
Download
Original Article
Combining network pharmacology and bioinformatics to identify bioactive compounds and potential mechanisms of action of Sedum aizoon L in the treatment of atherosclerosis
Bo Jie Zhu1#, Guan Ye Nai2#, Tian Xiao Pan1, Zhuo Fei Ma3*, Wei Jie Zhou4*
1Department of Chinese Medicine, the People’s Hospital of Baise, Baise, China;
2Department of Hematology, The Affiliated Hospital of Youjiang Medical University for Nationalities, Baise, China;
3School of Dentistry, The Youjiang Medical University for Nationalities, Baise, China;
4Department of Clinical Laboratory, the People’s Hospital of Baise, Baise, China
#These authors contributed equally to this work.
Abstract
Sedum aizoon L (SL) is a medicinal plant containing several active components with anti-inflammatory, hemostatic, and blood pressure lowering effects. The aim of this research was to investigate the main pathways, mechanisms, and active components of SL to treat atherosclerosis (AS) through network pharmacology. The active ingredients and their targets of action were obtained by setting the active ingredient-screening conditions using SL as a keyword in the Traditional Chinese Medicine System Pharmacology Database and Analysis Platform. The differentially expressed genes related to AS were obtained from the Gene Expression Omnibus database, and the targets related to the treatment of AS were retrieved from databases, such as DisGeNet and GENECARDs, and the targets of AS and SL were intersected using the Cytoscape software platform and applied to construct a drug–compound–target pathway network map. Gene ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses, and protein–protein interaction were performed to explore the mechanisms of action of SL against AS. In all, 12 active ingredients were screened from the chemical composition of SL, among which myricetin, oleanolic acid, ursolic acid, sitosterol, and beta-sitosterol were the major active ingredients for the anti-atherosclerotic effect of SL. Combining the active ingredient–target network and disease–target protein–protein interaction (PPI) network, GO and KEGG analysis, tumor necrosis factor signaling pathway, and interleukin-17 signaling pathway were the key pathways of action. SL acts as an anti-atherosclerotic agent through multiple chemical components, targets, and pathways. The active ingredients of SL mainly play the role of prevention and treatment of AS by inhibiting inflammatory response, as an antioxidant, and by lowering blood lipids, thereby providing the theoretical basis for its clinical use.
Key words: Sedum aizoon L, atherosclerosis, network pharmacology
*Corresponding Authors: Zhuo Fei Ma, School of Dentistry, The Youjiang Medical University for Nationalities, Baise, China. Email: [email protected]; Wei Jie Zhou, Department of Clinical Laboratory, the People’s -Hospital of Baise, Baise, China. Email: [email protected]
Received: 1 June 2023; Accepted: 3 July 2023; Published: 31 August 2023
© 2023 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Cardiovascular disease is currently one of the leading causes of human mortality, and atherosclerosis (AS) is the pathophysiological basis and etiology of all cardiovascular diseases (Humphries et al., 2018; Liu et al., 2019; Sun et al., 2018; Troidl et al., 2020; Xi et al., 2021). AS is a slowly developing, complex inflammatory vascular disease with arterial wall thickening, loss of elasticity, sclerosis, and luminal narrowing as its main features, and atherosclerotic plaques deposited in the intima being its key clinical pathological basis (Kobiyama and Ley, 2018). According to the statistical data, the incidence of AS is increasing and trends are toward younger generation (Keeter et al., 2022; Wu et al., 2019). Scientific and effective prevention and treatment of AS has become the key to prevent and treat cardiovascular diseases. Statins and anti-platelet aggregation drugs are commonly used as anti-AS drugs. Although they are effective in preventing and treating AS, their long-term use can lead to adverse effects, such as liver failure and kidney damage, rhabdomyolysis, and acute renal failure (Lee et al., 2019; Ott et al., 2015; Panes et al., 2017). Therefore, mining novel drugs for the pathogenesis of AS has significant clinical value for the prevention and treatment of AS.
In recent years, traditional Chinese medicine (TCM) and its active ingredients have become a hot spot for anti-AS drug research because of their mild effects, low toxicity, multiple pathways, and multiple targets in the prevention and treatment of AS and related cardiovascular and cerebrovascular diseases (Song et al., 2021). Researchers have now elucidated that andrographolide and berberine may exert their anti-AS effects through nuclear factor kappa B/CCAAT/enhancer-binding protein beta/peroxisome proliferator-activated receptor gamma (NF-κB/CEBPB/PPARG) signaling pathway, and mitogen-activated protein kinase (MAPK) and phosphoinositide-3-kinase–Ak strain transforming (PI3K/Akt) signaling pathway, providing a theoretical basis for the treatment of AS by TCM (Shi et al., 2022; Xie et al., 2020). The study by Zhang et al. (2018) revealed phenylpropanoids, flavonoids, terpenoids, and alkaloids via the protection of vascular endothelial cells from oxidative stress activation of Nrf2/HO-1 signaling (35024180). Other research showed that berberine protected from carotid atherosclerosis via PI3K/AKTmTOR signaling pathway34592881. Sedum aizoon L (SL) is a perennial herb of the genus Sedum belonging to the family Sedum. It contains a large number of saprophytic, phenolic, and flavonoid substances (Wang et al., 2021). Current studies have shown that SL has anti-inflammatory, hemostatic, antibacterial, and blood pressure-lowering effects, in addition to in vitro inhibition of tumor cell proliferation and regulation of oxidative stress in the body (Li et al., 2017; Wang et al., 2021; Xu et al., 2015). Combined with the above findings, it is suggested that SL has a potential anti-AS clinical value and deserves to be explored in depth.
Network pharmacology has become a popular tool for drug research in recent years, and its systematic and holistic features provide new ideas for the study of complex drug systems widely used to screen active ingredients and elucidate mechanisms of action etc. to provide technical support for rational clinical drug use and promote the application and development of drugs, especially in TCM, where significant results have been achieved (Wu et al., 2018; Zhou et al., 2020). Therefore, in this research, we attempted to elucidate the targets, active ingredients, and possible mechanisms of SL for treating AS.
Method
Active ingredients of Sedum aizoon L
We used the Traditional Chinese Medicine System Pharmacology Database (TCMSP) database to search for the main active ingredients of SL and obtain main target genes for the action of active ingredients. Finally, we used the Cytoscape software platform (version 3.9.0) to visualize main active ingredients and their targets. The shades of color and the size of graphs were used to visualize connection between active ingredients and targets. The shades of color and the size of graphs were determined by the connectivity of active ingredients and target.
Therapeutic targets of atherosclerosis
We searched DisGeNet (repositories of human gene–disease associations; https://www.disgenet.org/) and GENECARDs (human genes and model orthologues; https://www.genecards.org/) databases with the keyword “atherosclerosis.” We searched therapeutic targets for AS using the keyword “atherosclerosis.” Subsequently, AS-related databases were searched from the Gene Expression Omnibus (GEO) database, and GSE28829 and GSE43292 were included. We then normalized the data set using “Limma” (“linear models for microarray data”) package in R language and searched for differentially expressed genes (DEGs); the criteria had a fold change of more than 1, with P < 0.05, and included them in the follow-up study.
SL potential therapeutic targets
We obtained DEGs from the GEO database, and then obtained AS-related targets with DisGeNet and GENECARDs databases, and at least two or three intersections were considered as AS-related therapeutic targets. The AS-related therapeutic targets were then intersected with the therapeutic targets of Jing tian Panax notoginseng (Chinese ginseng), and the intersected genes were the AS-related targets of Jing tian Panax notoginseng.
Protein–protein interaction (PPI), Kyoto Encyclopedia of Genes and Genomes, and gene ontology (GO) analysis
We obtained the target genes associated with Jing tian Panax notoginseng for AS treatment, and to reveal the functions of the target genes, we performed GO annotation and KEGG pathway enrichment analysis using the R package clusterProfiler. GO terms consisted of the following: biological process (BP), cellular component (CC), and molecular function (MF); q < 0.05 was considered as statistically significant. The target genes were imported into the Search Tool for the Retrieval of Interacting Genes (STRING) database (https://string-db.org/) to construct a functional PPI network. Interaction score > 0.4 was fixed. Finally, we used Cytoscape (version 3.9.0) to visualize main active ingredients and their targets of action, with the shade of color and the size of the graph determined by the connectivity of active ingredients and the targets of action.
Molecular docking
First, the active ingredients from the core targets of SL for AS treatment were identified. Two-dimensional (2D) structures were retrieved from the PubChem website and converted to the lowest energy 3D structures using ChemBio3D. Then, the 3D structures of the core targets were obtained from the Protein Data Bank (PDB) database, and the “PyMOL” software was used to remove water and ligands. Next, we used the “AutoDockTools” software to convert proteins and drug components into Protein Data Bank, Partial Charge (Q), & Atom Type (T) (PDBQT) format files to identify active pockets. The AutoDock Vina software was used to perform molecular docking. Finally, the “PyMOL” (an open source but proprietary molecular visualization system) software was used to visualize docking results.
