
Download
ORIGINAL ARTICLE
Cortex Mori extract inhibits migration and invasion of lung adenocarcinoma cells by blocking RECQL4-induced NF-κB and ERK signaling pathways
Qin Li, Enyao Wei*, Wenbin Zhang, Feng Zhang
Department of Pulmonary and Critical Care Medicine, Chongqing Hospital of Traditional Chinese Medicine, Chongqing, China
Abstract
Lung adenocarcinoma (LUAC) is one of the usual tumors of the lung with high mortality rate. RecQ-like helicase 4 (RECQL4) gene has been discovered to take part in the progression of different cancers by undertaking as an oncogene, and is relevant with poor prognosis of LUAC. Cortex Mori (CM) extract has been investigated to affect cellular progress to regulate different diseases. However, the detailed functioning of RECQL4 and CM extract, as well as their regulatory mechanisms in LUAC, has not been illustrated. The purpose of the present study was to probe the impact of RECQL4 and CM extract on progression of LUAC. The expression of RECQL4 in LUAC was assessed by The Cancer Genome Atlas (TCGA) database. The mRNA expression of RECQL4 was examined by real-time quantitative polymerase chain reaction. The protein expressions (epithelial–mesenchymal transition [EMT] process, nuclear factor kappa B [NF-κB] and extracellular signal-regulated kinase [ERK] signaling pathways-related proteins) were determined by Western blot analysis. The cell proliferation was tested through cell counting kit-8 assay. Cell migration and invasion was affirmed by wound-healing and transwell assays. The cell senescence was assessed through senescence-associated beta-galactosidase staining. The cell cycle was inspected by flow cytometry. Our findings demonstrated that RECQL4 exhibited higher expression in LUAC tissues and cell lines. Through functional experiments, we found that RECQL4 facilitated cell proliferation, migration, and invasion as well as EMT progression. In addition, RECQL4 relieved cell cycle arrest and cell senescence. Moreover, RECQL4 activated NF-κB and ERK signaling pathways by enhancing phospho(p)-p65–p65 and p-ERK–ERK levels in LUAC. CM extract exhibited antitumor effects in LUAC, and blocked RECQL4-induced NF-κB and ERK signaling pathways. Our results manifested that CM extract inhibited migration and invasion of LUAC cells by blocking RECQL4-induced NF-κB and ERK signaling pathways. This result could provide a promising therapeutic strategy for LUAC.
Key words: Cortex Mori (CM) extract, RECQL4, lung adenocarcinoma, NF-κB and ERK signaling pathways
*Corresponding author: Enyao Wei, Department of Pulmonary and Critical Care Medicine, Chongqing Hospital of Traditional Chinese Medicine, No. 40, Daomenkou, Chongqing 400010, Yuzhong District, China. Email: [email protected]
Received: 14 February 2023; Accepted: 28 February 2023; Published: 1 July 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Lung cancer, the most hackneyed type of cancer, is one of the leading causes of deaths globally (Nasim et al., 2019; Romaszko and Doboszyńska, 2018). In China, the occurrence and mortality rate because of lung cancer are rising rapidly and is on peak in both males and females (Jones and Baldwin, 2018; Wu et al., 2021). Non-small cell lung cancer (NSCLC) is the major type of lung cancer (about 85%) which includes lung adenocarcinoma (LUAC) and lung squamous cell carcinoma (LUSC) (Herbst et al., 2018). At present, the most effective treatment for early LUAC patients are mediastinal lymph node dissection and anatomical surgical resection (Denisenko et al., 2018). The advanced LUAC patients have diversified disease manifestations, and thus need complex multimodal treatments, including systemic treatment and local treatment (chemotherapy, radiotherapy, etc.; Succony et al., 2021). In recent years, although targeted therapies have been developed to prolong the survival period of cancer patients, the 5-year survival rate for lung cancer patients has been consistently low over the past 20 years (Hirsch et al., 2017; Lemjabbar-Alaoui et al., 2015). Therefore, finding new biological targets is essential for the early diagnosis and cure of lung cancer as well as enhancing the survival period of the patients.
RecQ-like helicase 4 (RECQL4) gene is a member of RECQ helicase family and combines in maintaining the genomes’ stability of nucleus and mitochondria (Balajee, 2021; Croteau et al., 2012). RECQL4 gene mutations are associated with three human autosomal recessive disorders, including Rothmund–Thomson syndrome, RAPADILLINO syndrome, and Baller–Gerold syndrome (Kitao et al., 1999; Siitonen et al., 2003; Van Maldergem et al., 2006). Recent reports have indicated that RECQL4 is highly expressed in gastric cancer and enhances cisplatin resistance through modulating the Ak strain transforming–Y box binding protein 1–multidrug resistance protein 1 (AKT–YB1–MDR1) signaling pathway (Mo et al., 2016). In addition, RECQL4 also exhibits higher expression in esophageal squamous cell carcinoma, modulates nuclear factor kappa B (NF-κB) and extracellular signal-regulated kinase (ERK) signaling pathways, and plays a pivotal role in regulating DNA damage responses, redox homeostasis, and cell survival (Lyu et al., 2021). RECQL4 stimulates ovarian cancer cell proliferation and invasion through regulating MAFB (MAF bZIP transcription factor B) gene expression (Guo et al., 2020). It has been illustrated that RECQL4 is in relation to poor prognosis of LUAC (Jiang et al., 2021). In the present study, through TCGA database, we discovered that the RECQL4 expression in LUAC tissues exists higher than that in normal tissues. Nonetheless, the roles of RECQL4 in LUAC remain unclear.
Therefore, this study mainly explored the role and specific regulatory mechanism of RECQL4 in LUAC. In conclusion, our findings unveiled that Cortex Mori (CM) extract inhibited migration and invasion of LUAC cells by blocking RECQL4-induced NF-κB and ERK signaling pathways. This study highlighted that RECQL4 may be a helpful bio-target for CM extract treatment in LUAC.
Materials and Methods
Bioinformatic analysis
The RECQL4 expression in normal or primary tumor tissues was analyzed from TCGA database through the UALCAN online platform (http://ualcan.path.uab.edu/). The survival rate of LUAC patients with low or high RECQL4 expression was evaluated through the Kaplan–Meier plotter database (https://kmplot.com/analysis/).
Cell lines and cell culture
The NSCLC cell lines (A549, PC-9, HCC827, and NCI-H2009) and human normal lung epithelial cell line (BEAS-2B) were purchased from the American Type Culture Collection (ATCC, Manassas, VA). RPMI-1640 medium (Procell Life Science & Technology, China) was supplemented with 10% fetal bovine serum (FBS; Procell), 1% GlutaMax (Procell), and 1% penicillin–streptomycin solution (100×; Procell). The cells were cultured in supplemented RPMI-1640 medium at 37°C with 5% CO2.
In the present study, CM (5 mg/mL; Tianjiang Pharmaceutical, China) was applied to treat A549 and PC-9 cells for 24 h.
Transfection
Small interfering RNA (siRNA) for knocking down of RECQL4 (si RECQL4) and its negative control (siNC) were acquired from GenePharma (Shanghai, China). The plasmid cloning (pc)DNA3.1 was designed to overexpress RECQL4 (RECQL4), and its negative control (pcDNA3.1) was also acquired in the same manner. The transfection of these plasmids in LUAC cells was done through Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA).
