
Download

ORIGINAL ARTICLE
Eriocalyxin B inhibits inflammation induced by CCI-induced microglia activation to relieve neuropathic pain through inhibition of JAK2/STAT3 and NF-κB pathways
Zhifeng Sheng, Xiaoyan Pan*
Department of Anesthesiology, Changzhou Traditional Chinese Medicine Hospital, Changzhou, Jiangsu, China
Abstract
Neuropathic pain is a very troublesome disease that seriously affects human life. Eriocalyxin B (EriB) has been revealed to attenuate various diseases through its anti-inflammatory effects, but its regulatory effects on neuropathic pain remains unclear. The paw withdrawal threshold and paw withdrawal thermal latency were detected through mechanical allodynia and thermal hyperalgesia tests. The spinal injury was assessed through hematoxylin and eosin staining. The cell apoptosis was measured through terminal deoxynucleotide transferase-mediated dUTP nick end-labeling assay. The protein expressions were examined through Western blot analysis. The mRNA expression was examined through reverse transcription-quantitative polymerase chain reaction. The ionized calcium-binding adaptor molecule 1 level in the spinal cord was evaluated through immunofluorescence assay. The levels of tumor necrosis factor-α, interleukin (IL)-1β, and IL-6 were measured through enzyme-linked-immunosorbent serologic assay. The chronic constriction injury (CCI) rat model was constructed for the study. Our results demonstrated that EriB relieved CCI-stimulated neuropathic pain and nerve damage. In addition, the enhanced neural apoptosis mediated by CCI induction was reduced after EriB treatment. In addition, EriB inhibited CCI-induced microglia activity and inflammation. At last, the Janus kinase 2–signal transducer and activator of transcription 3 (JAK2/STAT3) and nuclear factor kappa B (NF-κB) pathways were activated in CCI rat model, which were attenuated following EriB treatment. Importantly, EriB (10 mg/kg) had a strong effect that was similar to the positive control (1-μg/kg dexmedetomidine), suggesting that EriB may be an effective drug for neuropathic pain. This study demonstrated that EriB inhibited inflammation caused by CCI-induced microglia activation to relieve neuropathic pain through inhibition of JAK2/STAT3 and NF-κB pathways. This study may highlight the regulatory functions of EriB in the treatment of neuropathic pain.
Key words: EriB, inflammation, microglia activation, neuropathic pain, JAK2/STAT3 and NF-κB pathways
*Corresponding Author: Xiaoyan Pan, Department of Anesthesiology, Changzhou Traditional Chinese Medicine Hospital, No. 25, North Heping Road, Changzhou, Jiangsu, 213000, China. Email: [email protected]
Received: 30 January 2023; Accepted: 16 February 2023; Published: 20 April 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Neuropathic pain is a common and persistent disease (Baron et al., 2010; Devor, 1991). The International Association for the Study of Pain (IASP) has defined neuropathic pain as a disorder of the somatosensory nervous system that disrupts the central nervous system (CNS) and/or the peripheral nervous system (Bouhassira, 2019). The pathogenesis of neuropathic pain is complex, and some studies have proved that tissue damage is the direct consequence of neuropathic pain that influences the nervous system and stimulate the ectopic discharge phenomenon of bypass conduction (Cohen and Mao, 2014; Guo et al., 2022). Nevertheless, the mechanism of neuropathic pain has not been fully understood; hence, most of the current treatments are based on narcotic drugs, such as dexmedetomidine, to attenuate discomfort (Huang et al., 2017; Mücke et al., 2018; Xu. and Xu., 2021). Inflammation is a very remarkable feature of neuropathic pain; thus, reducing inflammation is the first step to relieve this disease. Therefore, seeking drugs that are more effective has become crucial.
In recent years, traditional Chinese medicine has played an increasingly important role in the treatment of neuropathic pain (Feng et al., 2014; Li et al., 2020). Eriocalyxin B (EriB) extracted from Isodon eriocalyx var is a biologically active ingredient. In traditional Chinese medicine, Isodon eriocalyx var has been used as a drug for anti-inflammatory treatment, and its extract EriB has also been developed as a drug for treating sore throats and inflammation (Leung et al., 2006; Niu et al., 2002). Studies have illustrated that EriB has anticancer effect, which can suppress cell proliferation, migration, invasion, and other malignant phenotypes in colon, pancreatic, and breast cancers (Duan et al., 2021; Li et al., 2012; Riaz et al., 2019). EriB exerts anti-inflammatory effects through inhibiting the differentiation of T helper 1 (Th1) and Th17 cells and the enhancement of reactive oxygen species (ROC), thereby improving autoimmune encephalomyelitis (Lu et al., 2013). Moreover, EriB exerts anti-inflammatory effects through selectively regulating the conversion of microglia to M2 phenotype (resolution of inflammation and tissue repair) by targeting the nuclear factor kappa B (NF-κB) signaling pathway, thereby alleviating Parkinson’s symptoms (Dou et al., 2018). However, the effects of EriB in neuropathic pain and its’ regulatory mechanism remain unclear.
Both Janus kinase 2–signal transducer and activator of transcription 3 (JAK2/STAT3) and NF-κB pathways have been revealed to participate into the progression of neuropathic pain (Fei et al., 2017; Popiolek-Barczyk and Mika, 2016; Song et al., 2021). However, whether EriB affects JAK2/STAT3 and NF-κB pathways to relieve neuropathic pain is unclear.
This study aimed to explore the regulatory role of EriB in the progression of neuropathic pain. Our study showed that EriB inhibited inflammation caused by chronic constriction injury (CCI)-induced microglia activation to relieve neuropathic pain through inhibition of JAK2/STAT3 and NF-κB pathways. These findings suggested that EriB could be a useful novel drug to treat neuropathic pain.