Identification of core active ingredients and their mechanisms of action
We performed intersection analysis of the therapeutic targets of active ingredients and AS. The most -overlapping target genes were considered as core ingredients to treat AS and were included in the subsequent analysis. We continued to construct PPIs using STRING and Cytoscape in the same way as done earlier. Overlapping genes were also subjected to GO and KEGG using the R language to assess their potential mechanism to treat AS.
Statistical analysis
Data were mainly obtained by downloading from public databases. Data were considered statistically significant for differential gene identification with a fold change of more than 1, with P < 0.05, and for GO and KEGG analysis, with q < 0.05.
Results
Active components and targets of Sedum aizoon L
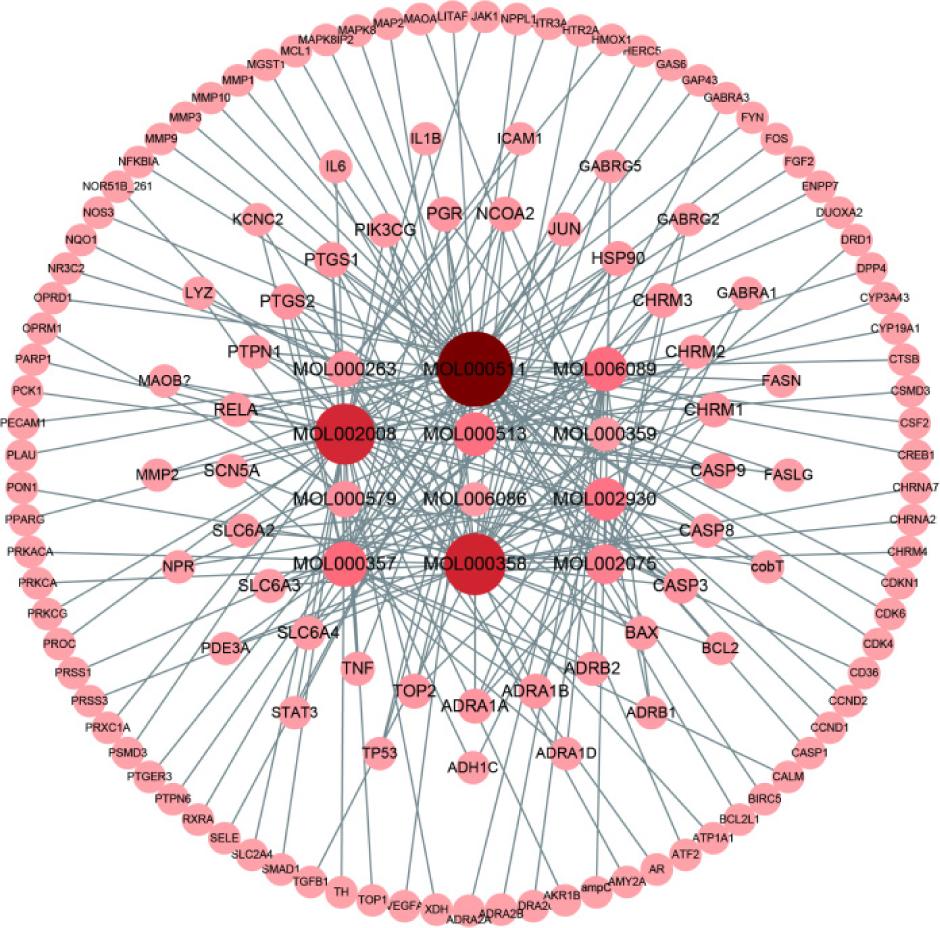
According to predetermined criteria, 134 therapeutic targets and 12 active ingredients of SL were acquired from TCMSP, and the regulatory network of active ingredients and targets of SL were constructed by using Cytoscape. As shown in Figure 1, the center circle is the active ingredient of SL and the surrounding circles are the targets. This shows that the top three active ingredients are ursolic acid, beta-sitosterol, and myricetin (Table 1).
Figure 1. Targets and active ingredients of SL.
Table 1. Active ingredients of SL.
| MOL ID | Active Ingredients |
|---|---|
| MOL002008 | Myricetin |
| MOL002075 | 1-[(2S)-1-Methyl-2-piperidyl]acetone |
| MOL000263 | Oleanolic acid |
| MOL002930 | Tyrosol |
| MOL000357 | Sitogluside |
| MOL000358 | Beta-sitosterol |
| MOL000359 | Sitosterol |
| MOL000511 | Ursolic acid |
| MOL000513 | 3,4,5-Trihydroxybenzoic acid |
| MOL000579 | Hydroquinone |
| MOL006086 | Isomyricitrin |
| MOL006089 | (1R)-2-[(2S)-1-Methyl-2-piperidyl]-1-phenyl-ethanol |
MOL: molecular.Supplementary
Target screening of atherosclerosis
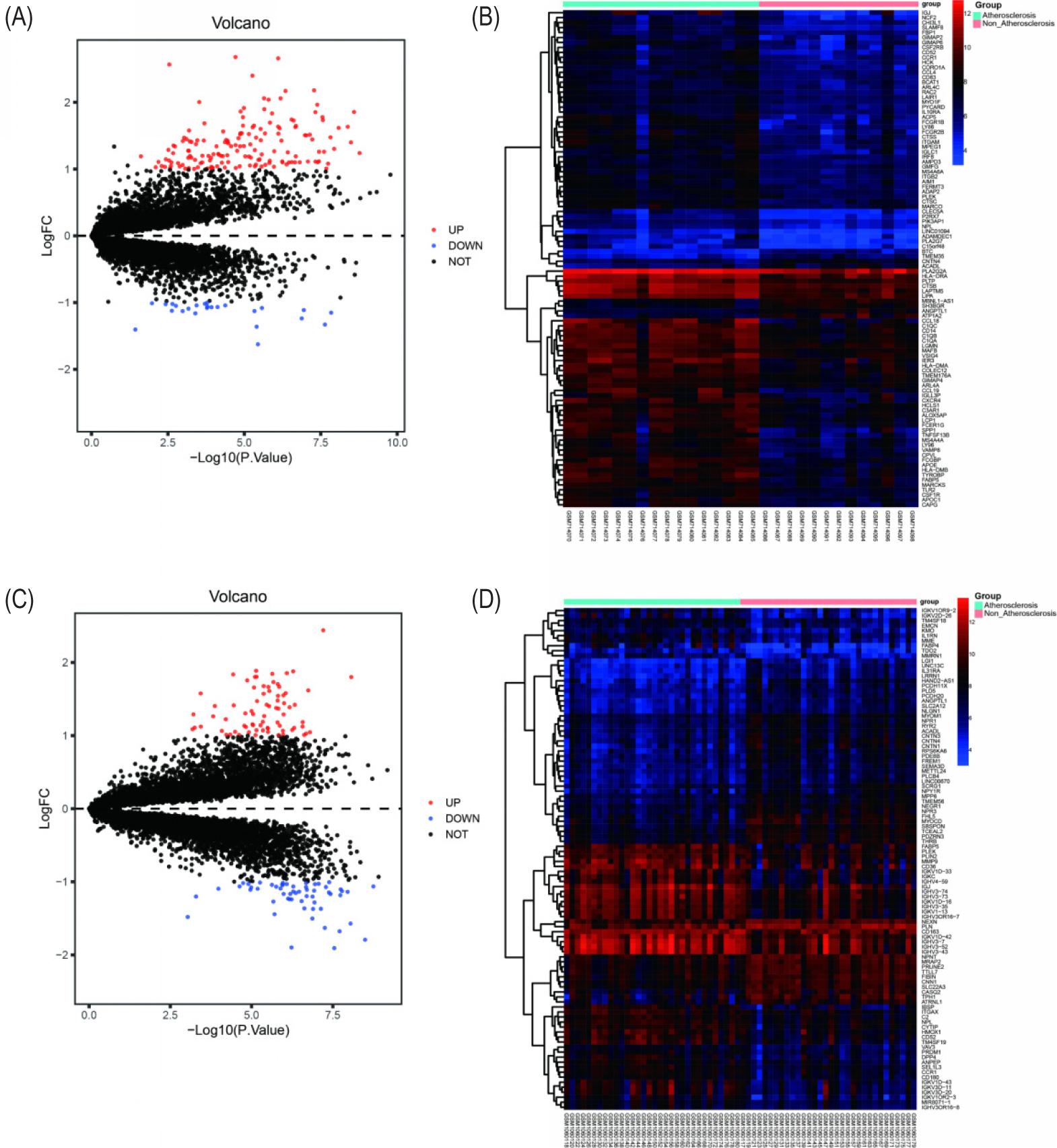
Atherosclerosis-related database was searched by DisGeNet and GENECARDs databases with the keyword “atherosclerosis” to obtain the therapeutic targets of AS. GSE28829 and GSE43292 data sets were obtained. In the GSE28829 dataset, 157 genes were up-regulated and 25 genes were down-regulated. In the GSE43292 dataset, 75 genes were up-regulated and 57 genes were down-regulated (Figure 2).