RECQL4 siRNA: 5'-CGGAAGAAAGGGGAGTGTTTTGG-3'. The sequence of the plasmid overexpression of RECQL4 is shown in Supplementary 1.
Real-time quantitative polymerase chain reaction (RT-qPCR)
TRIzol reagent (Invitrogen) was used to gain total RNA from LUAC cells. The reverse transcription of RNA to complementary DNA (cDNA) was performed through SuperScript™ II reverse transcriptase kit (Invitrogen). Next, RT-qPCR was carried out through the SYBR Premix Ex Taq™ (Takara, Shanghai, China). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was deemed as an internal reference, and the data quantification was performed with 2−ΔΔCt method.
The primer sequences (5'-3') were displayed as follows: RECQL4-forward: GAT CCT GGC TGG TTA CAG CG; RECQL4-reverse: AGT TGT GAT TCC TCT GAG CCT; GAPDH-forward: GTC AGC CGC ATC TTC TTT TG; and GAPDH-reverse: GCG CCC AAT ACG ACC AAA TC.
Western blot analysis
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) 10% was executed for the segregation of extracted proteins (40 µg), which were then placed to polyvinylidene difluoride (PVDF) membranes (Millipore). Next, after sealing, the membranes were mixed with primary antibodies overnight at 4°C against RECQL4 (1:1,000, ab188125, rabbit; Abcam, Cambridge, UK); E-cadherin (1:1,000, ab1416, rabbit); N-cadherin (1:1,000, ab76011, rabbit); snail (1:1,000, ab180714, rabbit); slug (1:1,000, ab180714, rabbit); vimentin (1:1,000, ab92547, rabbit); phospho(p)-p65 (1:1,000, ab76302, rabbit); p65 (1:1,000, ab16502, rabbit); phospho (p)-ERK (1:1,000, ab201015, rabbit); ERK (1:1,000, ab184699, rabbit); and GAPDH (1:1000, ab8245, mouse, the internal reference). Afterward, appropriate secondary antibodies (1:2,000, ab7090, goat anti-rabbit immunoglobulin G [IgG] H&L (HRP) pre-adsorbed; Abcam) were mixed into membranes at room temperature for 1 h incubation. Finally, the assessment of bands was done through chemiluminescence detection kit (Thermo Fisher Scientific, MA).
Cell Counting Kit-8 (CCK-8) assay
At first, the LUAC cells cultured in 96-well plate at 24, 48, and 72 h were mixed with CCK-8 reagent (20 μL; Sigma-Aldrich, St. Louis, MO). The assessment of the OD value was done at 450 nm, post-incubation of 4 h, under microplate reader (SpectraMax M5, Molecular Devices, San Jose, CA).
Wound healing assay
A549 and PC-9 cells were plated into a six-well plate. Next, a wound was created by tip of a 200-µL pipette. After gently rinsing with phosphate-buffered saline solution (PBS), the cells were mixed with serum-free medium. Images of the wound were obtained at 0 and 24 h under a light microscope (Olympus Corporation, Tokyo, Japan).
Transwell assay
The Matrigel, a gelatinous protein mixture (Becton Dickinson, USA)-coated Transwell chambers (Corning Life Sciences, Corning, NY) were applied for invasion assay. Briefly, cells (1×104) and Dulbecco’s modified eagle medium (DMEM) without FBS were placed in the upper chamber, and DMEM with 10% FBS was placed in the lower chamber. Then, the fixing (90% ethanol) and dyeing (0.1% crystal violet) of the invaded cells were performed. The invaded cells were counted through inverted light microscope (Olympus Corporation):
The cell invasion rate = (Invaded cells/Total cells) ×100%.
Flow cytometry (for cell cycle)
Cells were first fixed in 70% cold alcohol. After washing, staining of cells was performed with propidium iodide (PI; Beijing Solarbio Science & Technology, Beijing, China). The cell cycle phases (G1 [Gap 1 phase], S [synthesis phase], and G2 [Gap 2 phase]) were assessed under a FACS caliber flow cytometer (BD Bioscience, San Diego, CA).
Senescence-associated beta-galactosidase (SA-β-gal) staining
Fixing with 4% paraformaldehyde (Beyotime Institute of Biotechnology, Shanghai, China) was done for A549 and PC-9 cells. SA-β-gal activity was examined through β-galactosidase staining kit (Beyotime Institute of Biotechnology). At pH 6.0, 4-methylumbelliferyl β-D-galactopyranoside (MUG) was converted into 4-methylumbellifer-one (4-MU), a fluorescent hydrolysis product, and the SA-β-gal activity was verified to assess cell senescence. Images were observed using a light microscope (Olympus Corporation).
Statistical analysis
The SPSS 20.0 software was used for statistical analysis. All data were expressed as mean ± standard deviation (SD). Student’s t-test (between two groups) or one-way ANOVA (for multiple groups) was applied for statistical comparisons. All experiments were performed in triplicate; P < 0.05 was considered statistically significant.
Results
RECQL4 possessed higher expression in LUAC
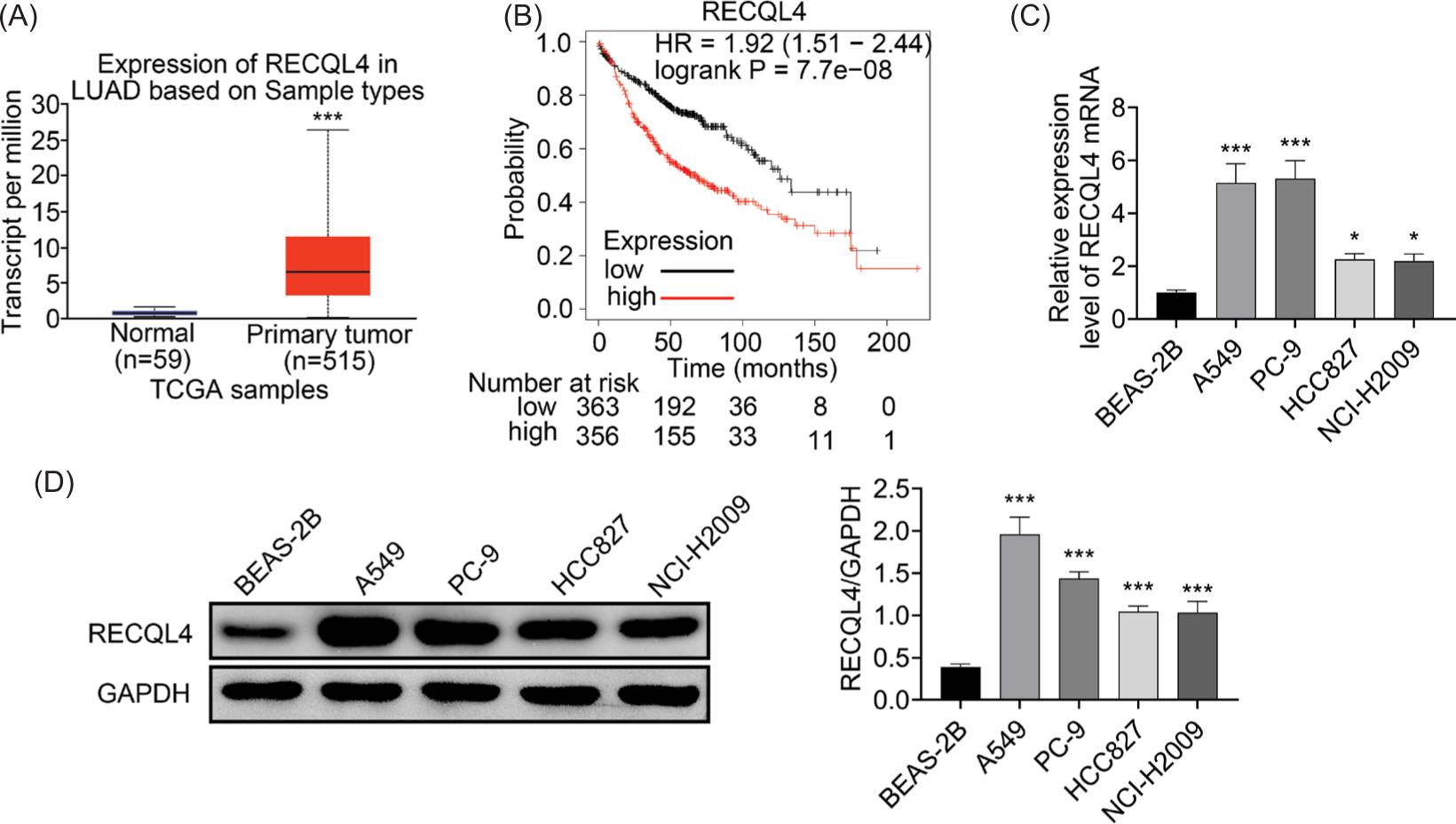
At first, it was discovered that RECQL4 exhibited higher expression in LUAC tissues in TCGA database through UALCAN online platform (http://ualcan.path.uab.edu/) (Figure 1a). Through the Kaplan–Meier plotter database (https://kmplot.com/analysis/), LUAC patients with high RECQL4 expression have worse survival rate than that of low RECQL4 expression (Figure 1b). Additionally, the enhanced mRNA expression of RECQL4 was uncovered in LUAC cell lines (A549, PC-9, HCC827, and NCI-H2009; Figure 1c). Similarly, the RECQL4 protein expression was also increased in LUAC cell lines (Figure 1D). To sum up, RECQL4 exhibited higher expression in LUAC.
