
Download
Dressing percentage % = carcass weight live weight × 100 % . 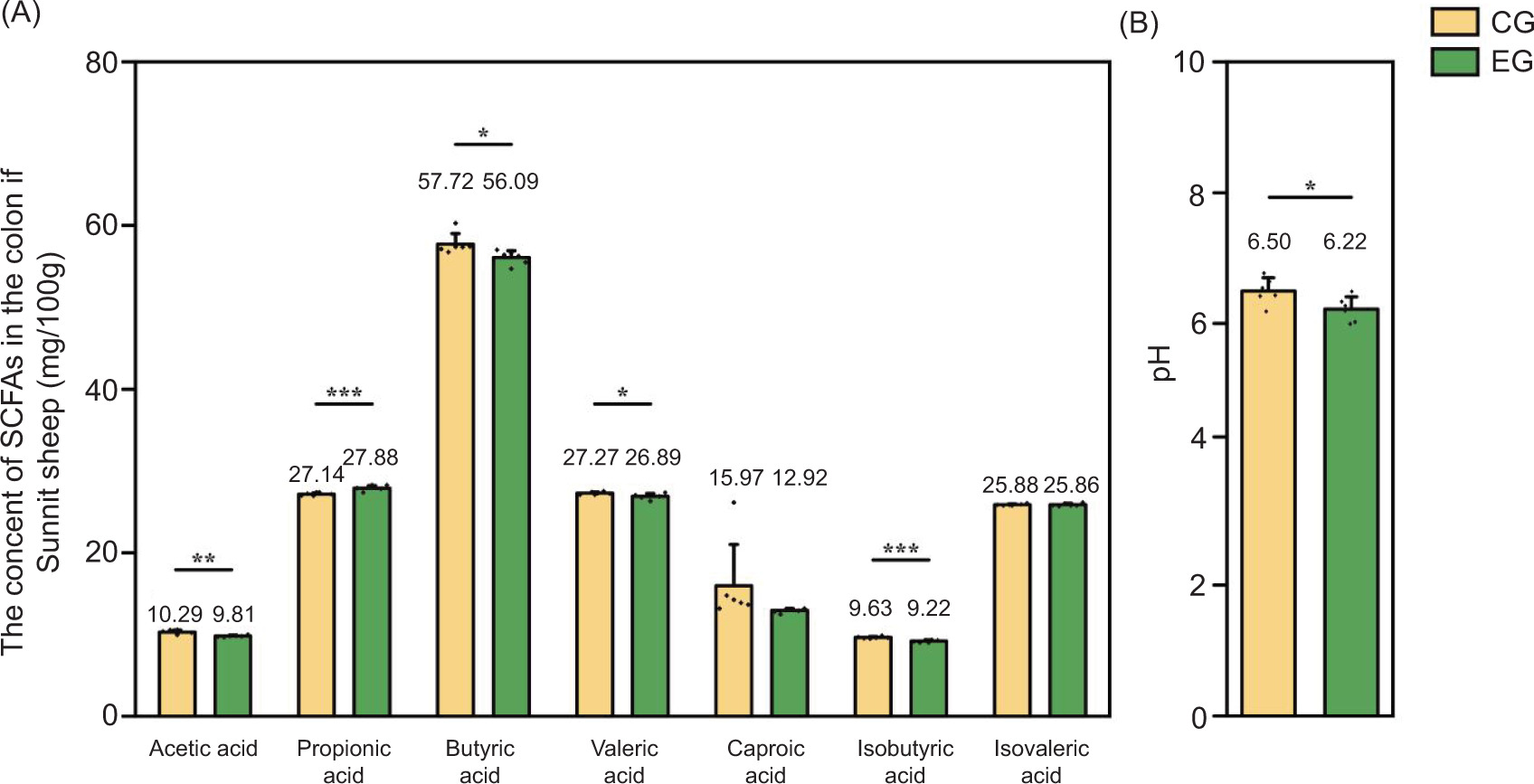
Research Article
Exercise influences fatty acids in the longissimus dorsi muscle of Sunit lambs and improves dressing percentage by affecting digestion, absorption, and lipid metabolism
Ting Liu1,2, Taiwu Zhang1,2, Yanni Zhang3, Min Zhang1,2, Maoqin Zhai1,2, Weihao Wang1,2, Chenlei Wang1,2, Yan Duan1,2*, Ye Jin1,2*
1College of Food Science and Engineering, Inner Mongolia Agricultural University, Hohhot, China;
2Integrative Research Base of Beef and Lamb Processing Technology, Ministry of Agriculture and Rural Affairs of the People’s Republic of China, Hohhot;
3Beijing Youyuan Animal Husbandry Co. Ltd, Shuguang Xili Jia, Chaoyang District, Beijing, PR China
Abstract
The transition from grazing to captive rearing resulted in a significant decline in meat quality. To enhance the quality of meat from captive sheep, we examined the impact of exercise on Sunit lambs. The lambs, with similar body weight and aged 3 months underwent a 90-day period of thought-driven exercise. At the end of the exercise period, we collected colonic contents, epithelial tissues, and the longissimus dorsi muscle, and recorded slaughter performance. Exercise was found to have a significant impact on the content of short-chain fatty acids in the colon. Additionally, it generally reduced the mRNA expression of fatty acid absorption transporter genes in the colonic epithelium and lipid metabolism-related genes in the longissimus dorsi muscle. Furthermore, exercise significantly affected the content of fatty acids in the longissimus dorsi muscle of Sunit lambs, and increased its dressing percentage. Exercise influences the composition of fatty acids of the longissimus dorsi muscle and improves dressing percentage by affecting the digestion, absorption, and metabolism of lipids in Sunit lambs. This could be profitable for the livestock industry, and could alter the nutrition and flavor of lamb meat.