Figure 2. Therapeutic targets of AS. (A and B) Volcano plot and heat map of GSE28829 dataset. (C and D) Volcano plot and heat map of GSE43292 dataset.
Potential therapeutic targets
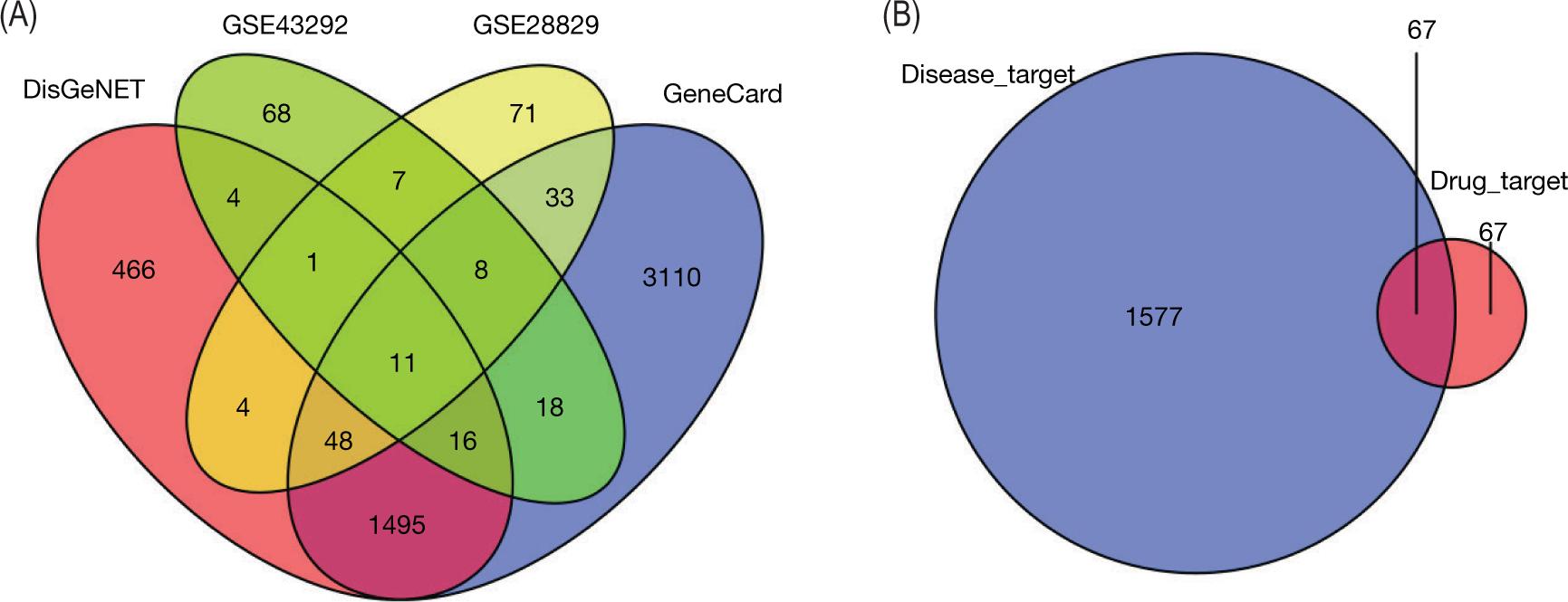
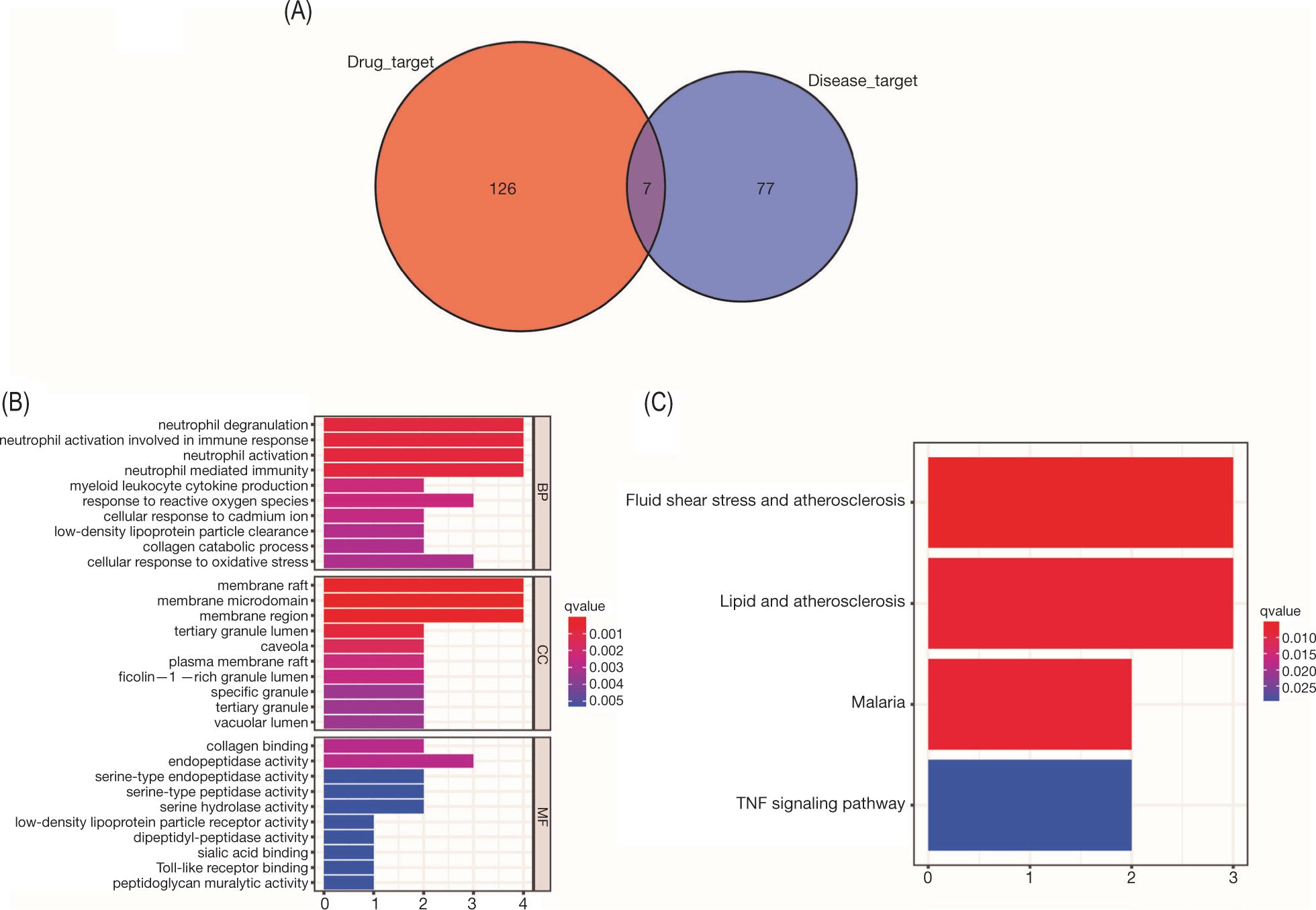
While DEGs were screened from the GEO dataset, the AS-related gene intersections from the DisGeNet and GENECARDs databases were screened for targets that intersected at least twice or thrice and defined as AS-related therapeutic targets. Finally, we screened 67 potential targets of SL for treating AS (Figure 3).
Figure 3. Targets associated with AS treatment. (A) Venn diagrams of AS treatment in two disease-databases and GEO dataset. (B) Venn diagrams of targets in at least two databases in (A) and SL treatment targets.