Figure 1 RECQL4 exhibited higher expression in LUAC. (A) The expression of RECQL4 in LUAC was assessed by TCGA database. (B) The survival rate of LUAC patients with low or high RECQL4 expression was evaluated through Kaplan–Meier plotter database. (C) The mRNA expression of RECQL4 was tested through RT-qPCR in LUAC cell lines (A549, PC-9, HCC827, and NCI-H2009) and the normal lung epithelial cells (BEAS-2B). (D) The protein expression of RECQL4 was tested by Western blot analysis. *P < 0.05, ***P < 0.001.
RECQL4 enhanced cell proliferation
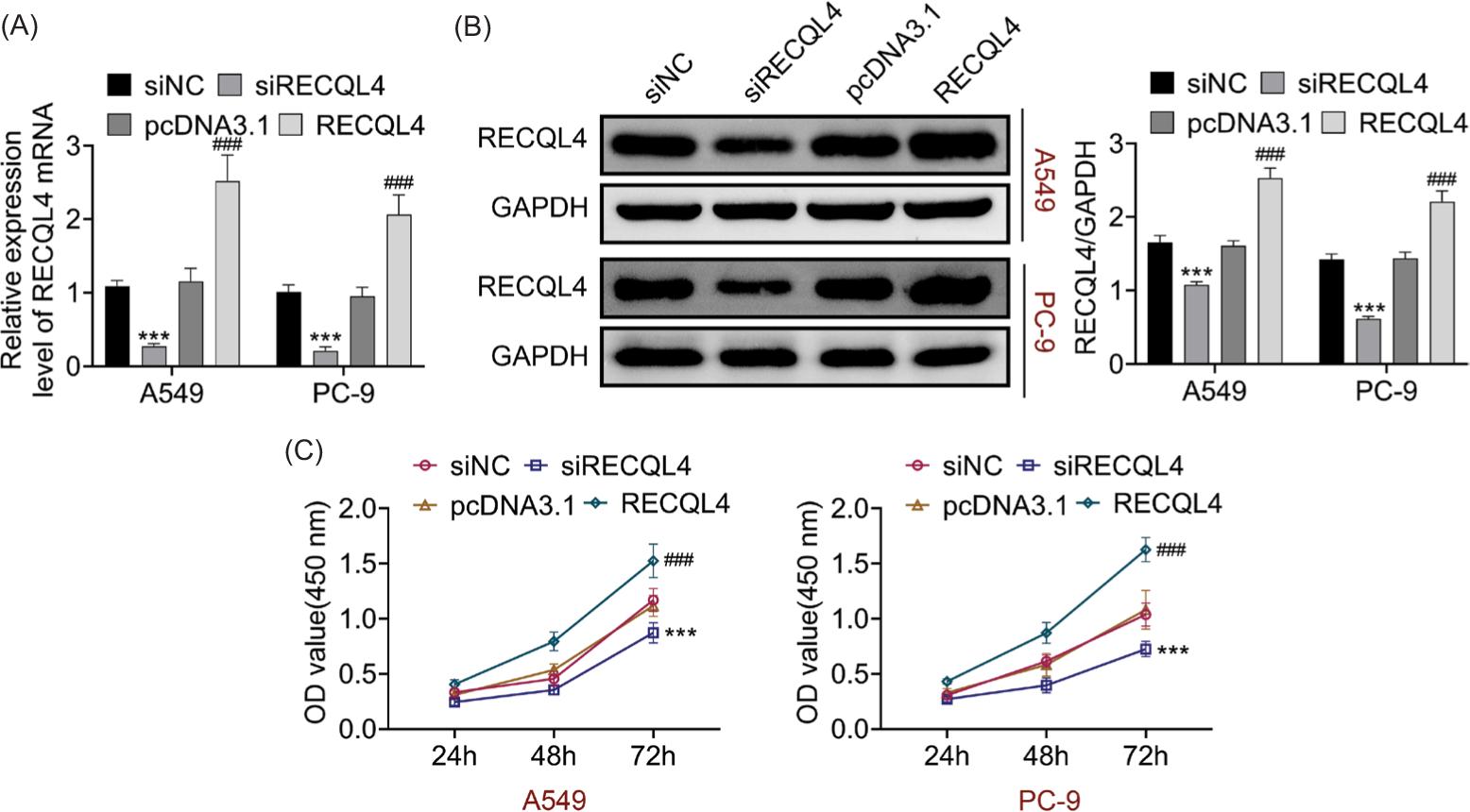
The knockdown and overexpression efficiencies of RECQL4 mRNA and protein levels were confirmed as displayed in Figures 2a and 2b. Furthermore, it was demonstrated that the cell viability decreased after silencing RECQL4 and increased after overexpressing RECQL4 (Figure 2c). This finding suggested that RECQL4 enhanced cell proliferation.
Figure 2 RECQL4 enhanced cell proliferation. (A,B) The knockdown and overexpression efficiency of RECQL4 in A549 and PC-9 cells were verified through RT-qPCR and Western blot analysis. (C) The cell proliferation was measured after silencing or up-regulating RECQL4 through CCK-8 assay. *P < 0.05, ***P < 0.001 vs siNC; ###P < 0.001 vs pcDNA3.1. NC: negative control.