Materials and methods
CCI rat model
Male Sprague-Dawley (SD) mice (aged 6–8 weeks, n = 30) were acquired for experiments (Vital River, Beijing, China). Free food and water were supplied to animals, and they were kept at 25°C into a 12-h light/dark cycle. All animal experiments were performed in line with the guidelines of the Animal Experiments Ethics Committee of Changzhou Traditional Chinese Medicine Hospital.
Mice (n = 6 in each group) were randomly separated into the following five groups: Sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, and CCI+1-μg/kg dexmedetomidine. The CCI-induced rat model for neuropathic pain was created as described by Zhang et al. (2019). After anesthetizing with pentobarbital sodium (40 mg/kg), mice in the CCI group were subjected to expose sciatic nerves, and ligated with 4-0 chromic gut suture (Ethicon Inc., Cincinnati, USA), to induce CCI. EriB (5 or 10 mg/kg) or dexmedetomidine (1 μg/kg) was injected intraperitoneally to CCI mice for 7 days post-operation. Mice in the sham group were subjected to the same surgery but no sciatic nerve ligation was made. Finally, mice were euthanized with pentobarbital sodium (160 mg/kg). The L4–L6 spinal cord was removed from euthanized animals for additional experiments. EriB (B30248, purity >98%) was bought from Shanghai YuanYe Biotechnology Co (Shanghai, China).
Mechanical allodynia and thermal hyperalgesia tests
The paw withdrawal threshold (PWT) was measured through von Frey hair stimulation (Stoelting Co.) to assess mechanical allodynia (Inoue et al., 2009). von Frey filaments were put into the hind paw’s dorsal surface with sufficient force. When mice withdrew their paws, PWT was recorded in terms of pressure (g). Each trial was done for six times at 3-min interval.
The paw withdrawal thermal latency (PWL) was examined to evaluate thermal hyperalgesia (Zheng et al., 2019). Briefly, the infrared light beam from a modified Hargreaves device (Ugo Basile SRL) was irradiated to the hind paw’s plantar surface. Subsequently, when mice withdrew their paws, the PWL was recorded. All experimental mice underwent thermal and mechanical pain tests at 0, 3, 7, 14, and 21 days post-operation.
Hematoxylin and eosin (H&E) staining
The collected spinal cords were fixed in 4% paraformaldehyde. Next, the spinal cords embedded in paraffin and dehydrated, and cut into 5-µm sections. All sections were stained in H&E solution. Finally, histopathologic changes were observed under a light microscope.
The scoring criteria of spinal cord injury were as described by Shan et al. (2021) as follows:
0 = no lesion;
1 = gray matter containing 1–5 eosinophilic neurons;
2 = gray matter containing 6–10 eosinophilic neurons;
3 = gray matter containing >10 eosinophilic neurons;
4 = infarction of <1/3 of the gray matter area;
5 = infarction of 1/3–1/2 of the gray matter area; and
6 = infarction of >1/2 of the gray matter area.
Terminal deoxynucleotide transferase-mediated dUTP nick end-labeling (TUNEL) assay
The in situ cell death detection kit (Cat. No. 11684817910; Roche, Basel, Switzerland) was applied for measuring cell apoptosis. Sections of the spinal cord were permeabilized through Triton X-100 (0.1%) and sodium citrate. The sections were then incubated with TUNEL staining solution at 37°C in darkness for 1 h. After washing, incubation with converter-peroxidase (POD) and diaminobenzidine (DAB) was performed for spinal cord sections. 4',6-Diamidino-2-phenylindole (DAPI) was used for staining of nucleus. Eventually, the fluorescent images were developed under fluorescence microscope (Olympus, Tokyo, Japan).
Western blot analysis
Proteins were isolated from spinal cords through radioimmunoprecipitation (RIPA) lysis assay buffer (Thermo Fisher Scientific, MA). These proteins were separated through 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto polyvinylidene difluoride (PVDF) membranes (Beyotime, Shanghai, China). After blocking, the membranes were incubated at 4°C for 12 h with the following primary antibodies: B-cell lymphoma-2 (BCL-2, 1:1,000; ab196495; Abcam, Shanghai, China), Bcl-2-associated X (BAX 1:2,000; ab182733), cleaved caspase-3 (1:5,000; ab214430), cluster of differentiation (CD) 86 cells (1 µg/mL; ab112490), CD206 (1 µg/ml; ab64693), phospho Janus kinase 2 (p-JAK2, 1:1,000; ab32101), (JAK2, 1:5,000; ab108596), p-signal transducer and activator of transcription 3 (STAT3, 1:1,000; ab32143), STAT3 (1:1,000; ab68153), and β-actin (1 µg/mL; ab8226). Next, the appropriate secondary antibodies (1:2,000; ab7090; Abcam) were added to membranes. Ultimately, the chemiluminescence detection kit (Thermo Fisher Scientific Inc.) was used for evaluating the blots (Rahmati and Taherabadi, 2021).
Immunofluorescence (IF) staining
After washing, sections of the spinal cords were blocked in bovine serum albumin phosphate–buffered saline solution (BSA/PBS, 3%) and fetal bovine serum (FBS)/PBS (10%). Next, the sections were incubated with primary antibody of ionized calcium-binding adaptor molecule 1 (IBA1; 0.5 µg/mL, ab178846; Abcam) for 12 h at 4°C. The secondary antibodies (1:1,000, ab150080) were mixed with sections for 2 h post-washing. Fluorescence microscope (Olympus) was utilized for acquiring fluorescent images (Bostani et al., 2020; Rahmati and Rashno, 2021).