GO, KEGG, and PPI network analysis
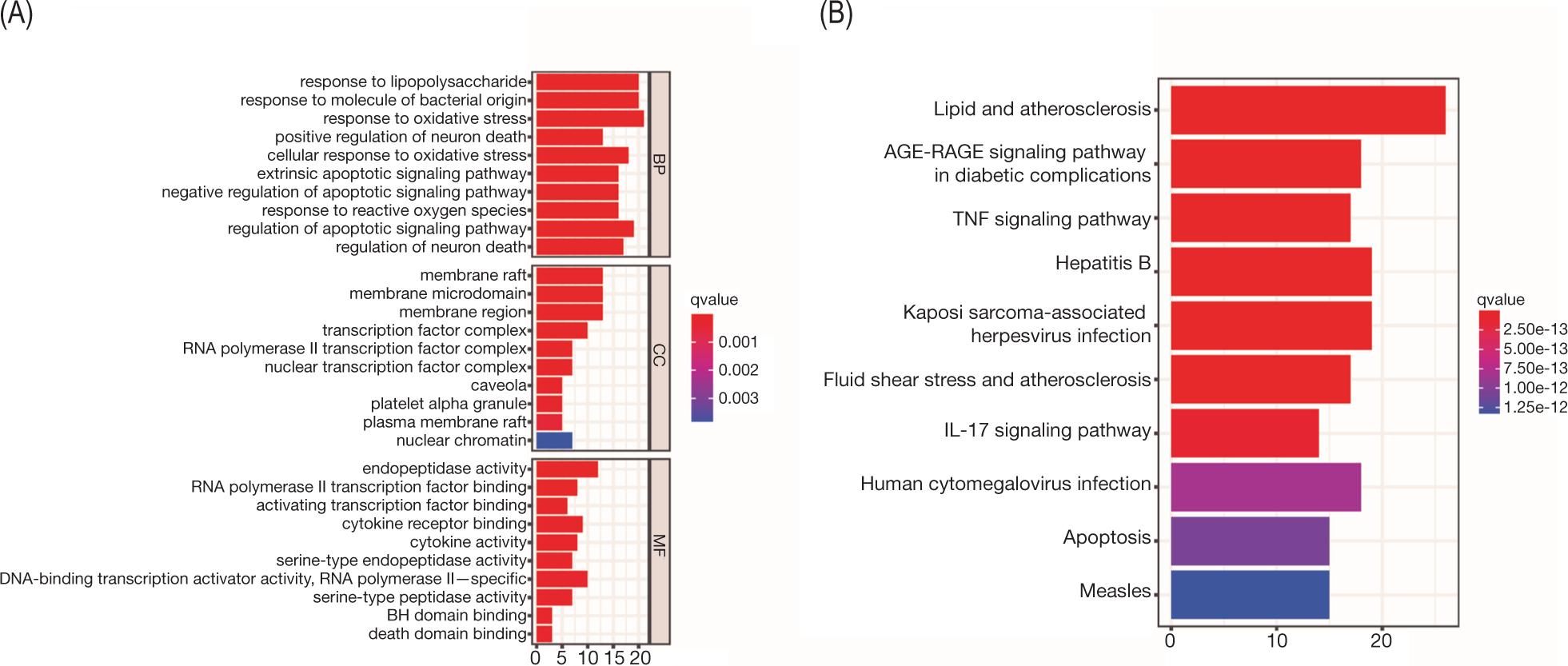
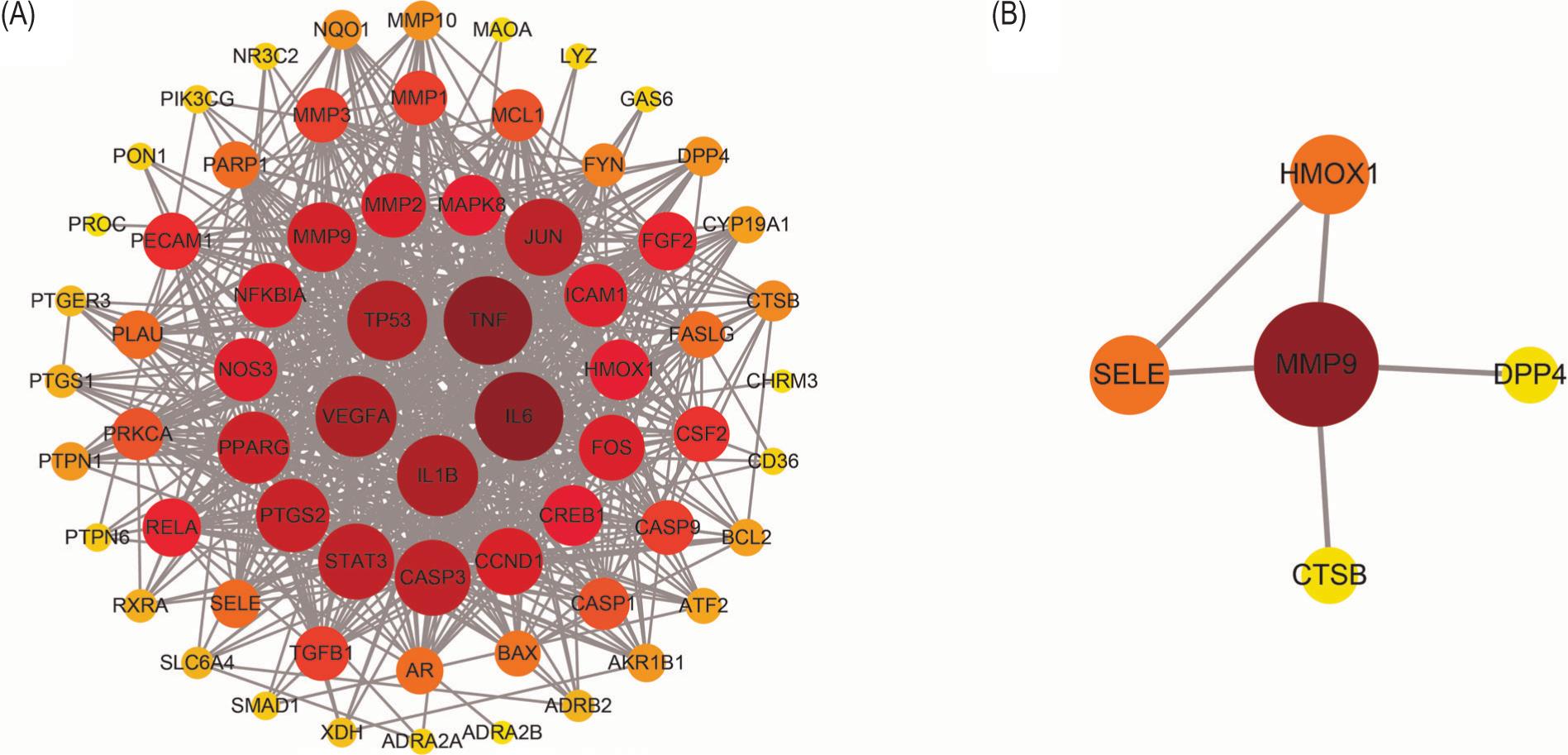
In order to explore further the mechanism of treating AS, we performed enrichment analysis and PPI network construction for the above-screened targets. First, we identified genes that intersected in the two databases. In terms of BP, the targets were mainly enriched in response to lipopolysaccharide and response to oxidative stress, mainly focused on membrane raft and membrane microdomain. On MF, mainly endopeptidase activity, RNA polymerase II transcription factor binding and KEGG enrichment analysis showed that the targets were mainly enriched in lipid and AS, AGE-RAGE signaling pathway in diabetic complications, tumor necrosis factor (TNF) signaling pathway in diabetic complications, TNF signaling pathway and interleukin-17 (IL-17) signaling pathway (Figure 4). While seven intersecting genes were present in three databases; enrichment analysis showed that BP mainly involved neutrophil degranulation, and neutrophil activation involved in immune response; in CC, membrane raft and membrane microdomain were involved; and MF was mainly related to collagen binding and endopeptidase activity. The pathway enrichment showed fluid shear stress and AS, lipid and AS, and TNF signaling pathway (Figure 5). Subsequent construction of PPI networks for potential therapeutic targets showed that VEGFA, IL-1B, IL-6, TP53, and TNF were the core genes with intersecting genes in two databases, while MMP9, SELE, HMOX1, DPP4, and CTSB were core target genes in three datasets (Figure 6).
Figure 4. Gene ontology (GO) and KEGG enrichment analysis of SL treatment targets in at least two databases. (A) GO enrichment analysis [top 10 results for biological process (BP), cellular component (CC), and molecular function (MF) enrichment analysis]. (B) KEGG enrichment analysis of treatment targets (top 10 results).
Figure 5. Enrichment analysis of three databases shows SL therapeutic targets. (A) Venn diagram of three databases and disease targets. (B) GO enrichment analysis [top 10 results for biological process (BP), cellular component (CC), and molecular function (MF) enrichment analysis]. (C) KEGG enrichment analysis of therapeutic targets (top 10 results).
Figure 6. Protein–protein interaction (PPI) network of SL treatment AS. (A) PPI network analysis of at least two database intersection targets. (B) PPI network analysis of three database intersection targets.