RECQL4 facilitated cell migration and invasion as well as epithelial–mesenchymal transition (EMT) progression
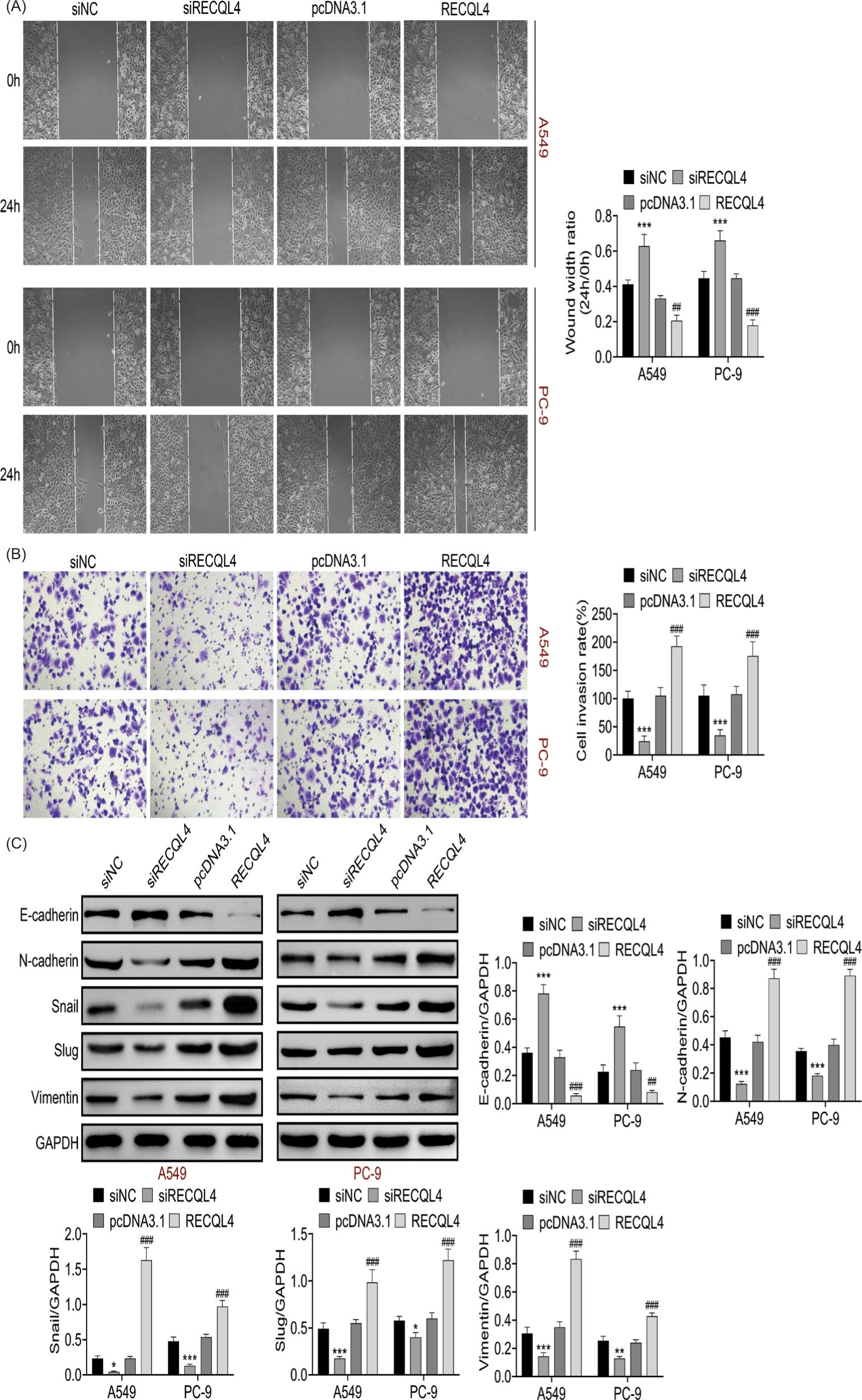
Findings revealed that both cell migration and invasion were suppressed by RECQL4 inhibition but strengthened by RECQL4 overexpression (Figures 3a and 3b). In addition, the E-cadherin protein expression was enhanced after repressing RECQL4, but it was reduced after overexpressing RECQL4. Meanwhile, the N-cadherin, snail, slug, and vimentin were decreased after RECQL4 suppression but increased after RECQL4 overexpression (Figure 3c). These data indicated that RECQL4 facilitated EMT progression.
Figure 3 RECQL4 facilitated cell migration and invasion as well as EMT progression. (A) The migration ability was measured after inhibiting or overexpressing RECQL4 through wound healing assay. (B) The invasion ability was discovered after RECQL4 knockdown or overexpression through transwell assay. (C) The EMT progression-related proteins (E-cadherin, N-cadherin, snail, slug, and vimentin) were inspected by Western blot analysis. *P < 0.05, **P < 0.01, ***P < 0.001 vs siNC; ##P < 0.01, ###P < 0.001 vs pcDNA3.1. NC: negative control.
RECQL4 relieved cell cycle arrest and cell senescence
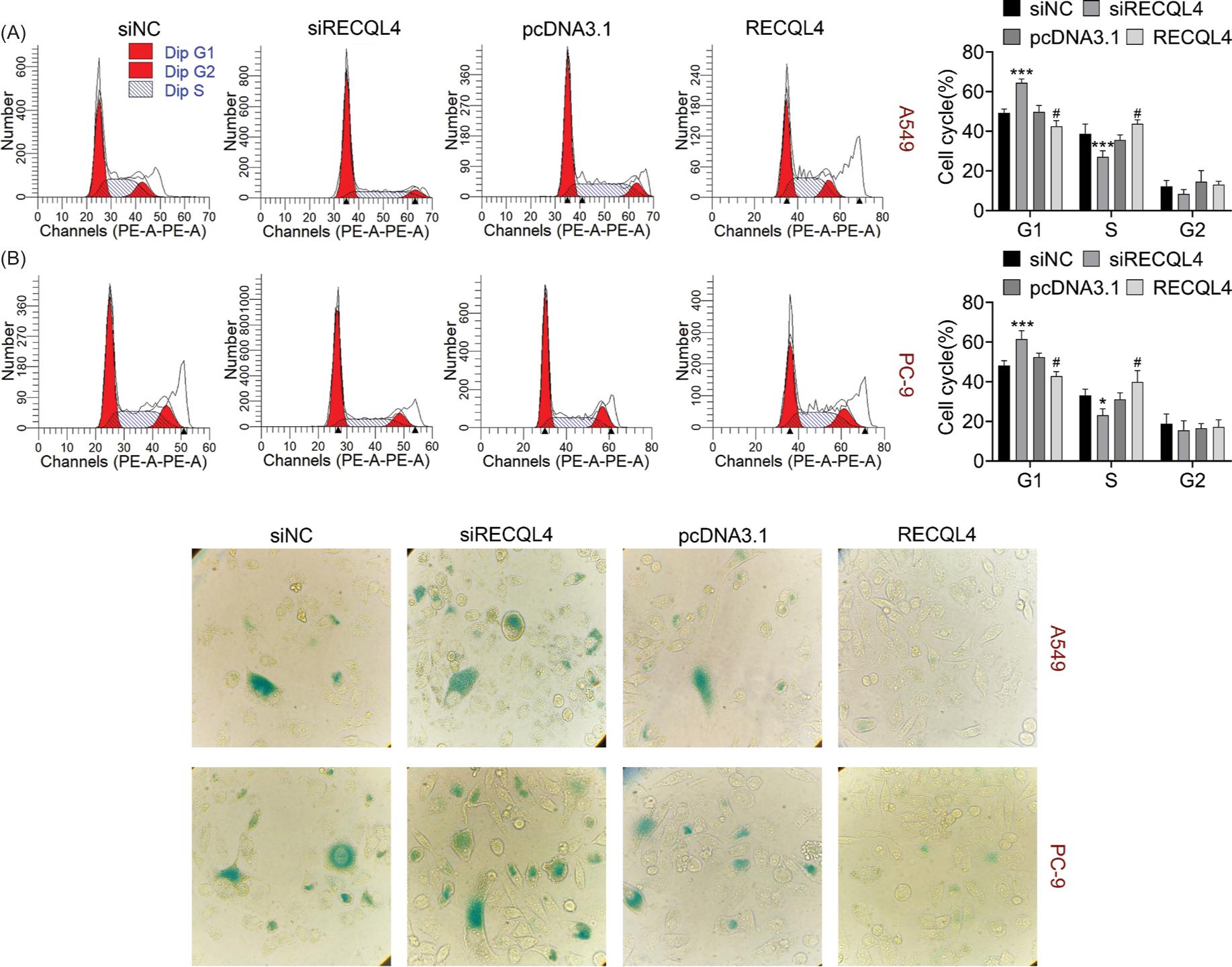
It was demonstrated by flow cytometry that the cell cycle was retarded at G1 phase after RECQL4 knockdown but promoted after RECQL4 overexpression (Figure 4a). In addition, cell senescence was heightened after inhibiting RECQL4 but retarded after overexpression of RECQL4 (Figure 4b). Taken together, RECQL4 relieved both cell cycle arrest and cell senescence.