Key words: lamb, absorption, slaughter performance, short-chain fatty acids, fatty acid
*Corresponding Authors: Yan Duan, College of Food Science and Engineering, Inner Mongolia Agricultural University, 306 Zhaowuda Road, Hohhot 010018, China. Email: [email protected]; Ye Jin, Integrative Research Base of Beef and Lamb Processing Technology, Ministry of Agriculture and Rural Affairs of the People’s Republic of China, Hohhot 010018, PR China. Email: [email protected]
Received: 30 October 2023; Accepted: 17 February 2024; Published: 16 May 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Sheep have become a preferred option for livestock farming because of expensiveness of their meat, low feeding costs, and adaptability. Among them, Sunit lambs raised in the Xilin Gol grassland of Inner Mongolia Autonomous Region in China are highly valued due to their high nutritional, protein, and low fat contents, making them popular with both herders and consumers (Yao et al., 2022). Captive breeding of sheep has become the mainstream approach to implementing sustainable development practices, resulting in significant improvement to the ecological environment of pastoral areas. Captive breeding is keeping sheep in a pen, as opposed to grazing in the open. Captive sheep spend their time in pens from birth to slaughter. However, captive breeding can result in decreased immunological capacity and nutritional quality of sheep meat because of factors, such as a lack of exercise (Joo et al., 2013).
Exercise is one of the critical factors affecting meat quality that demands attention. Research has shown that exercise has an effect on meat quality, particularly meat colour, with cattle raised in rough conditions having dark coloured muscles (Dunne et al., 2011).
Increase in exercise affects the metabolism of beef, resulting in decreased tenderness of meat. Hence, appropriate increase in livestock and poultry exercise has an important impact on meat quality, such as tenderness and meat colour (Gangnat et al., 2017). Two methods to improve the quality of meat are supplemental feeding (adding feed additives to captive sheep feeds) and exercise (expansion of feeding grounds to force the movement of captive sheep) to improve the intestinal system that affects the meat quality. However, the effects of exercise on the intestinal system have been focused on humans and small animals, and fewer studies have been reported in the ruminants.
Exercise and gastrointestinal flora can interact with each other (Hughes and Holscher, 2021), while some studies have reported that exercise can influence lipid metabolism in the body by affecting intestinal flora (Quiroga et al., 2020). Wang et al. (2021) found a significant correlation between rumen flora and muscle fatty acid (FA) composition in Sunit sheep, and a study found that the slaughter rate of sheep could be changed after changing their feed while changing their rumen flora (Wang et al., 2023). The mammalian gut harbours a vast and complex community of microorganisms, primarily comprising anaerobic bacteria with a small fraction of aerobic and facultative bacteria (Wen et al., 2021; Zimmermann and Curtis, 2019). The digestion and absorption of nutrients from feed occur in the intestinal tract, fueling various metabolic processes in the animal’s body. Consequently, there was a significant correlation between gastrointestinal flora and metabolites of the flora and quality of lamb meat (Du et al., 2022; Ma et al., 2023). Exercise is known to influence a variety of intestinal microbiota in animals (Rebec et al., 2022), and several studies have reported that insufficient animal exercise leads to reduced appetite and weight loss in livestock and poultry, predisposing them to diseases (Jang et al., 2017). The intestinal monolayer epithelium’s tolerance to the microbiota is critical, as it provides a physical barrier against intestinal pathogens, while the intestinal crypt is an invagination of intestinal epithelial cells (IECs) (Spence et al., 2011). Acetic acid has been shown to promote cupped cell mucin secretion (Wrzosek et al., 2013), while butyric acid enters colonic cells via transporter proteins, such as MCT1 and SCT1, and undergoes β-oxidation and tricarboxylic acid cycle metabolism. Furthermore, butyric acid promotes β-oxidation through signaling of intestinal epithelial cells by the peroxisome proliferator-activated receptor family (PPAR) (Dias et al., 2022) and induces Interleukin-10 receptor (IL-10RA) while inhibiting Claudin-2 to enhance epithelial barrier function and improve health of an organism (Zheng et al., 2017).