Molecular docking
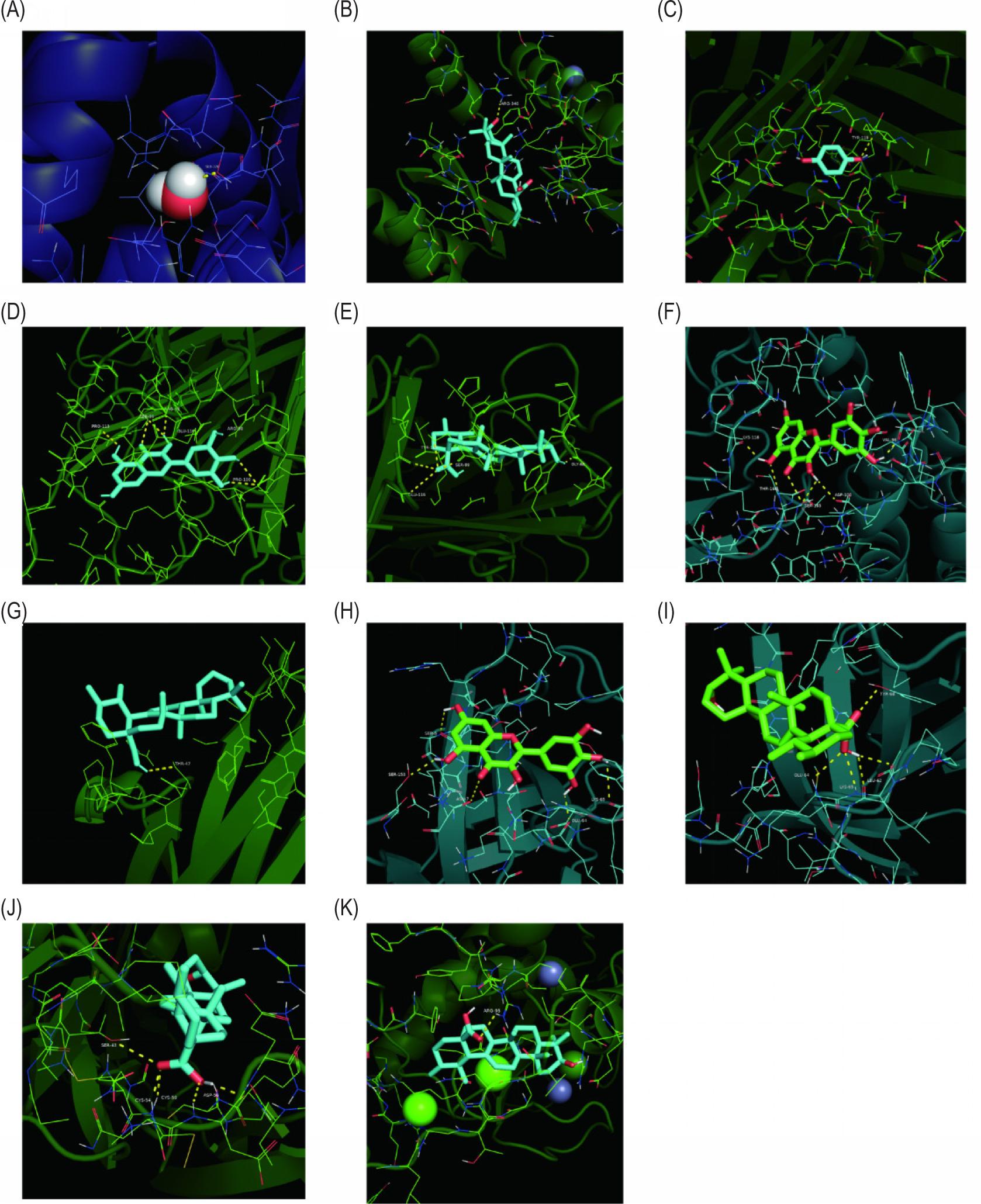
Results of PPI network analysis showed that six targets with the highest degree of TNF, TP53, IL-6, IL-1B, VEGFA, and MMP9 were identified, and the corresponding active ingredients were identified by correlated molecular docking (Figure 7).
Figure 7. Results of the docking of core targets of PPI network with their corresponding active ingredient molecules. (A and B) p53 with 3,4,5-trihydroxybenzoic acid (gallic acid) and ursolic acid docking results. (C–E) Tumor necrosis factor (TNF) with hydroquinone, myricetin, and ursolic acid docking results. (F and G) Docking results of IL-6 with myricetin and ursolic acid. (H and I) Docking results of IL-1B with myricetin and ursolic acid. (J) Docking results of VEGFA with ursolic acid. (K) Docking results of MMP9 with ursolic acid.
Identification of core active ingredients and their mechanisms of action
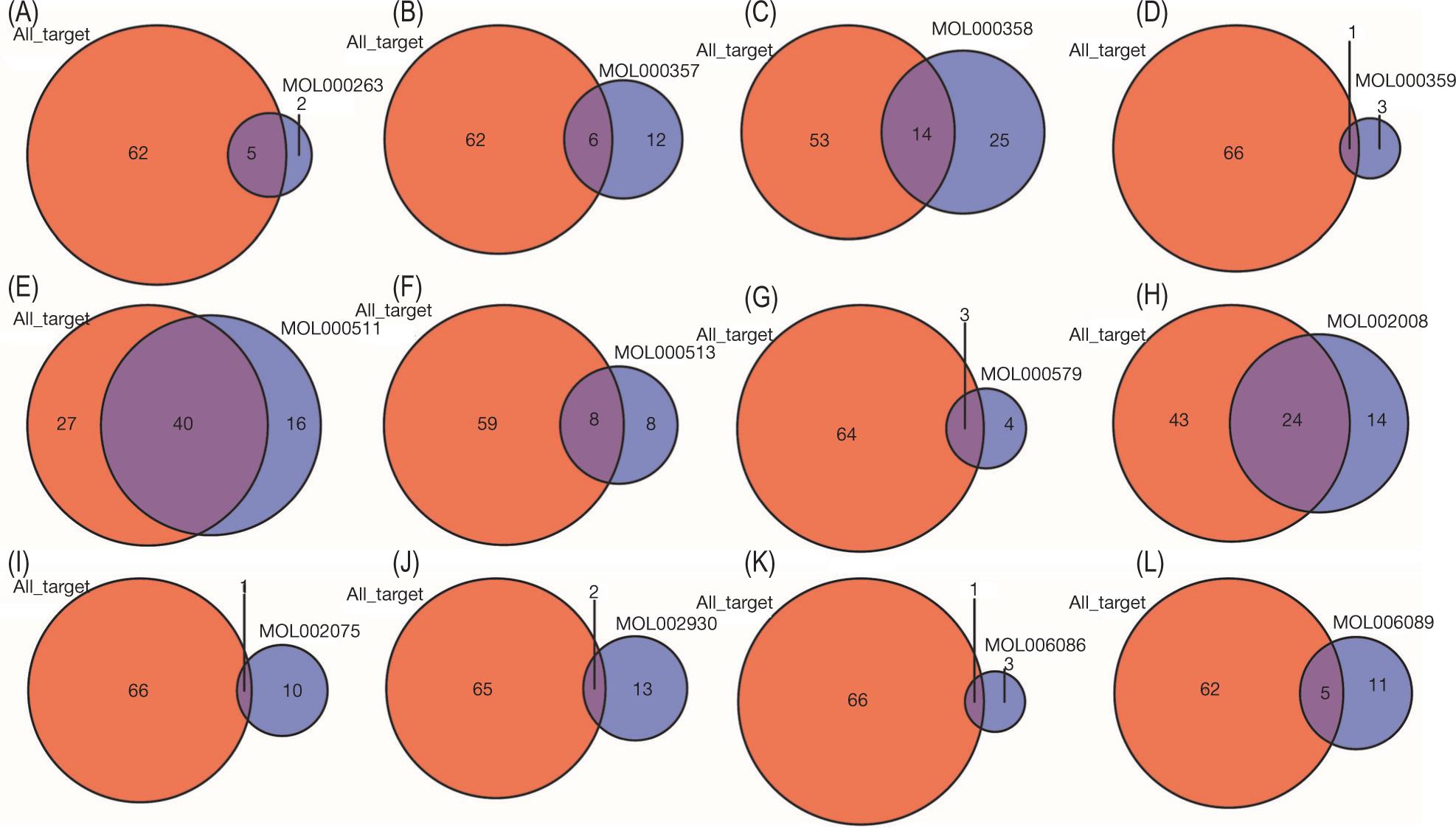
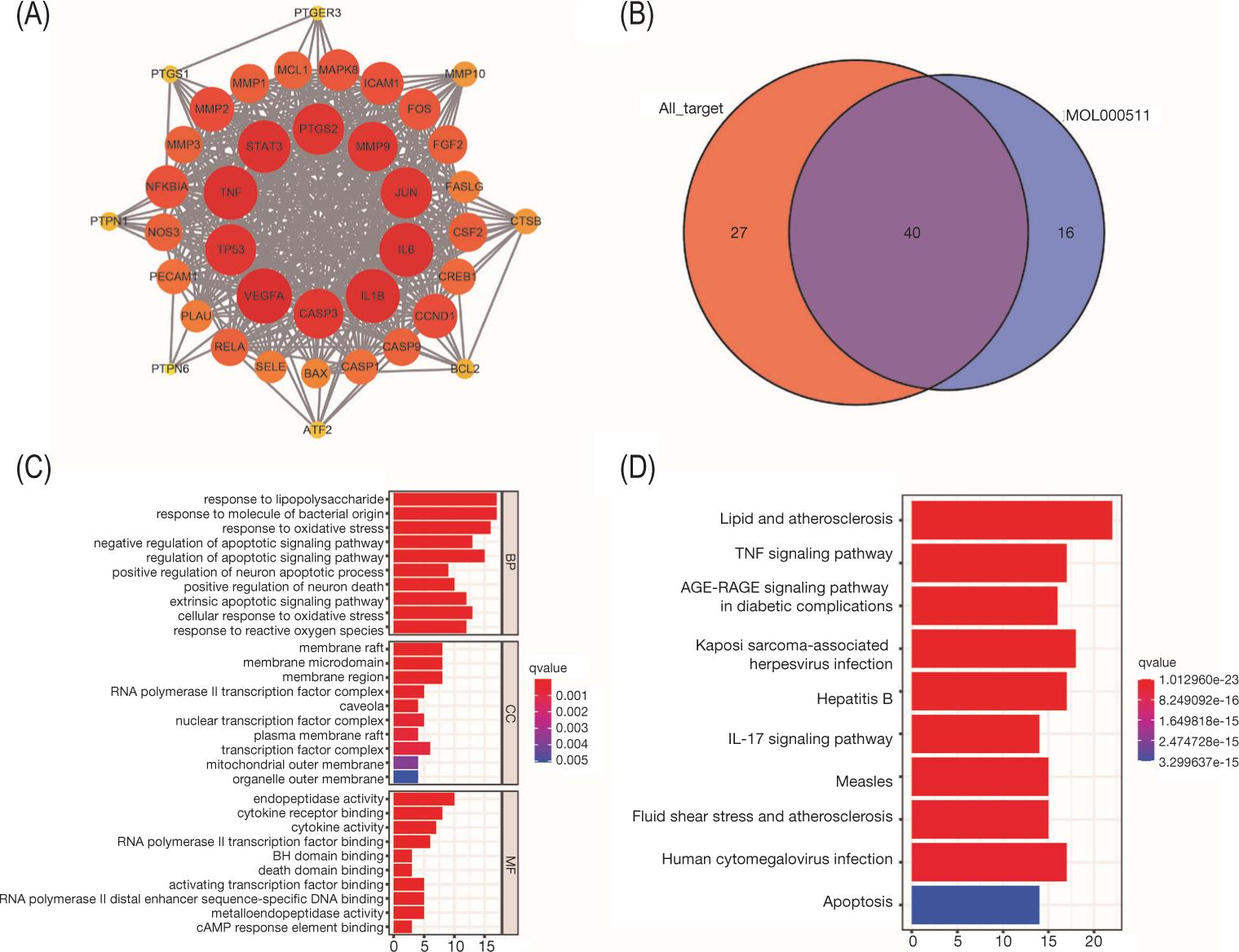
We intersected the identified active ingredients of SL with the disease targets of AS and found that they have at least one and a maximum of 40 target genes (Figure 8). Finally, we identified ursolic acid as the main core ingredient of SL, and 40 targets of ursolic acid overlapped with the potential therapeutic targets of SL for AS. Further, PPI network and enrichment analyses showed that ursolic acid had a high overlapping with the core target genes of SL, GO, and KEGG (Figure 9, Table S1).
Figure 8. Intersection of active ingredients of SL with AS therapeutic targets. (A) MOL000263. (B) MOL000357. (C) MOL000358. (D) MOL000359. (E) MOL000511. (F) MOL000513. (G) MOL000579. (H) MOL002008. (I) MOL002075. (J) MOL002930. (K) MOL006086. (L) MOL006089.
Figure 9. Bioinformatics analysis of potential therapeutic targets of ursolic acid. (A) Protein–protein interaction (PPI) network of therapeutic targets. (B) Venn diagram of potential therapeutic targets of ursolic acid and therapeutic targets of Kuntai -Capsule (KTC). (C) Gene ontology (GO) enrichment analysis of therapeutic targets (top 10 results for biological process [BP], cellular component [CC], and molecular function [MF] enrichment analysis). (D) Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of therapeutic targets (top 10 results).
Discussion
The pathogenesis of AS is more diverse, with lipid deposition in the circulation. The generation of reactive oxygen species (ROS) causes damage to vascular endothelial function, inducing monocyte migration foam, inflammatory mediator release, and intimal fibrous proliferation, leading to atheromatous plaques (Hafiane, 2019). The complex and variable nature of the AS process poses a great challenge for treatment, and although statins are the main measure for treating AS, because of their single target of action, long-term use can bring serious adverse effects (Varatharajalu et al., 2016). Recent studies have shown that SL has a preventive and curative effect on cardiovascular disease and has the advantage of low toxic adverse reactions. In our study, we proposed to construct a component–target–disease pathway network with the help of network pharmacology, and used PPI to construct network targets for SL and AS from a holistic and systemic perspective, further screening the core genes, identifying the active ingredients through molecular docking, and subsequently exploring their potential mechanisms of action through GO and KEGG enrichment analysis.