Enzyme Linked Immunosorbent Serologic Assay (ELISA)
The levels of tumor necrosis factor-α (TNF-α; ab236712), interleukin (IL)-1β (ab255730), and IL-6 (ab234570) in serum were examined by using commercial ELISA kits (Abcam) in line with the manufacturer’s instructions.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
The isolation of RNAs from the spinal cords of mice was conducted using the Trizol reagent (Termo Fisher, Waltham, MA). The synthesis from RNAs to complementary DNA (cDNA) was performed through the PrimeScript™ RT reagent kit (Takara, Dalian, China), and RT-qPCR was executed using the SYBR Green PCR kit (Toyobo, Japan). The calculation of mRNA expressions was done through the 2−ΔΔCt method, with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) acting as an internal control. The primers are listed in Table 1.
Table 1. The sequences of primers.
| TNF-α |
| F: 5’-GAA ACA CAC GAG ACG CTG AA-3’ |
| R: 5’-AGG GAG GCC TGA GAC ATC TT-3’ |
| IL-1β |
| F: 5’-TAC AGG CTC CGA GAT GAA CAA C-3’ |
| R: 5’-TTT GAG GCC CAA GGC CAC AG-3’ |
| IL-6 |
| F: 5’-CCA GAA ACC GCT ATG AAG TTC C-3’ |
| R: 5’-GTTGGGAGTGGTATCCTCTGTGA-3’ |
| GAPDH |
| F: 5’-ACC ACA GTC CAT GCC ATC AC-3 |
| R: 5’-TCC ACC ACC CTG TTG CTG TA-3’ |
Statistical analysis
GraphPad Prism 9.0 (GraphPad Software Inc.) was employed for statistical analysis. The data were presented as mean ± standard deviation (SD). Comparisons in the groups (two or multiple) were evaluated through the Student’s t-test or one-way ANOVA with Tukey’s post hoc test. Normality and variance homogeneity was checked for all cases. P < 0.05 was considered as statistically significant.
Results
EriB relieved CCI-stimulated neuropathic pain and nerve damage
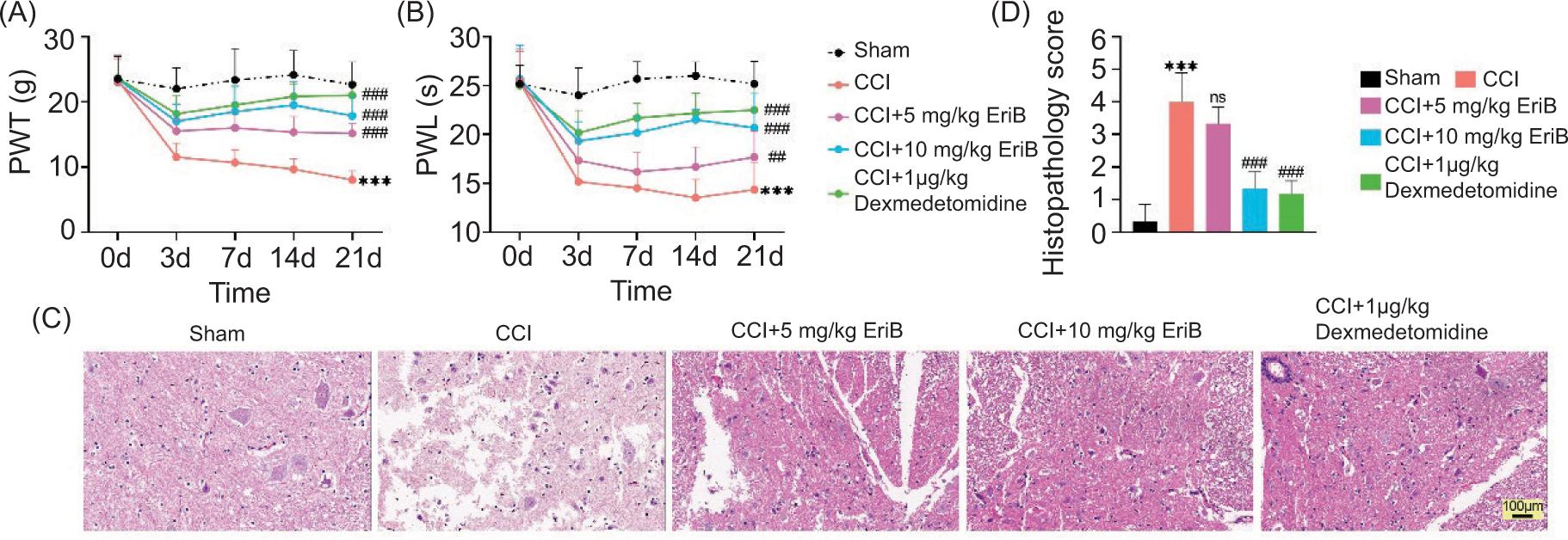
At first, the CCI rat model for neuropathic pain was constructed. In Figure 1A, the PWT decreased in CCI rat model, but this change reversed after treatment with EriB (5 mg/kg or 10 mg/kg) or dexmedetomidine (1 μg/kg). Moreover, the PWL demonstrated similar results (Figure 1B). H&E staining showed that nerve fiber swelling and fiber structure disorder were observed in the CCI model group, but they were restored after EriB or dexmedetomidine treatment (Figure 1C). In addition, histopathology score was strengthened in the CCI model group, but it was reduced following EriB or dexmedetomidine treatment (Figure 1D). Importantly, EriB (10 mg/kg) had a strong effect, which was similar to the positive control (1-μg/kg dexmedetomidine). These findings indicated that EriB relieved CCI-stimulated neuropathic pain and nerve damage.