Figure 4 RECQL4 relieved cell cycle arrest and cell senescence. (A) The cell cycle was evaluated after RECQL4 suppression or overexpression through flow cytometry. (B) The cell senescence was confirmed after RECQL4 down-regulation or up-regulation through SA-β-gal staining. *P < 0.05, ***P < 0.001 vs siNC; #P < 0.05 vs pcDNA3.1. NC: negative control.
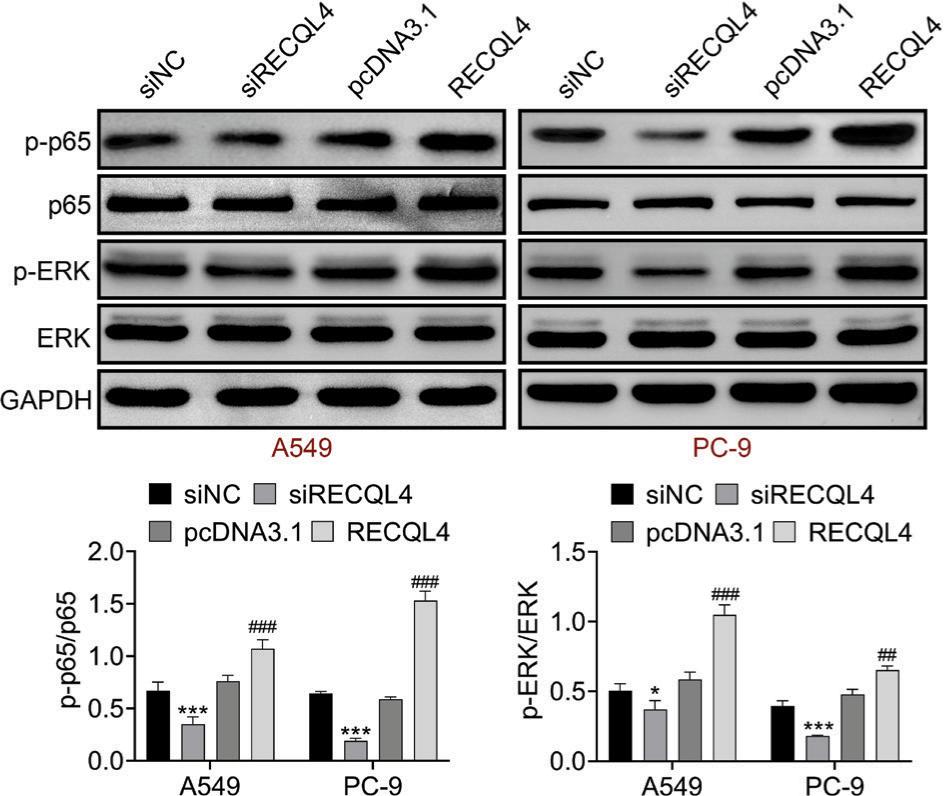
RECQL4 activated NF-κB and ERK signaling pathways
We probed whether RECQL4 modulated the NF-κB and ERK signaling pathways. Western blot analysis illuminated that the levels of phospho(p)-p65/p65 and p-ERK/ERK were reduced after RECQL4 repression but enhanced after RECQL4 overexpression (Figure 5). This data revealed that RECQL4 activated NF-κB and ERK signaling pathways in LUAC.
Figure 5 RECQL4 activated NF-κB and ERK signaling pathways. The protein expressions of p-p65, p65, p-ERK, and ERK were examined after silencing or overexpressing RECQL4 through Western blot analysis. *P < 0.05, ***P < 0.001 vs siNC; ##P < 0.01, ###P < 0.001 vs pcDNA3.1. NC: negative control.