The targets of disease and SL were crossed to finally obtain 67 potential therapeutic targets of SL for AS, and five core targets with intersection in three databases; MMP9, SELE, HMOX1, DPP4, and CTSB were further screened. MMP9 plays an important role in the metabolism of extracellular matrix and collagen, and promotes smooth muscle migration to the subendothelium to form atheromatous plaques, thereby accelerating the progression of atherosclerotic plaque formation. It is also associated with plaque instability and rupture (Bakhshian et al., 2017; Rossano et al., 2014; Sierra et al., 2018). SELE encodes the protein present in cytokine-stimulated endothelial cells, and the current study showed that SELE had a key role in the development of AS, and targeting SELE would reduce atherosclerotic lesions, adverse cardiac remodeling, and dysfunction (Fereydouni et al., 2020; Tsoref et al., 2018).
Studies have shown that HMOX1 is highly expressed in plaques of AS patients and positively correlates with disease, accompanied by the production of MMPs and macrophage infiltration (Kishimoto et al., 2018; Wu et al., 2022). DPP4 has pleiotropic properties, and DPP4 inhibitors exert anti-AS effects by a variety of mechanisms that inhibit inflammation and oxidative stress, and improve endothelial cell dysfunction, in addition to improving glycemia, which helps to reduce cardiovascular risk in patients with comorbid diabetes (Liu et al., 2020; Love and Liu, 2021). Studies have shown that CTSB is upregulated in AS lesions; it degrades extracellular matrix; increases the fragility of atherosclerotic plaques; and has a crucial role in the pathogenesis of AS (Zhao and Herrington, 2016).
The KEGG results suggest that SL can act simultaneously on multiple signaling pathways associated with AS disease, including lipid, TNF signaling pathway, IL-17 signaling pathway, and AGE-RAGE signaling pathway in diabetic complications. The AGE-RAGE axis in diabetic patients promotes AS and decreases glycation end--products. RAGE expression inhibits the progression of AS; this progression may be associated with vascular protection.
The pathogenesis of AS begins with vascular endothelial cell dysfunction accompanied by low-density lipoprotein (LDL) retention, modified LDL promoting endothelial cell activation and vesicular cell formation, and massive accumulation of intracellular and extracellular matrix lipids as the initial manifestation of AS (Allahverdian et al., 2014; Mundi et al., 2018; Stary et al., 1994).
The most common diabetic complication is cardiovascular and cerebrovascular diseases. Many studies have proved that TNF signaling pathway had a critical role in various diabetic complications. TNF apoptosis, as well as inflammation, and TNF cells are major mediators of inflammation and immunity, and are associated with the pathogenesis of several human diseases. TNF mainly includes TNF-β and TNF-α, which are secreted by T lymphocytes and macrophages, respectively. TNF-α can increase endothelial inflammation and AS by mediating the NF-κB pathway and is involved in the development of AS (Chen and Goeddel, 2002; Gao et al., 2016).
IL-17 is mainly produced by Th17 cells and coordinates local tissue inflammation by inducing the release of pro-inflammatory cytokines and mobilization of chemokines by neutrophils from various cell types, including epithelial cells (Veldhoen et al., 2006). IL-17 also has a significant inflammation pathway. The role of IL-17 has been extensively studied in a variety of inflammatory and autoimmune diseases, but its role in the development of AS remains debatable. Induction of IL-17R-deficient mice on a Western-type diet revealed a 46% reduction in aortic roots and plaques. whereas in vivo use of IL-17A-blocking antibodies slowed the development of AS (Kumbhani et al., 2015; Liu et al., 2014). In contrast to these studies, increased production of IL-10I and L-17A induced an anti-inflammatory macrophage phenotype, slowed down lesion progression, and reduced vascular inflammation while acting as a protective cytokine for AS (Schraml et al., 2009). From these we conclude that these signaling pathways play a critical role in the progression of AS. However, well-defined functioning of these signaling pathways in AS needs further research.