Figure 1. EriB relieved CCI-stimulated neuropathic pain and nerve damage. Mice were divided into sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, CCI+1-μg/kg dexmedetomidine groups. (A) PWT was observed through mechanical allodynia test. (B) PWL was examined through thermal hyperalgesia test. (C) Spinal injury was assessed through H&E staining. (D) Histopathology score was also verified. ***P < 0.001 vs the sham group; ##P < 0.01, ###P < 0.001 vs the CCI group.
EriB suppressed CCI-mediated neural apoptosis
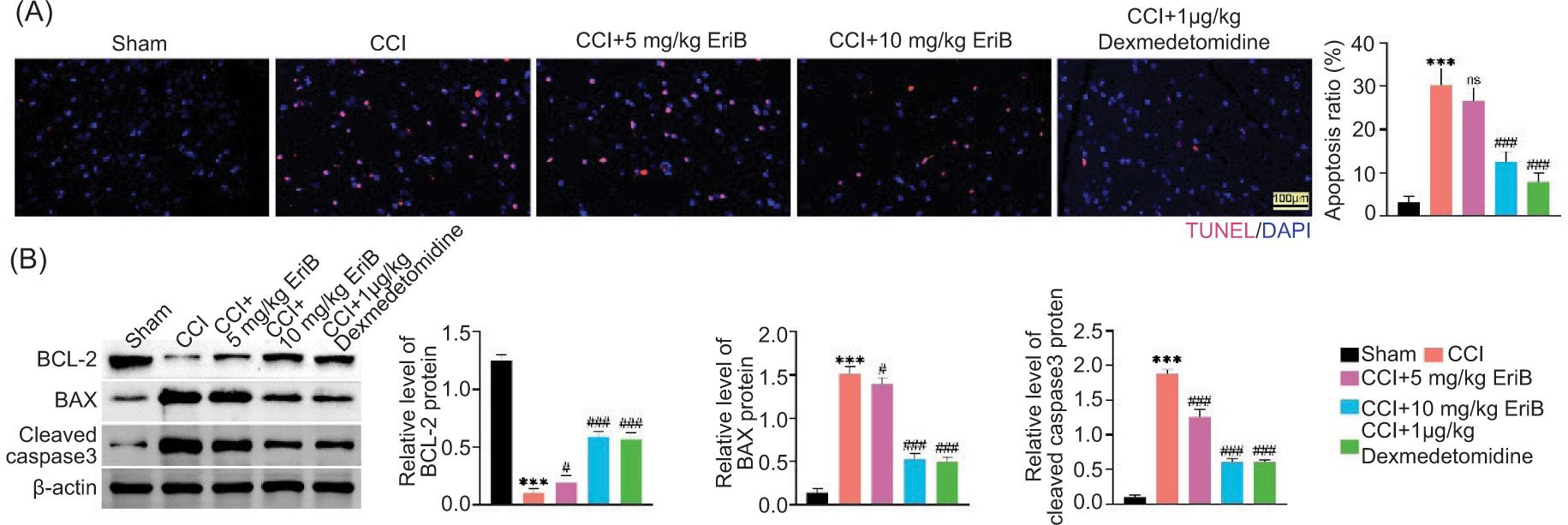
Cell apoptosis was enhanced in CCI rat model, but it was weakened after EriB or dexmedetomidine treatment (Figure 2A). In addition, the BCL-2 protein expression was decreased, while the BAX and cleaved caspase-3 protein expressions were increased in CCI rat model, but these changes were attenuated by EriB or dexmedetomidine treatment (Figure 2B). The effect of EriB (10 mg/kg) was similar to the positive control (1-μg/kg dexmedetomidine). Taken together, EriB suppressed CCI-mediated neural apoptosis.
Figure 2. EriB suppressed CCI-mediated neural apoptosis. Mice were divided into sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, and CCI+1-μg/kg dexmedetomidine groups. (A) Cell apoptosis in the spinal cord was measured by TUNEL assay. (B) The protein expressions of BCL-2, BAX, and Cleaved caspase-3 were assessed by Western blot analysis. ***P < 0.001 vs the sham group; #P < 0.05, ###P < 0.001 vs the CCI group.
EriB inhibited CCI-induced microglia activity
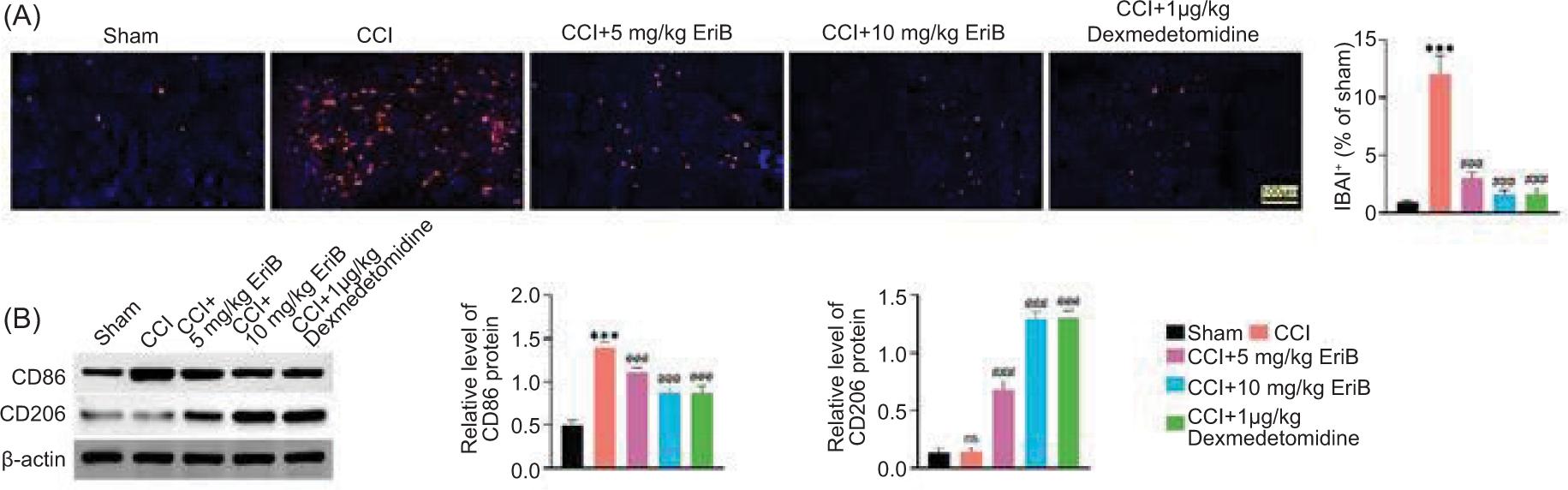
As displayed in Figure 3A, the IBA1 level was enhanced in CCI rat model, but this effect was reduced following EriB or dexmedetomidine treatment. In addition, the CD86 protein expression was increased in CCI rat model, but this change was weakened after EriB or dexmedetomidine treatment. The CD206 protein expression did not change after CCI induction and was increased after EriB or dexmedetomidine treatment (Figure 3B). Compared to the positive control (1-μg/kg dexmedetomidine), EriB (10 mg/kg) had the similar attenuated effects. These data confirmed that EriB inhibited CCI-induced microglia activity.
