.106047
Takemura-Uchiyama, I., Uchiyama, J., Osanai, M., Morimoto, N., Asagiri, T., Ujihara, T., et al. 2014. Experimental phage therapy against lethal lung-derived septicemia caused by Staphylococcus aureus in mice. Microbes & Infections 16: 512–517. 10.1016/j.micinf.2014.02.011
Witte, S., Huijboom, L., Klamert, S., van de Straat, L., Hagens, S., Fieseler, L., et al. 2022. Application of bacteriophages EP75 and EP335 efficiently reduces viable cell counts of Escherichia coli O157 on beef and vegetables. Food Microbiology 104: 103978. 10.1016/j.fm.2022.103978
Wu, Y.N., Liu, X.M., Chen, Q., Liu, H., Dai, Y., Zhou, Y.J., et al. 2018. Surveillance for foodborne disease outbreaks in China, 2003 to 2008. Food Control 84: 382–388. 10.1016/j.foodcont.2017.08.010
Yang, M., Liang, Y., Huang, S., Zhang, J., Wang, J., Chen, H., et al. 2020. Isolation and characterization of the novel phages vB_VpS_BA3 and vB_VpS_CA8 for lysing vibrio parahaemolyticus. Frontiers in Microbiology 11: 259. 10.3389/fmicb.2020.00259
Zhang, L., Shahin, K., Soleimani-Delfan, A., Ding, H., Wang, H., Sun, L., et al. 2022b. Phage JS02, a putative temperate phage, a novel biofilm-degrading agent for Staphylococcus aureus. Letters in Applied Microbiology 75: 643–654. 10.1111/lam.13663
Zhang, F., Wu, S., Lei, T., Wu, Q., Zhang, J., Huang, J., et al. 2022a. Presence and characterization of methicillin-resistant Staphylococcus aureus co-carrying the multidrug resistance genes cfr and lsa (E) in retail food in China. International Journal of Food Microbiology 363: 109512. 10.1016/j.ijfoodmicro.2021.109512
Zhou, Y., Li, L., Han, K., Wang, L., Cao, Y., Ma, D., et al. 2022. A polyvalent broad-spectrum Escherichia phage Tequatro virus EP01 capable of controlling Salmonella and Escherichia coli contamination in foods. Viruses (Basel) 14. 10.3390/v14020286
Zhou, W.Y., Li, X., Osmundson, T., Shi, L., Ren, J. and Yan, H., 2018a. WGS analysis of ST9-MRSA-XII isolates from live pigs in China provides insights into transmission among porcine, human and bovine hosts. Journal of Antimicrobial Chemotherapy 73: 2652–2661. 10.1093/jac/dky245
Zhou, W.Y., Li, X., Shi, L., Wang, H.H. and Yan, H., 2018b. Novel SCCmec type XII methicillin-resistant Staphylococcus aureus isolates identified from a swine production and processing chain. Veterinary Microbiology 225: 105–113. 10.1016/j.vetmic.2018.09.007
Zhou, W.Y., Sun, S.F., Zhang, Y.S., Hu, Q., Zheng, X.F., Yang, Z.Q., et al. 2021. Isolation and characterization of a virulent bacteriophage for controlling Salmonella Enteritidis growth in ready-to-eat nixed-ingredient salads. Journal of Food Protection 84: 1629–1639. 10.4315/JFP-20-460
Supplementary
Figure S1. In vitro lytic activity of phage SapYZU12 against S. aureus ATCC 29213 in LB broth.
Figure S2. In vitro lytic activity of phage SapYZU13 against S. aureus ATCC 29213 in LB broth.
Figure S3. In vitro lytic activity of phage SapYZU11 against S. aureus YZUsa1 in LB broth.
Figure S4. In vitro lytic activity of phage SapYZU11 against S. aureus YZUsa4 in LB broth.
Figure S5. In vitro lytic activity of phage SapYZU11 against S. aureus YZUsa12 in LB broth.
Figure S6. In vitro lytic activity of phage SapYZU11 against S. aureus YZUsa14 in LB broth.