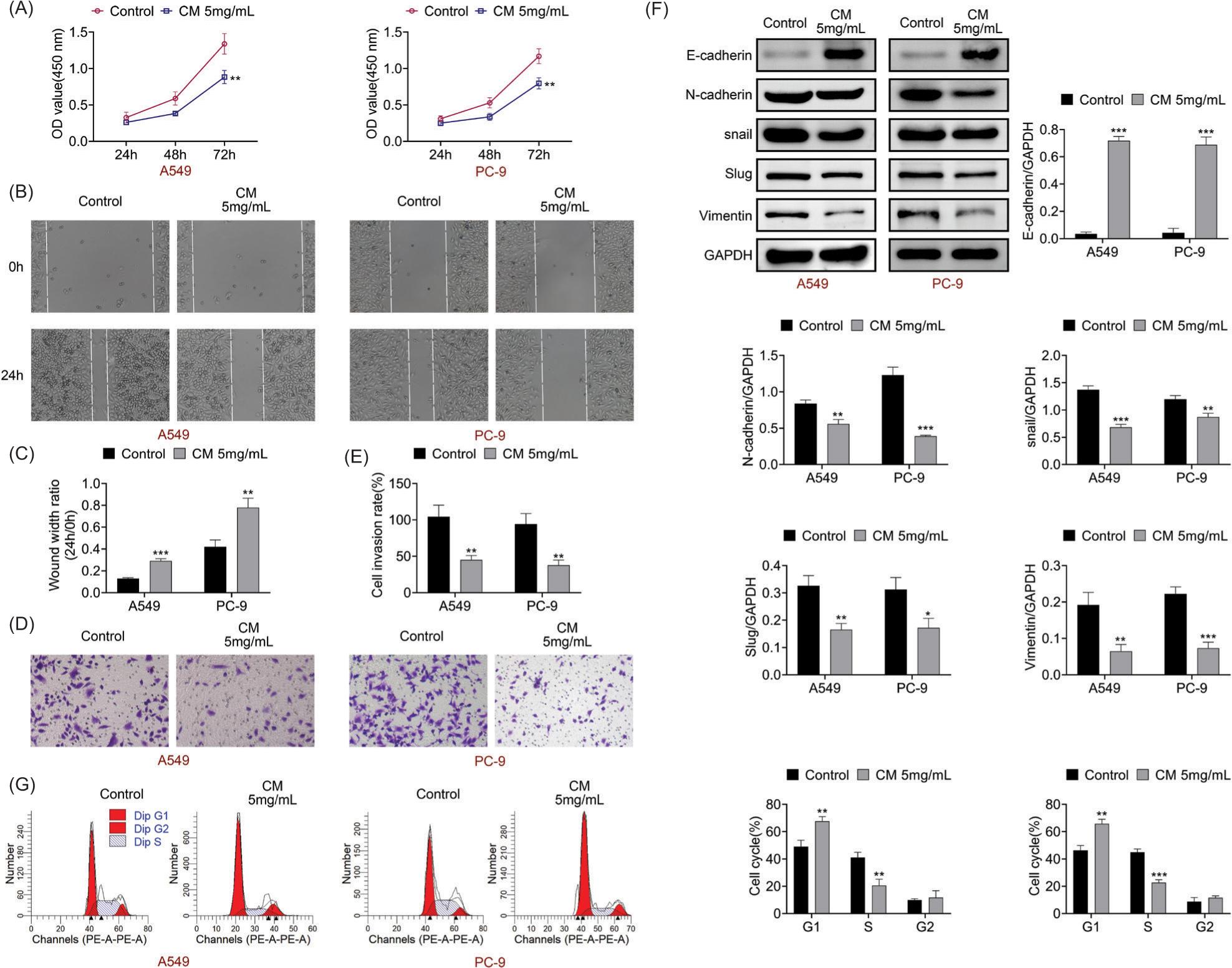
Cortex Mori extract exhibited antitumor effects in LUAC
It was demonstrated that CM (5 mg/mL) treatment reduced cell viability (Figure 6a). In addition, cell migration and invasion were weakened after CM (5 mg/mL) treatment (Figures 6b–6e). The E-cadherin protein expression was increased, but the N-cadherin, snail, slug and vimentin protein expressions were decreased after CM (5 mg/mL) treatment (Figure 6f). Besides, the S phase of cell cycle was retarded after CM (5 mg/mL) treatment (Figure 6g). In general, CM extract exhibited antitumor effects in LUAC.
Figure 6 Cortex Mori extract exhibited antitumor effects in LUAC. (A) Cell viability was measured after CM (5 mg/mL) treatment through CCK-8 assay. (B,C) Cell migration was tested after CM (5 mg/mL) treatment through wound healing assay. (D,E) Cell invasion was examined after CM (5 mg/mL) treatment through Transwell assay. (F) The protein expressions of E-cadherin, N-cadherin, snail, slug and vimentin were confirmed after CM (5 mg/mL) treatment through Western blot analysis. (G) The cell cycle was detected after CM (5 mg/mL) treatment through flow cytometry. *P < 0.05, **P < 0.01, ***P < 0.001 vs control.
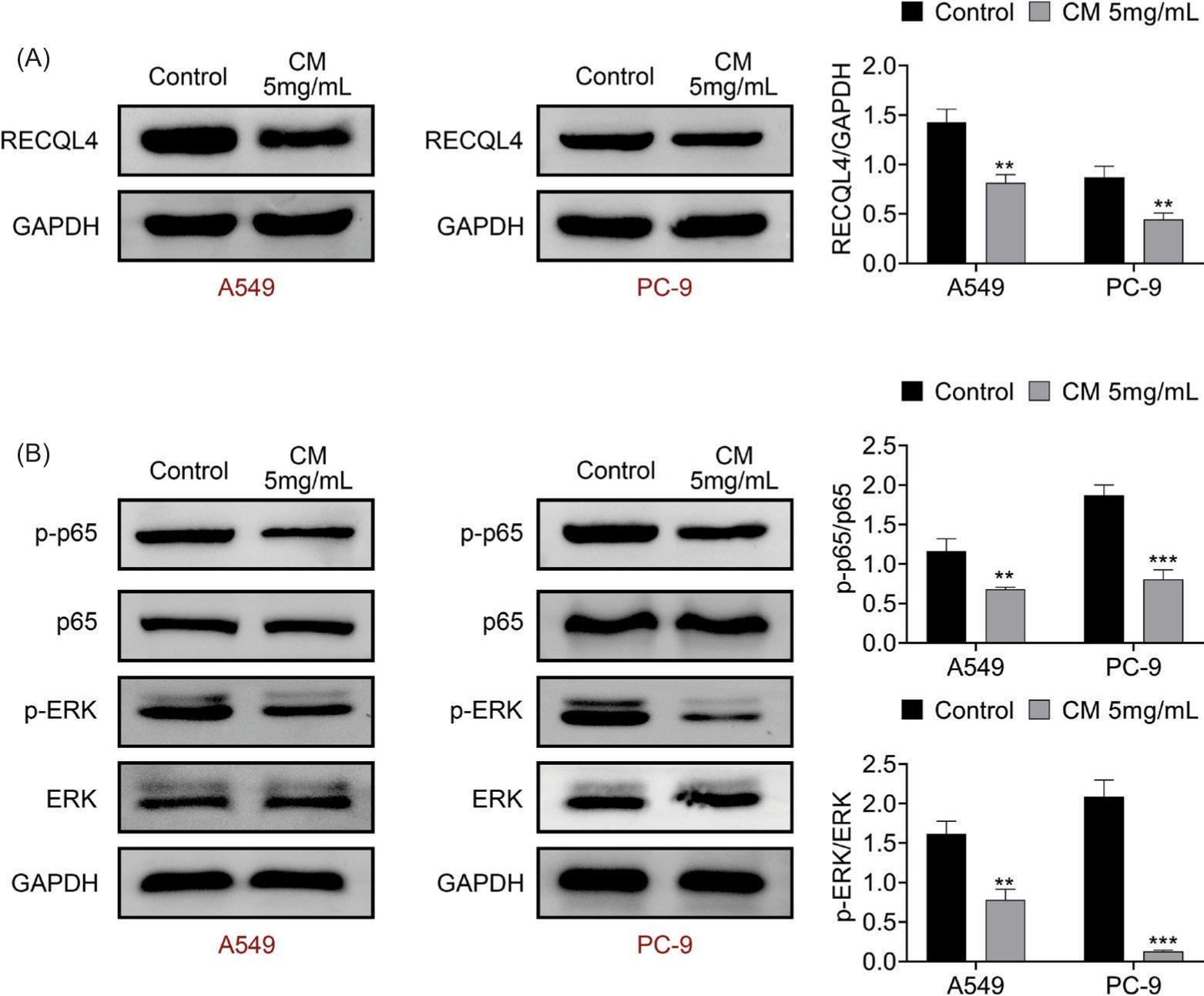
Cortex Mori extract blocked RECQL4-induced NF-κB and ERK signaling pathways
The RECQL4 protein expression was reduced after CM (5 mg/mL) treatment (Figure 7a). In addition, the levels of both p-p65/p65 and p-ERK/ERK were decreased following CM (5 mg/mL) treatment (Figure 7b). These findings indicated that CM extract blocked RECQL4-induced NF-κB and ERK signaling pathways.