Figure 3. EriB inhibited CCI-induced microglia activity. Mice were divided into sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, and CCI+1-μg/kg dexmedetomidine groups. (A) IBA1 level in the spinal cord was evaluated through immunofluorescence assay. (B) The protein expressions of CD86 and CD206 were assessed by Western blot analysis. ***P < 0.001 vs the sham group; ###P < 0.001 vs the CCI group.
EriB reduced CCI-triggered inflammation
Results from ELISA indicated that the levels of TNF-α, IL-1β, and IL-6 were increased in CCI rat model, but these changes were attenuated following EriB or dexmedetomidine treatment (Figure 4A). In addition, the mRNA expressions of TNF-α, IL-1β, and IL-6 also had similar changes (Figure 4B). The remission effects of EriB (10 mg/kg) were similar to the positive control (1-μg/kg dexmedetomidine). In short, EriB reduced CCI-triggered inflammation.
Figure 4. EriB reduced CCI-triggered inflammation. Mice were divided into sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, and CCI+1-μg/kg dexmedetomidine groups. (A) Levels of TNF-α, IL-1β, and IL-6 in serum were measured by ELISA. (B) The mRNA expressions of TNF-α, IL-1β, and IL-6 were confirmed by RT-qPCR. ***P < 0.001 vs the sham group; ##P < 0.01, ###P < 0.001 vs the CCI group.
EriB retarded JAK2/STAT3 and NF-κB pathways
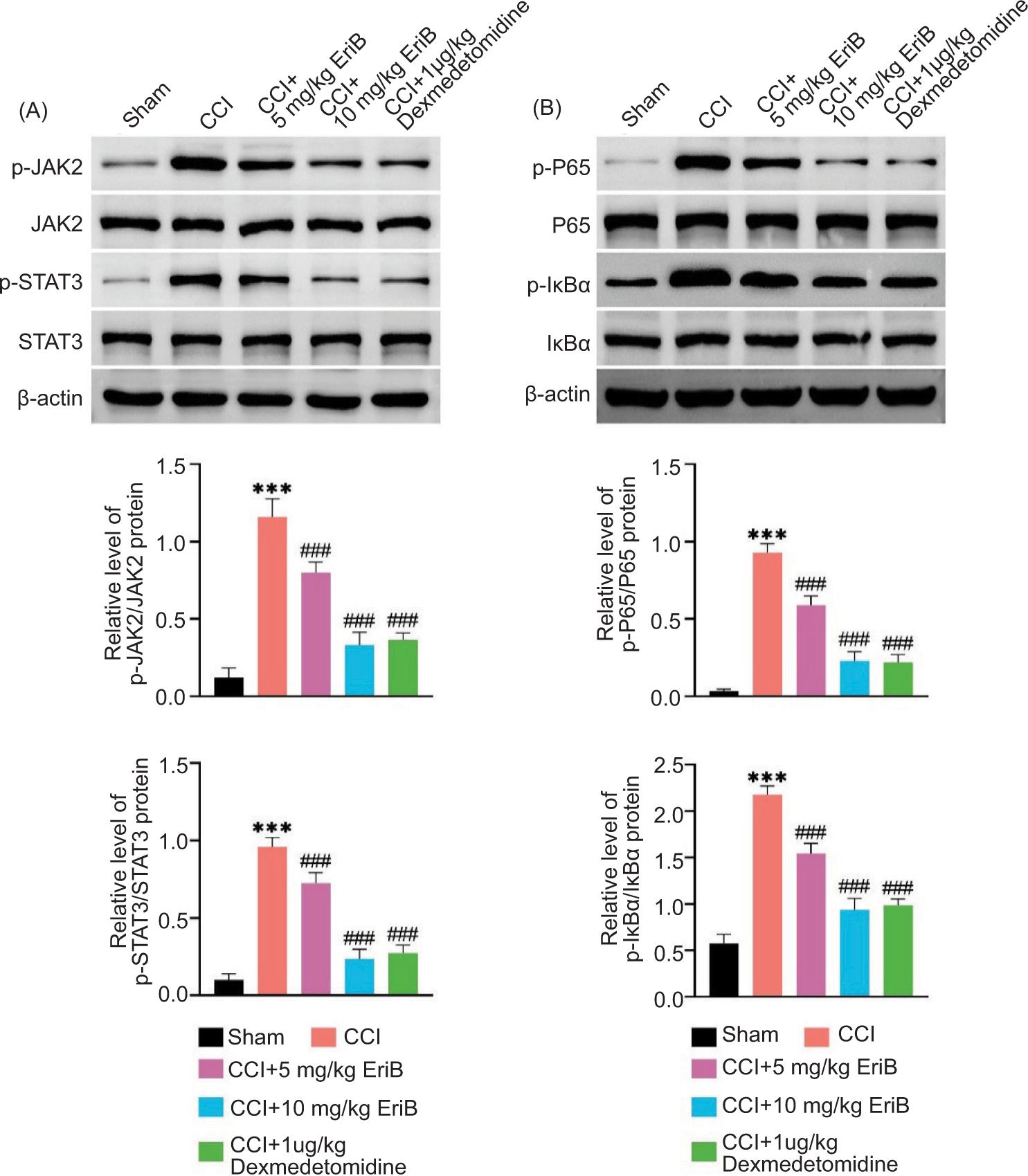
Finally, the effects of EriB were investigated on JAK2/STAT3 and NF-κB pathways. Figure 5A illustrated that p-JAK2/JAK2 and p-STAT3/STAT3 levels were increased in CCI rat model, but these effects were relieved after EriB or dexmedetomidine treatment. Similarly, the p-P65/P65 and p-IκBα/IκBα levels were enhanced in CCI rat model, but these changes were reversed after EriB or dexmedetomidine treatment (Figure 5B). Among them, EriB (10 mg/kg) had similar effect as that of positive control (1-μg/kg dexmedetomidine) on JAK2/STAT3 and NF-κB pathways. To sum up, EriB retarded JAK2/STAT3 and NF-κB pathways.