The main active ingredients of SL include myricetin, oleanolic acid, ursolic acid, sitosterol, and beta-sitosterol. Myricetin is a natural flavonoid extracted from berries, grapes, and herbs with antioxidant properties. Current studies have shown that it can inhibit cholesterol accumulation in foam cells, thereby improving AS, in addition to inhibiting vascular smooth muscle cell proliferation and migration and suppressing of neoplastic endothelial proliferation (Chen et al., 2021; Meng et al., 2019).
Oleanolic acid is a pentacyclic triterpenoid widely found in plants, and recent studies have shown that it mainly exerts antioxidant effects but could be an antitumor as well (Li et al., 2015; Pollier and Goossens, 2012; Wang et al., 2013). Recent researches have shown that oleanolic acid could prevent the development of AS and protect human umbilical vein endothelial cells (Pan et al., 2018; Zhang et al., 2018).
Ursolic acid is a pentacyclic triterpene carboxylic acid that mainly exerts anti-inflammatory, antioxidant, antitumor, and anti-hyperlipidemic effects (Ali et al., 2007; Ikeda et al., 2008; Somova et al., 2003). In a subsequent study, ursolic acid was found to attenuate AS formation through ROS/NF-κB pathway-mediated lectin-like oxidized low-density lipoprotein receptor 1 (LOX-1) (Li et al., 2018). Both sitosterol and beta-sitosterol are to phytosterols, and studies have shown that sitosterol significantly reduces the expression of NLRP3 gene, a key protein of macrophage inflammatory vesicles, and inhibits the activation of apoptotic proteases, thus acting as an anti-inflammatory agent Beta-sitosterol inhibits the secretion of inflammatory elements by macrophages and suppresses the migration of vascular smooth muscle cells (He et al., 2022; Liao et al., 2018). The subsequent functional and pathway enrichment analysis of these active components further validated the above mechanism of screening target genes with consistency. Taken together, these studies suggest that the main active ingredients of SL exert anti-atherosclerotic effects through antioxidant, anti-inflammatory, and -anti-lipidemic effects, acting on different targets through multiple signaling pathways, thus preventing and treating cardiovascular diseases. However, the limitations of this study including shorting the validation experiments. As well, some imformation such as the patients treatment regimens is lacking.
Conclusion
In summary, this study found that SL can exert anti--atherosclerotic effects through 12 major components, including the regulation of TNF and IL-17 signaling pathway, thus providing a theoretical basis for the follow-up study of SL against AS.
Author Contributions
Conception and design: BJ Zhu, GY Nai, WJ Zhou, and ZF Ma; Administrative support: TX Pan; Provision of study materials/patients: GM Ling and ZD Huang; Collection and assembly of data: ZZ Shi, JX Lin; Data analysis and interpretation: BJ Zhu, GY Nai, and ZF Ma. All authors were involved in the writing of this paper and approving its final draft.
Funding
This research was supported Baise Scientific Research and Technology Development Program “City-School Science and Technology Cooperation” Project (Baise Science and Technology 20170813).
REFERENCES
Ali, M., Ibrahim, S., Jalil, S., et al., 2007. Ursolic acid: a potent inhibitor of superoxides produced in the cellular system. Phytotherapy Research: PTR 21: 558–561. 10.1002/ptr.2108
Allahverdian, S., Chehroudi, A., McManus, B., et al., 2014. Contribution of intimal smooth muscle cells to cholesterol accumulation and macrophage-like cells in human atherosclerosis. Circulation 129: 1551–1559. 10.1161/CIRCULATIONAHA.113.005015
Bakhshian, Nik A., Hutcheson, J., Aikawa, E., 2017. Extracellular vesicles as mediators of cardiovascular calcification. Frontiers in Cardiovascular Medicine 4: 78. 10.3389/fcvm.2017.00078
Chen, G. and Goeddel, D., 2002. TNF-R1 signaling: a beautiful pathway. Science (New York, NY) 296: 1634–1635. 10.1126/science.1071924
Chen, G., Xu, H., Wu, Y., et al., 2021. Myricetin suppresses the proliferation and migration of vascular smooth muscle cells and inhibits neointimal hyperplasia via suppressing TGFBR1 signaling pathways. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology 92: 153719. 10.1016/j.phymed.2021.153719
Fereydouni, Z., Amirinezhad, Fard E., Mansouri, K., et al., 2020. Saponins from Tribulus terrestris L. extract down-regulate the expression of ICAM-1, VCAM-1 and E-selectin in human endothelial cell lines. International Journal of Molecular and Cellular Medicine 9: 73–83.
Gao, W., Liu, H., Yuan, J., et al., 2016. Exosomes derived from mature dendritic cells increase endothelial inflammation and atherosclerosis via membrane TNF-α mediated NF-κB pathway. Journal of Cellular and Molecular Medicine 20: 2318–2327. 10.1111/jcmm.12923
Hafiane, A., 2019. Vulnerable plaque, characteristics, detection, and potential therapies. Journal of Cardiovascular Development and Disease 6. 10.3390/jcdd6030026
He, S., He, S., Chen, Y., et al., 2022. Beta-sitosterol modulates the migration of vascular smooth muscle cells via the PPARG/AMPK/mTOR pathway. Pharmacology 1–15. 10.1159/000525218
Humphries, S., Cooper, J., Seed, M., et al., 2018. Coronary heart disease mortality in treated familial hypercholesterolaemia: update of the UK Simon Broome FH register. Atherosclerosis 274: 41–46. 10.1016/j.atherosclerosis.2018.04.040
Ikeda, Y., Murakami, A. and Ohigashi, H., 2008. Ursolic acid: an anti-and pro-inflammatory triterpenoid. Molecular Nutrition & Food Research 52: 26–42. 10.1002/mnfr.200700389
Keeter, W., Ma, S., Stahr, N., et al., 2022. Atherosclerosis and multi-organ-associated pathologies. Seminars in Immunopathology 44: 363–374. 10.1007/s00281-022-00914-y
Kishimoto, Y., Sasaki, K., Saita, E., et al., 2018. Plasma heme oxygenase-1 levels and carotid atherosclerosis. Stroke 49: 2230–2232. 10.1161/STROKEAHA.118.022256
Kobiyama, K. and Ley, K., 2018. Atherosclerosis. Circulation Research 123: 1118–1120. 10.1161/CIRCRESAHA.118.313816
Kumbhani, D., Marso, S., Alvarez, C., et al., 2015. State-of-the-art: hypo-responsiveness to oral antiplatelet therapy in patients with type 2 diabetes mellitus. Current Cardiovascular Risk Reports 9: 4. 10.1007/s12170-014-0430-5
Lee, Y., Cho, Y., Kim, E., et al., 2019. Reduced expression of pyruvate kinase in kidney proximal tubule cells is a potential mechanism of pravastatin altered glucose metabolism. Scientific Reports 9: 5318. 10.1038/s41598-019-39461-2
Li, M., Qi, Z., Hao, Y., et al., 2017. New adducts of iriflophene and flavonoids isolated from Sedum aizoon L. with potential antitumor activity. Molecules (Basel, Switzerland) 22. 10.3390/molecules22111859
Li, L., Wei, L., Shen, A., et al., 2015. Oleanolic acid modulates multiple intracellular targets to inhibit colorectal cancer growth. International Journal of Oncology 47: 2247–2254. 10.3892/ijo.2015.3198
Li, Q., Zhao, W., Zeng, X., et al., 2018. Ursolic acid attenuates atherosclerosis in ApoE mice: role of LOX-1 mediated by ROS/NF-κB pathway. Molecules (Basel, Switzerland) 23. 10.3390/molecules23051101
Liao, P., Lai, M., Hsu, K., et al., 2018. Identification of β-sitosterol as in vitro anti-inflammatory constituent in Moringa oleifera. Journal of Agricultural and Food Chemistry 66: 10748–10759. 10.1021/acs.jafc.8b04555
Liu, H., Guo, L., Xing, J., et al., 2020. The protective role of DPP4 inhibitors in atherosclerosis. European Journal of Pharmacology 875: 173037. 10.1016/j.ejphar.2020.173037
Liu, M., Li, X., Lu, L., et al., 2014. Cardiovascular disease and its relationship with chronic kidney disease. European Review for Medical and Pharmacological Sciences 18: 2918–2926.