Figure 7 Cortex Mori extract blocked RECQL4-induced NF-κB and ERK signaling pathways. (A) The protein expression of RECQL4 was tested after CM (5 mg/mL) treatment through Western blot analysis. (B) The protein expressions of p-p65, p65, p-ERK, and ERK were examined after CM (5 mg/mL) treatment through Western blot analysis. **P < 0.01, ***P < 0.001 vs control.
Discussion
RECQL4 has been shown to exhibit higher expressions and act as a facilitator in many types of cancers. For example, RECQL4 aggravates resistance to oxaliplatin by modulating PI3K/AKT signaling pathway in colon adenocarcinoma (Zhou et al., 2021)]. In hepatocellular carcinoma, the increased RECQL4 expression results in poor prognosis (Li et al., 2018a). Moreover, suppression of RECQL4 weakens cell invasion ability in metastatic prostate cancer (Su et al., 2010). In the present work, it was discovered that the higher expression of RECQL4 existed in LUAC tissues and cell lines.
In all, 90% cancer-related deaths are caused by metastasis, but this process is one of the most unintelligible pathological processes in development of cancers (Suhail et al., 2019). The development of novel treatment strategies for key factors driving metastasis is a very challenging objective for researchers (Chaffer and Weinberg, 2011).
In embryogenesis, the EMT process occurs in embryonic cells, resulting in epithelial cells to move to distant places (Nieto, 2013; Thiery et al., 2009). Once the migrated embryonic cells reach their destination, they undergo an opposite process, that is, the transformation from mesenchymal cells to epithelial cells (MET), so as to proliferate and differentiate into diverse organs (Nieto, 2013). Similarly, cancer cells also own metastatic process (Zhang and Weinberg, 2018). EMT progression provides cancerous epithelial cells the migration and invasion abilities, allowing them to enter the mesenchymal-like state (Bischoff, 2019). This change results in metastasis and colonization of primary tumor in distal organs, and forms secondary tumors (Piera-Velazquez and Jimenez, 2019) In addition, these cancer cells after EMT progression are discovered as frequently resistant to radiotherapy/chemotherapy drugs and are accompanied by tumor recurrence and re-metastasis (Du and Shim, 2016; Pastushenko and Blanpain, 2019). Understanding the molecular regulation mechanisms of EMT progression could provide novel targets and options for effective cancer treatment. In our study, through functional experiments, we found that RECQL4 facilitated cell proliferation and EMT progression. In addition, through flow cytometry and SA-β-gal staining assays, RECQL4 had been proved to relieve cell cycle arrest and cell senescence.
The NF-κB and ERK signaling pathways are highly active in diversified cancers, and aggravate tumor growth, metastasis, and EMT progression. For instance, BMP9 modulates the MAPK/ERK and NF-κB signaling pathways to suppress the proliferation and migration in LUAC (Wang et al., 2016). Osteopontin affects the PI3K, AKT, ERK, and NF-κB pathways to exacerbate cell migration in lung cancer (Fong et al., 2009). Perfluorooctanoic acid (PFOA) regulates the ERK/NF-κB/MMP-2/-9 pathways to induce cell metastasis in ovarian cancer (Li et al., 2018b). Puerarin retards the NF-κB and ERK signaling pathways to reduce cell migration and adhesion in LPS-mediated breast cancer (Liu et al., 2017). In our study, it was revealed that RECQL4 activated NF-κB and ERK signaling pathways by enhancing p-p65/p65 and p-ERK/ERK levels in LUAC.
Cortex Mori extract has been investigated to affect cellular progresses to regulate different diseases. For example, CM extract improves insulin resistance and nonalcoholic fatty liver disease in high-fat diet-/streptozotocin-stimulated type 2 diabetes (Ma et al., 2018). In addition, CM extract suppresses microtubule assembly to trigger cancer cell apoptosis (Nam et al., 2002). Besides, CM extract retards PI3K/AKT signaling to enhance apoptosis and reduce tumor invasion in melanoma (Hu et al., 2022). However, the detailed functions of RECQL4 and CM extract, as well as their regulatory mechanisms, in LUAC have not been illustrated. In this study, it was discovered that CM extract exhibited antitumor effects in LUAC, and blocked RECQL4-induced NF-κB and ERK signaling pathways.
In short, it has been discovered for the first time that CM extract inhibited migration and invasion of LUAC cells by blocking RECQL4-induced NF-κB and ERK signaling pathways. The discovery could help clarify the functioning of RECQL4 in improving the CM treatment of LUAC.
Nevertheless, this study also had some limitations concerning the effects of RECQL4 on CM treatment in LUAC progression, such as the functions of RECQL4 and CM extract in LUAC clinical samples, animal models and other cellular progression are not involved. In the future, more experiments are required to probe the other functions of CM extract, and its regulatory mechanism on RECQL4 in LUAC progression.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Competing interests
The authors stated that there was no conflict of interest to disclose.
Ethics approval
Not applicable.
Author Contributions
Qin Li and Enyao Wei designed the study, and supervised data collection. Wenbin Zhang analyzed and interpreted the data. Feng Zhang prepared the manuscript for publication and reviewed the draft of the manuscript. All authors read and approved the final manuscript.
REFERENCES
Balajee, A.S., 2021. Human RecQL4 as a novel molecular target for cancer therapy. Cytogenetic and Genome Research 161(6–7): 305–327. 10.1159/000516568
Bischoff, J., 2019. Endothelial-to-mesenchymal transition. Circulation Research 124(8): 1163–1165. 10.1161/CIRCRESAHA.119.314813
Chaffer, C.L. and Weinberg, R.A., 2011. A perspective on cancer cell metastasis. Science 331(6024): 1559–1564. 10.1126/science.1203543
Croteau D.L., Singh D.K., Hoh Ferrarelli L., Lu H., Bohr V.A. RECQL4 in genomic instability and aging. Trends in Genetics 28(12): 624–631. 10.1016/j.tig.2012.08.003
Denisenko, T.V., Budkevich, I.N. and Zhivotovsky, B., 2018. Cell death-based treatment of lung adenocarcinoma. Cell Death and Disease 9(2): 117. 10.1038/s41419-017-0063-y
Du, B. and Shim, J.S., 2016. Targeting epithelial-mesenchymal transition (EMT) to overcome drug resistance in cancer. Molecules 21(7): 965. 10.3390/molecules21070965