Figure 5. EriB retarded JAK2/STAT3 and NF-κB pathways. Mice were divided into sham, CCI, CCI+5-mg/kg EriB, CCI+10-mg/kg EriB, and CCI+1-μg/kg dexmedetomidine groups. (A) The protein expressions of p-JAK2, JAK2, p-STAT3, and STAT3 were assessed by Western blot analysis. (B) The protein expressions of p-P65, P65, p-IκBα, and IκBα were evaluated by Western blot analysis. ***P < 0.001 vs the sham group; ###P < 0.001 vs the CCI group.
Discussion
Neuropathic pain is caused by nerve inflammation, nerve damage, virus infection, and other factors (Gierthmühlen and Baron, 2016). More and more Chinese herb extracts have been investigated to ameliorate neuropathic pain (Luo et al., 2020; Xu et al., 2016). EriB was discovered to have regulatory effects on some diseases (Dou et al., 2018; Duan et al., 2021; Leung et al., 2006; Li et al., 2012; Lu et al., 2013; Riaz et al., 2019), but its roles in neuropathic pain remained unclear. In this study, a CCI rat model was constructed. Our results demonstrated that EriB relieved CCI-stimulated neuropathic pain and nerve damage. Moreover, the enhanced neural apoptosis mediated by CCI induction was reduced after EriB treatment.
Microglia are vital glial cells in the spinal cord that contribute to sensitize and maintain chronic pain (Prinz and Priller, 2014). Cortical and spinal microglial cells are key immune components with important functions in regulating inflammatory responses in the CNS Fujita and Yamashita, 2019; Streit, 1993). When immune homeostasis is disrupted, microglia changes into two extremely polarization states: M1 phenotype (pro-inflammatory) and M2 phenotype (Orihuela et al., 2016). Studies have revealed that activated microglia stimulates pro-inflammatory response and produces pro-inflammatory factors, such as TNF-α, IL-1β, and ROS, which additionally contribute to neuronal damage, thus inducing neuropathic pain (Wu et al., 2020; Zheng et al., 2019). In addition, a previous study has demonstrated that blocking of p38 mitogen-activated protein kinase (MAPK) and extracellular signal-regulated kinase (ERK) pathways could suppress the activity of microglia BV2, and decreased neuroinflammatory factors to relieve neuropathic pain (Choi et al., 2012). In this study, our experiments manifested that EriB inhibited CCI-induced microglia activity and inflammation.
The JAK2/STAT3 and NF-κB pathways also have been shown to take part into the regulation of neuropathic pain progression. For instance, red nucleus IL-6 modulates JAK/STAT3 and ERK signaling pathways to affect the maintenance of neuropathic pain (Ding et al., 2018). In addition, ligustrazine retards JAK/STAT3 pathway to relieve neuropathic pain (Wang et al., 2016). Astaxanthin suppresses MAPK and NF-κB pathways to decrease neuropathic pain (Zhao et al., 2021). Moreover, gallic acid represses the P2X7 receptor-mediated NF-κB/STAT3 pathways to mitigate neuropathic pain behaviors (Yang et al., 2021). Moreover, valproic acid affects STAT1/NF-κB and JAK2/STAT3 signaling pathways to regulate microglial function and reduce neuroinflammatory response, thereby relieving spinal nerve ligation-triggered neuropathic pain (Guo et al., 2021). In our study, the results showed that JAK2/STAT3 and NF-κB pathways were activated in CCI rat model, but these were attenuated following EriB treatment. Importantly, EriB (10 mg/kg) had the similar strong effects with the positive control (1-μg/kg dexmedetomidine), suggesting that EriB may be a useful drug to treat neuropathic pain.
Conclusion
This study demonstrated that EriB inhibited inflammation caused by CCI-induced microglia activation to relieve neuropathic pain through inhibition of JAK2/STAT3 and NF-κB pathways. However, this study also had some limitations for the regulatory functions of EriB on neuropathic pain progression, such as lacking other phenotypes and human samples. More experiments are required in the future to foster probe the other roles of EriB in neuropathic pain.
Availability of Data and Materials
All data generated or analyzed in this study are included in this published article. The datasets used and/or analyzed in the present study are available from the corresponding author on reasonable request.