Liu, S., Li, Y., Zeng, X., et al., 2019. Burden of cardiovascular diseases in China, 1990–2016: Findings from the 2016 Global Burden of Disease Study. JAMA Cardiology 4: 342–352. 10.1001/jamacardio.2019.0295
Love, K. and Liu, Z., 2021. DPP4 activity, hyperinsulinemia, and atherosclerosis. Journal of Clinical Endocrinology and Metabolism 106: 1553–1565. 10.1210/clinem/dgab078
Meng, Z., Wang, M., Xing, J., et al., 2019. Myricetin ameliorates atherosclerosis in the low-density-lipoprotein receptor knockout mice by suppression of cholesterol accumulation in macrophage foam cells. Nutrition & Metabolism 16: 25. 10.1186/s12986-019-0354-7
Mundi, S., Massaro, M., Scoditti, E., et al., 2018, Endothelial permeability, LDL deposition, and cardiovascular risk factors-a review. Cardiovascular Research 114: 35–52. 10.1093/cvr/cvx226
Ott, B., Daiello, L., Dahabreh, I., et al., 2015. Do statins impair cognition? A systematic review and meta-analysis of randomized controlled trials. Journal of General Internal Medicine 30: 348–58. 10.1007/s11606-014-3115-3
Pan, Y., Zhou, F., Song, Z., et al., 2018. Oleanolic acid protects against pathogenesis of atherosclerosis, possibly via FXR-mediated angiotensin (Ang)-(1–7) upregulation. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie 97: 1694–1700. 10.1016/j.biopha.2017.11.151
Panes, O., González, C., Hidalgo, P., et al., 2017. Platelet tissue factor activity and membrane cholesterol are increased in hypercholesterolemia and normalized by rosuvastatin, but not by atorvastatin. Atherosclerosis 257: 164–71. 10.1016/j.atherosclerosis.2016.12.019
Pollier, J. and Goossens, A., 2012. Oleanolic acid. Phytochemistry 77: 10–15. 10.1016/j.phytochem.2011.12.022
Rossano, R., Larocca, M., Riviello, L., et al., 2014. Heterogeneity of serum gelatinases MMP-2 and MMP-9 isoforms and charge variants. Journal of Cellular and Molecular Medicine 18: 242–252. 10.1111/jcmm.12181
Schraml, B., Hildner, K., Ise, W., et al., 2009. The AP-1 transcription factor Batf controls T(H)17 differentiation. Nature 460: 405–409. 10.1038/nature08114
Shi, S., Ji, X., Shi, J., et al., 2022. Andrographolide in atherosclerosis: integrating network pharmacology and in vitro pharmacological evaluation. Bioscience Reports 42. 10.1042/BSR20212812
Sierra, S., Luquin, N. and Navarro-Otano, J., 2018. The endocannabinoid system in cardiovascular function: novel insights and clinical implications. Clinical Autonomic Research: Official Journal of the Clinical Autonomic Research Society 28: 35–52. 10.1007/s10286-017-0488-5
Somova, L., Nadar, A., Rammanan, P., et al., 2003. Cardiovascular, antihyperlipidemic and antioxidant effects of oleanolic and ursolic acids in experimental hypertension. Phytomedicine: International Journal of Phytotherapy and Phytopharmacology 10: 115–121. 10.1078/094471103321659807
Song, L., Zhang, J., Lai, R., et al., 2021. Chinese herbal medicines and active metabolites: potential antioxidant treatments for atherosclerosis. Frontiers in Pharmacology 12: 675999. 10.3389/fphar.2021.675999
Stary, H., Chandler, A., Glagov, S., et al., 1994. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 89: 2462–2478. 10.1161/01.CIR.89.5.2462
Sun, J., Yin, X., Liu, H., et al., 2018. Rapamycin inhibits ox-LDL--induced inflammation in human endothelial cells in vitro by inhibiting the mTORC2/PKC/c-Fos pathway. Acta Pharmacologica Sinica 39: 336–344. 10.1038/aps.2017.102
Troidl, K., Schubert, C., Vlacil, A., et al., 2020. The lipopeptide MALP-2 promotes collateral growth. Cells 9. 10.3390/cells9040997
Tsoref, O., Tyomkin, D., Amit, U., et al., 2018. E-selectin-targeted copolymer reduces atherosclerotic lesions, adverse cardiac remodeling, and dysfunction. Journal of Controlled Release: Official Journal of the Controlled Release Society 288: 136–147. 10.1016/j.jconrel.2018.08.029
Varatharajalu, R., Garige, M., Leckey, L., et al., 2016. Protective role of dietary curcumin in the prevention of the oxidative stress induced by chronic alcohol with respect to hepatic injury and antiatherogenic markers. Oxidative Medicine and Cellular Longevity 2016: 5017460. 10.1155/2016/5017460
Veldhoen, M., Hocking, R., Atkins, C., et al., 2006. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24: 179–189. 10.1016/j.immuni.2006.01.001
Wang, X., Liu, R., Zhang, W., et al., 2013. Oleanolic acid improves hepatic insulin resistance via antioxidant, hypolipidemic and anti-inflammatory effects. Molecular and Cellular Endocrinology 376: 70–80. 10.1016/j.mce.2013.06.014
Wang, H., Xu, F., Zhang, X., et al., 2021. Transcriptomic analysis reveals antibacterial mechanism of flavonoids from Sedum aizoon L. against Pseudomonas fragi. 10.1016/j.foodcont.2021.108755
Wu, D., Hu, Q., Wang, Y., et al. 2022. Identification of HMOX1 as a critical ferroptosis-related gene in atherosclerosis. Frontiers in Cardiovascular Medicine 9: 833642. 10.3389/fcvm.2022.833642
Wu, Z., Li, W., Liu, G., et al., 2018. Network-based methods for prediction of drug-target interactions. Frontiers in Pharmacology 9: 1134. 10.3389/fphar.2018.01134
Wu, M., Xu, K., Guo, Y., et al., 2019. Lipoprotein(a) and-atherosclerotic cardiovascular disease: current understanding and future perspectives. Cardiovascular Drugs and Therapy 33: 739–48. 10.1007/s10557-019-06906-9
Xi, J., Rong, Y., Zhao, Z., et al., 2021. Scutellarin ameliorates high-glucose-induced vascular endothelial cells injury by activating PINK1/Parkin-mediated mitophagy. Journal of Ethnophar-macology 271: 113855. 10.1016/j.jep.2021.113855
Xie, X., Ma, X., Zeng, S., et al., 2020. Mechanisms of berberine for the treatment of atherosclerosis based on network pharmacology. Evidence-Based Complementary and Alternative Medicine: eCAM 2020: 3568756. 10.1155/2020/3568756
Xu, T., Wang, Z., Lei, T., et al., 2015. New flavonoid glycosides from Sedum aizoon L. Fitoterapia 101: 125–132. 10.1016/j.fitote.2014.12.014
Zhang, W., Feng, J., Cheng, B., et al., 2018. Oleanolic acid protects against oxidative stress-induced human umbilical vein endothelial cell injury by activating AKT/eNOS signaling. Molecular Medicine Reports 18: 3641–3648. 10.3892/mmr.2018.9354
Zhao, C. and Herrington, D., 2016. The function of cathepsins B, D, and X in atherosclerosis. American Journal of Cardiovascular Disease 6: 163–170.
Zhou, S., Ai, Z., Li, W., et al., 2020. In vitro deciphering the pharmacological mechanisms of taohe-chengqi decoction extract against renal fibrosis through integrating network pharmacology and experimental validation. Frontiers in Pharmacology 11: 425. 10.3389/fphar.2020.00425
Supplementary
Table S1. Overlapping genes of drug targets and disease targets.
| Genes | |
|---|---|
| AR | BCL2 |
| PPARG | PRKCA |
| PTGS2 | TGFB1 |
| PIK3CG | PON1 |
| DPP4 | NR3C2 |
| PTGS1 | PLAU |
| AKR1B1 | CTSB |
| SMAD1 | VEGFA |
| FYN | CCND1 |
| RELA | FOS |
| STAT3 | MMP9 |
| BAX | TP53 |
| CASP9 | MAPK8 |
| MMP2 | NFKBIA |
| TNF | MMP1 |
| JUN | MMP3 |
| IL6 | FGF2 |
| CASP3 | MMP10 |
| XDH | CREB1 |
| IL1B | SELE |
| CD36 | PTGER3 |
| CYP19A1 | MCL1 |
| PARP1 | ATF2 |
| PROC | CSF2 |
| CHRM3 | PECAM1 |
| HMOX1 | PTPN6 |
| ICAM1 | NOS3 |
| NQO1 | PTPN1 |
| MAOA | FASLG |
| LYZ | CASP1 |
| GAS6 | ENPP7 |
| RXRA | ADRA2A |
| ADRB2 | ADRA2B |
| SLC6A4 | |