Figure S7. Whole genome map of phage SapYZU11.
Table S1. Genomic function annotation of SapYZU11.
| ORFs | Start | End | Strand | Length (bp) | Protein (aa) | Putative function | Best phage homolog | Identity (%) | Accession No. |
|---|---|---|---|---|---|---|---|---|---|
| ORF1 | 266 | 625 | + | 360 | 120 | Hypothetical protein | Staphylococcus phage SA03-CTH2 | 77.87 | QDH84802.1 |
| ORF2 | 638 | 820 | + | 183 | 61 | Hypothetical protein | Staphylococcus phage SAP-2 | 100.00 | YP_001491544.1 |
| ORF3 | 827 | 2053 | + | 1,227 | 409 | Major head protein | Staphylococcus phage SAP-2 | 98.53 | YP_001491544.2 |
| ORF4 | 2068 | 3051 | + | 984 | 328 | Upper collar protein | Staphylococcus phage CSA13 | 98.17 | YP_009819982.1 |
| ORF5 | 3044 | 3799 | + | 756 | 252 | Lower collar protein | Staphylococcus phage SLPW] | 98.41 | YP_009278569.1 |
| ORF6 | 3812 | 5755 | + | 1,944 | 648 | Minor structural protein | Staphylococcus phage SAP-2 | 97.99 | YP_001491544.2 |
| ORF7 | 5767 | 6519 | + | 753 | 251 | Phage lysin | Staphylococcus phage GRCS | 97.60 | YP_009004305.1 |
| ORF8 | 6582 | 7478 | + | 897 | 299 | Tail fiber protein | Staphylococcus phage LSA2366 | 94.16 | QQO38231.1 |
| ORF9 | 7538 | 9301 | + | 1,764 | 588 | Tail fiber protein | Staphylococcus phage LSA2366 | 99.66 | QQO38232.1 |
| ORF10 | 9303 | 9722 | + | 420 | 140 | Holin | Staphylococcus phage LSA2366 | 100.00 | QQO38233.1 |
| ORF11 | 9700 | 11139 | + | 1,440 | 480 | Phage lysin | Staphylococcus phage Pabna | 99.37 | YP_009816546.1 |
| ORF12 | 11253 | 13538 | – | 2,286 | 762 | DNA polymerase | Staphylococcus phage SCH1 | 97.11 | YP_009787907.1 |
| ORF13 | 13554 | 14801 | – | 1,248 | 416 | DNA encapsidation protein | Staphylococcus phage Pabna | 99.52 | YP_009816544.1 |
| ORF14 | 14849 | 15328 | – | 480 | 160 | Hypothetical protein | Staphylococcus phage GRCS | 96.05 | YP_009004298.1 |
| ORF15 | 15331 | 15501 | – | 171 | 57 | Hypothetical protein | Staphylococcus phage S24-1 | 96.36 | YP_004957422.1 |
| ORF16 | 15494 | 15907 | – | 414 | 138 | Hypothetical protein | Staphylococcus phage BP39 | 91.97 | YP_009283981.1 |
| ORF17 | 15910 | 16089 | – | 180 | 60 | Hypothetical protein | Staphylococcus phage SLPW | 100.00 | YP_009278557.1 |
| ORF18 | 16140 | 16508 | – | 369 | 123 | Single-stranded DNA binding protein | Staphylococcus phage Pabna | 98.10 | NC_048107.1 |
| ORF19 | 16532 | 16768 | – | 237 | 79 | Hypothetical protein | Staphylococcus phage SLPW | 97.44 | YP_009278555.1 |
| ORF20 | 16786 | 17088 | – | 303 | 101 | Hypothetical protein | Staphylococcus phage SLPW | 99.00 | YP_009278554.1 |
| ORF21 | 17081 | 17278 | – | 198 | 66 | Hypothetical protein | Staphylococcus phage GRCS | 95.38 | YP_009004291.1 |
| ORF22 | 17282 | 17473 | – | 192 | 64 | Hypothetical protein | Staphylococcus phage BP39 | 87.76 | YP_009283976.1 |