Competing Interests
The authors stated that they had no conflict of interest to declare.
Author Contributions
Zhifeng Sheng designed the study, completed the experiment, and supervised data collection. Xiaoyan Pan analyzed and interpreted the data. Both authors prepared the manuscript for publication and reviewed its draft. Both authors read and approved the final manuscript.
REFERENCES
Baron, R., Binder, A. and Wasner, G., 2010. Neuropathic pain: diagnosis, pathophysiological mechanisms, and treatment. Lancet Neurology 9: 807–819. 10.1016/s1474-4422(10)70143-5
Bostani, M., Rahmati, M. and Mard, S.A., 2020. The effect of endurance training on levels of LINC complex proteins in skeletal muscle fibers of STZ-induced diabetic rats. Scientific Reports 10: 8738. 10.1038/s41598-020-65793-5
Bouhassira, D., 2019. Neuropathic pain: definition, assessment and epidemiology. Revue Neurologique (Paris) 175: 16–25. 10.1016/j.neurol.2018.09.016
Choi, D.C., Lee, J.Y., Lim, E.J., Baik, H.H., Oh, T.H. and Yune, T.Y., 2012. Inhibition of ROS-induced p38MAPK and ERK activation in microglia by acupuncture relieves neuropathic pain after spinal cord injury in rats. Experimental Neurology 236: 268–282. 10.1016/j.expneurol.2012.05.014
Cohen, S.P. and Mao, J., 2014. Neuropathic pain: mechanisms and their clinical implications. British Medical Journal 348: f7656. 10.1136/bmj.f7656
Devor, M., 1991. Neuropathic pain and injured nerve: peripheral mechanisms. British Medical Bulletin 47: 619–630. 10.1093/oxfordjournals.bmb.a072496
Ding, C.P., Guo, Y.J., Li, H.N., Wang, J.Y. and Zeng, X.Y., 2018. Red nucleus interleukin-6 participates in the maintenance of neuropathic pain through JAK/STAT3 and ERK signaling pathways. Experimental Neurology 300: 212–221. 10.1016/j.expneurol.2017.11.012
Dou, F., Chu, X., Zhang, B., Liang, L., Lu, G., Ding, J., et al. 2018. EriB-targeted inhibition of microglia activity attenuates MPP(+)-induced DA neuron injury through the NF-κB signaling pathway. Molecular Brain 11: 75. 10.1186/s13041-018-0418-z
Duan, D., Wang, Y., Jin, X., Li, M., Wang, L., Yan, Y., et al. 2021. Natural diterpenoid eriocalyxin B covalently modifies glutathione and selectively inhibits thioredoxin reductase inducing potent oxidative stress-mediated apoptosis in colorectal carcinoma RKO cells. Free Radical Biology and Medicine 177: 15–23. 10.1016/j.freeradbiomed.2021.10.013
Fei, R., Zhang, Y., Wang, S., Xiang, T. and Chen, W., 2017. α7 Nicotinic acetylcholine receptor in tumor-associated macrophages inhibits colorectal cancer metastasis through the JAK2/STAT3 signaling pathway. Oncology Reports 38: 2619–2628. 10.3892/or.2017.5935
Feng, L., Liu, W.K., Deng, L., Tian, J.X. and Tong, X.L., 2014. Clinical efficacy of aconitum-containing traditional Chinese medicine for diabetic peripheral neuropathic pain. American Journal of Chinese Medicine 42: 109–117. 10.1142/s0192415x14500074
Fujita, Y. and Yamashita, T., 2019. The effects of leptin on glial cells in neurological diseases. Frontiers in Neuroscience 13: 828. 10.3389/fnins.2019.00828
Gierthmühlen, J. and Baron, R., 2016. Neuropathic pain. Seminars in Neurology 36: 462–468. 10.1055/s-0036-1584950
Guo, C., Fan, Y., Kong, X. and Zhao, C., 2022. The effect of different water immersion strategies on delayed onset muscle soreness and inflammation in elite race walker. American Journal of Men’s Health 18(3): 64. 10.31083/j.jomh1803064
Guo, A., Li, J., Luo, L., Chen, C., Lu, Q., Ke, J., et al. 2021. Valproic acid mitigates spinal nerve ligation-induced neuropathic pain in rats by modulating microglial function and inhibiting neuroinflammatory response. International Immunopharmacology 92: 107332. 10.1016/j.intimp.2020.107332
Huang, X., Deng, R., Tu, W. and Hu, Z., 2017. Dexmedetomidine reduces neuropathic pain in a rat model of skin/muscle incision and retraction. Asian Journal of Surgery 40: 35–40. 10.1016/j.asjsur.2015.10.009
Inoue, N., Ito, S., Tajima, K., Nogawa, M., Takahashi, Y., Sasagawa, T., et al. 2009. Etodolac attenuates mechanical allodynia in a mouse model of neuropathic pain. Journal of Pharmacological Sciences 109: 600–605. 10.1254/jphs.08287fp
Leung, C.H., Grill, S.P., Lam, W., Gao, W., Sun, H.D., and Cheng, Y.C., 2006. Eriocalyxin B inhibits nuclear factor-kappaB activation by interfering with the binding of both p65 and p50 to the response element in a noncompetitive manner. Molecular Pharmacology 70: 1946–1955. 10.1124/mol.106.028480
Li, S.H., Li, L., Yang, R.N. and Liang, S.D., 2020. Compounds of traditional Chinese medicine and neuropathic pain. Chinese Journal of Natural Medicines (CJNM) 18: 28–35. 10.1016/s1875-5364(20)30002-9