Fong Y.C., Liu S.C., Huang C.Y., Li T.M., Hsu S.F., Kao S.T., et al. 2009. Osteopontin increases lung cancer cells migration via activation of the alphavbeta3 integrin/FAK/Akt and NF-kappaB-dependent pathway. Lung Cancer 64(3): 263–270. 10.1016/j.lungcan.2008.09.003
Guo L., Li Y., Zhao C., Peng J., Song K., Chen L., et al. 2020. RECQL4, negatively regulated by miR-10a-5p, facilitates cell proliferation and invasion via MAFB in ovarian cancer. Frontiers in Oncology 10: 524128. 10.3389/fonc.2020.524128
Herbst, R.S., Morgensztern, D. and Boshoff, C., 2018. The biology and management of non-small cell lung cancer. Nature 553(7689): 446–454. 10.1038/nature25183
Hirsch F.R., Scagliotti G.V., Mulshine J.L., Kwon R., Curran W.J., Jr., Wu Y.L., et al. 2017. Lung cancer: current therapies and new targeted treatments. Lancet 389(10066): 299–311. 10.1016/S0140-6736(16)30958-8
Hu X., Zhang K., Pan G., Wang Y., Shen Y., Peng C., et al. 2022. Cortex mori extracts induce apoptosis and inhibit tumor invasion via blockage of the PI3K/AKT signaling in melanoma cells. Frontiers in Pharmacology 13: 1007279. 10.3389/fphar.2022.1007279
Jiang W., Xu J., Liao Z., Li G., Zhang C., Feng Y. 2021. Prognostic signature for lung adenocarcinoma patients based on cell-cycle-related genes. Frontiers in Cell and Developmental Biology 9: 655950. 10.3389/fcell.2021.655950
Jones, G.S. and Baldwin, D.R., 2018. Recent advances in the management of lung cancer. Clinical Medicine (London) 18(Suppl 2): s41–s46. 10.7861/clinmedicine.18-2-s41
Kitao S., Lindor N.M., Shiratori M., Furuichi Y., Shimamoto A., 1999. Rothmund–Thomson syndrome responsible gene, RECQL4: genomic structure and products. Genomics 61(3): 268–276. 10.1006/geno.1999.5959
Lemjabbar-Alaoui H., Hassan O.U., Yang Y.W., Buchanan P., 2015. Lung cancer: biology and treatment options. Biochimica et Biophysica Acta 1856(2): 189–210. 10.1016/j.bbcan.2015.08.002
Li, Jin., Liao, Dang., Chen, Wu, Liao., 2018a. Upregulation of RECQL4 expression predicts poor prognosis in hepatocellular carcinoma. Oncology Letters 15(4): 4248–4254. 10.3892/ol.2018.7860
Li X., Bao C., Ma Z., Xu B., Ying X., Liu X., et al. 2018b. Perfluorooctanoic acid stimulates ovarian cancer cell migration, invasion via ERK/NF-κB/MMP-2/-9 pathway. Toxicology Letters 294: 44–50. 10.1016/j.toxlet.2018.05.009
Liu X., Zhao W., Wang W., Lin S., Yang L., 2017. Puerarin suppresses LPS-induced breast cancer cell migration, invasion and adhesion by blockage NF-κB and Erk pathway. Biomedicine & Pharmacotherapy 92: 429–436. 10.1016/j.biopha.2017.05.102
Lyu G., Su P., Hao X., Chen S., Ren S., Zhao Z., et al. 2021. RECQL4 regulates DNA damage response and redox homeostasis in esophageal cancer. Cancer Biology & Medicine 18(1): 120–138. 10.20892/j.issn.2095-3941.2020.0105
Ma L.L., Yuan Y.Y., Zhao M., Zhou X.R., Jehangir T., Wang F.Y., et al. 2018. Mori cortex extract ameliorates nonalcoholic fatty liver disease (NAFLD) and insulin resistance in high-fat-diet/streptozotocin-induced type 2 diabetes in rats. Chinese Journal of Natural Medicine 16(6): 411–417. 10.1016/S1875-5364(18)30074-8
Mo D., Fang H., Niu K., Liu J., Wu M., Li S., et al. 2016. Human helicase RECQL4 drives cisplatin resistance in gastric cancer by activating an AKT-YB1-MDR1 signaling pathway. Cancer Research 76(10): 3057–3066. 10.1158/0008-5472.CAN-15-2361
Nam S.Y., Yi H.K., Lee J.C., Kim J.C., Song C.H., Park J.W., et al. 2002. Cortex mori extract induces cancer cell apoptosis through inhibition of microtubule assembly. Archives of Pharmacal Research 25(2): 191–196. 10.1007/BF02976562
Nasim F., Sabath B.F., Eapen G.A., 2019. Lung cancer. Medical Clinics of North America 103(3): 463–473. 10.1016/j.mcna.2018.12.006
Nieto, M.A., 2013. Epithelial plasticity: a common theme in embryonic and cancer cells. Science 342(6159): 1234850. 10.1126/science.1234850
Pastushenko, I. and Blanpain, C., 2019. EMT transition states during tumor progression and metastasis. Trends in Cellular Biology 29(3): 212–226. 10.1016/j.tcb.2018.12.001
Piera-Velazquez, S. and Jimenez, S.A., 2019. Endothelial to mesenchymal transition: role in physiology and in the pathogenesis of human diseases. Physiological Reviews 99(2): 1281–1324. 10.1152/physrev.00021.2018
Romaszko, A.M. and Doboszyńska, A., 2018. Multiple primary lung cancer: a literature review. Advances in Clinical and Experimental Medicine 27(5): 725–730. 10.17219/acem/68631
Siitonen H.A., Kopra O., Kääriäinen H., Haravuori H., Winter R.M., Säämänen A.M., Peltonen L., et al. 2003. Molecular defect of RAPADILINO syndrome expands the phenotype spectrum of RECQL diseases. Human Molecular Genetics 12(21): 2837–2844. 10.1093/hmg/ddg306
Su Y., Meador J.A., Calaf G.M., Proietti De-Santis L., Zhao Y., Bohr V.A., et al. 2010. Human RecQL4 helicase plays critical roles in prostate carcinogenesis. Cancer Research 70(22): 9207–9217. 10.1158/0008-5472.CAN-10-1743
Succony L., Rassl D.M., Barker A.P., McCaughan F.M., Rintoul R.C., 2021. Adenocarcinoma spectrum lesions of the lung: detection, pathology and treatment strategies. Cancer Treatment Reviews 99: 102237. 10.1016/j.ctrv.2021.102237
Suhail Y., Cain M.P., Vanaja K., Kurywchak P.A., Levchenko A., Kalluri R., et al. 2019. Systems biology of cancer metastasis. Cell Systems 9(2): 109–127. 10.1016/j.cels.2019.07.003
Thiery J.P., Acloque H., Huang R.Y., Nieto M.A., 2009. Epithelial-mesenchymal transitions in development and disease. Cell 139(5): 871–890. 10.1016/j.cell.2009.11.007
Van Maldergem L., Siitonen H.A., Jalkh N., Chouery E., De Roy M., Delague V., et al. 2006. Revisiting the craniosynostosis-radial ray hypoplasia association: Baller–Gerold syndrome caused by mutations in the RECQL4 gene. Journal of Medical Genetics 43(2): 148–152. 10.1136/jmg.2005.031781
Wang J., Weng Y., Zhang M., Li Y., Fan M., Guo Y., et al. 2016. BMP9 inhibits the growth and migration of lung adenocarcinoma A549 cells in a bone marrow stromal cell-derived microenvironment through the MAPK/ERK and NF-κB pathways. Oncology Reports 36(1): 410–418. 10.3892/or.2016.4796
Wu, F., Wang, L. and Zhou, C., 2021. Lung cancer in China: current and prospect. Current Opinion in Oncology 33(1): 40–46. 10.1097/CCO.0000000000000703
Zhang, Y. and Weinberg, R.A., 2018. Epithelial-to-mesenchymal transition in cancer: complexity and opportunities. Frontiers in Medicine 12(4): 361–373. 10.1007/s11684-018-0656-6
Zhou F., Wang L., Jin K., Wu Y., 2021. RecQ-like helicase 4 (RECQL4) exacerbates resistance to oxaliplatin in colon adenocarcinoma via activation of the PI3K/AKT signaling pathway. Bioengineered 12(1): 5859–5869. 10.1080/21655979.2021.1964156