Li, L., Yue, G.G., Lau, C.B., Sun, H., Fung, K.P., Leung, P.C., et al. 2012. Eriocalyxin B induces apoptosis and cell cycle arrest in pancreatic adenocarcinoma cells through caspase-and p53-dependent pathways. Toxicology and Applied Pharmacology 262: 80–90. 10.1016/j.taap.2012.04.021
Lu, Y., Chen, B., Song, J.H., Zhen, T., Wang, B.Y., Li, X., et al. 2013. Eriocalyxin B ameliorates experimental autoimmune encephalomyelitis by suppressing Th1 and Th17 cells. Proceedings of Natlional Academy of Sciences, USA 110: 2258–2263. 10.1073/pnas.1222426110
Luo, Y., Wang, C.Z., Sawadogo, R., Tan, T. and Yuan, C.S., 2020. Effects of herbal medicines on pain management. American Journal of Chinese Medicine 48: 1–16. 10.1142/s0192415x20500019
Mücke, M., Phillips, T., Radbruch, L., Petzke, F. and Häuser, W., 2018. Cannabis-based medicines for chronic neuropathic pain in adults. Cochrane database of systematic reviews 3: Cd012182. 10.1002/14651858.CD012182.pub2
Niu, X.M., Li, S.H., Mei, S.X., Na, Z., Zhao, Q.S., Lin, Z.W., et al. 2002. Cytotoxic 3,20-epoxy-ent-kaurane diterpenoids from Isodon eriocalyx var. laxiflora. Journal of Natural Products 65: 1892–1896. 10.1021/np020059e
Orihuela, R., McPherson, C.A. and Harry, G.J., 2016. Microglial M1/M2 polarization and metabolic states. British Journal of Pharmacology 173: 649–665. 10.1111/bph.13139
Popiolek-Barczyk, K. and Mika, J., 2016. Targeting the microglial signaling pathways: new insights in the modulation of neuropathic pain. Current Medicinal Chemistry 23: 2908–2928. 10.2174/0929867323666160607120124
Prinz, M. and Priller, J., 2014. Microglia and brain macrophages in the molecular age: from origin to neuropsychiatric disease. Nature Reviews Neuroscience 15: 300–312. 10.1038/nrn3722
Rahmati, M. and Rashno, A., 2021. Automated image segmentation method to analyse skeletal muscle cross section in exercise-induced regenerating myofibers. Scientific Reports 11: 21327. 10.1038/s41598-021-00886-3
Rahmati, M. and Taherabadi, S.J., 2021. The effects of exercise training on kinesin and GAP-43 expression in skeletal muscle fibers of STZ-induced diabetic rats. Scientific Reports 11: 9535. 10.1038/s41598-021-89106-6
Riaz, A., Rasul, A., Hussain, G., Zubair, M., Samiullah, K., Sarfraz, I., et al. 2019. Eriocalyxin B induces apoptosis in human triple negative breast cancer cells via inhibiting STAT3 activation and mitochondrial dysfunction. Pakistan Journal of Pharmaceutical Sciences 32: 2843–2848.
Shan, W., Liao, X., Tang, Y. and Liu, J., 2021. Dexmedetomidine alleviates inflammation in neuropathic pain by suppressing NLRP3 via Nrf2 activation. Experimental and Therapeutic Medicine 22: 1046. 10.3892/etm.2021.10479
Song, C., Adili, A., Kari, A. and Abuduhaer, A., 2021. FSTL1 aggravates sepsis-induced acute kidney injury through regulating TLR4/MyD88/NF-κB pathway in newborn rats. Signa Vitae 17: 167–173.
Streit, W.J., 1993. Microglial-neuronal interactions. Journal of Chemical Neuroanatomy 6: 261–266. 10.1016/0891-0618(93)90047-8
Wang, S., Li, A. and Guo, S., 2016. Ligustrazine attenuates neuropathic pain by inhibition of JAK/STAT3 pathway in a rat model of chronic constriction injury. Die Pharmazie 71: 408–412. 10.1691/ph.2016.6546
Wu, J., Wang, C. and Ding, H., 2020. LncRNA MALAT1 promotes neuropathic pain progression through the miR-154-5p/AQP9 axis in CCI rat models. Molecular Medicine Reports 21: 291–303. 10.3892/mmr.2019.10829
Xu, H.-l. and Xu, S.-y., 2021. Tacrine alleviates neuropathic pain in mice by mimicking the cell adhesion molecule L1. Signa Vitae 17: 140–150.
Xu, L., Zhang, Y. and Huang, Y., 2016. Advances in the treatment of neuropathic pain. Advances in Experimental Medicine and Biologyl 904: 117–129. 10.1007/978-94-017-7537-3_9
Yang, R., Li, Z., Zou, Y., Yang, J., Li, L., Xu, X., et al. 2021. Gallic acid alleviates neuropathic pain behaviors in rats by inhibiting P2X7 receptor-mediated NF-κB/STAT3 signaling pathway. Frontiers in Pharmacology 12: 680139. 10.3389/fphar.2021.680139
Zhang, W., Suo, M., Yu, G. and Zhang, M., 2019. Antinociceptive and anti-inflammatory effects of cryptotanshinone through PI3K/Akt signaling pathway in a rat model of neuropathic pain. Chemico-Biological Interactions 305: 127–133. 10.1016/j.cbi.2019.03.016
Zhao, L., Tao, X. and Song, T., 2021. Astaxanthin alleviates neuropathic pain by inhibiting the MAPKs and NF-κB pathways. European Journal of Pharmacology 912: 174575. 10.1016/j.ejphar.2021.174575
Zheng, Y., Hou, X. and Yang, S., 2019. Lidocaine potentiates SOCS3 to attenuate inflammation in microglia and suppress neuropathic pain. Cellular and Molecular Neurobiology 39: 1081–1092. 10.1007/s10571-019-00703-6