Short-chain fatty acids (SCFAs), such as acetic acid, propionic acid, and butyric acid, are a group of saturated fatty acids (SFAs) produced by anaerobic bacteria or yeasts that play a critical role in maintaining the normal function of the large intestine and the morphology and function of colonic epithelial cells. SCFAs are highly absorbable, with approximately 95% absorption (Carey and Montag, 2021). When activated as ligands for G protein-coupled receptors FFAR2 and FFAR3, acetate acid and propionate acid result in the release of peptide YY (PYY) by enteroendocrine cells, thereby slowing down intestinal transit time and enhancing the efficiency of energy extraction (Tazoe et al., 2008). Propionic acid, the most potent agonist of FFAR2 with an EC50 value of approximately 12–274 μM, has been linked to microbially induced obesity (Koh et al., 2016). Mice deficient in FFAR3 (Ffar3-/-) exhibited a 30% increase in body weight and a 25% decrease in body fat at the same amount of food intake (Samuel et al., 2008). SCFAs may act as signaling molecules that influence animals’ nutritional absorption.
Inducing exercise in mice has been shown to influence the abundance of butyrate-producing bacteria in the colon, leading to alterations in butyrate concentration and a decrease in body fat content (Yang et al., 2020). The microbiome of mice that exercised was more diverse and had more metabolic capacity than that of the control group (Hou et al., 2020). Exercise has the potential to modulate the composition of SCFAs, which in turn may influence lipid and energy metabolism. However, research on the effects of exercise on gut microbiota metabolites, intestinal absorption capacity, and fatty acid content in the longissimus dorsi muscle of ruminants is limited. We, therefore, suggest that exercise may affect lamb quality by affecting the digestion and absorption ability of the gut. The study measured SCFAs content in the colon, crypt depth, mucosal thickness of the colonic epithelium, mRNA expression of the genes related to fatty acid absorption, and fatty acid content of the longissimus dorsi muscle in captive Sunit sheep after 3 months of intervention. The aim of this study was to provide a scientific basis for the future optimization of sheep feeding practices to obtain lambs with higher nutritional value.
Materials and Methods
Animal care
Animal experiments were conducted according to the National Institute of Animal Health of Chinese Standards for Animal Experimentation (GB 14925-2010) and with the approval of the Inner Mongolia Agricultural University’s Ethics Committee (NND2021072). This study was carried out following the recommendations of the Instructive Notions concerning Caring for Experimental Animals, Ministry of Technology of China.
Animals and experimental design
Sunit lambs from Wulate Middle Banner, Inner Mongolia, with an average body weight of 20.27±1.90 kg were selected for the experiment. In all, 12 healthy 3-month-old lambs were randomly assigned into two groups: captive group (CG, basal diet) and captive with exercise group (EG, baseline diet with artificially increased exercise), each consisting of six lambs, with equal gender distribution. Each group was fed with free eating and drinking twice a day (d), once in the morning and again in the evening. As the age of lambs increased, the weight of fed concentrate also increased: 200 g of concentrate per day for 0–30 d, 400 g of concentrate per day for 30–60 d, and 600 g of concentrate per day for 60–90 d. The composition of the two groups’ diet is shown in Table 1. According to conducted pre-tests, the EG lambs were exercised on a field of 18.5×10.5 m2 at an average speed of 75 m/min. They were driven by the herders twice a day (10:30–11:10 am and 7:00–7:40 pm) for a total of 80 min (pedometer daily steps of 10,000+ and distance of 6 km+ prevailed). During the experimental period, the lambs were moved around the feed trough and permitted to drink freely. Pre-feed for 3 days, animals were slaughtered and sampled for analysis after 90 days of formal feeding.
Table 1. Basic diet and nutrient level (%).
| Classification | Composition | Content (%) |
|---|---|---|
| Composition of raw | Corn | 60.0 |
| Sunflower meal | 18.0 | |
| Bean cake | 13.5 | |
| Cottonseed meal | 4.0 | |
| Stone Powder | 1.0 | |
| Sodium chloride | 0.8 | |
| Calcium hydrogen phosphate | 0.5 | |
| Vitamin and Mineral premix | 1.5 | |
| Nutrient level | Crude protein | 18.0 |
| Crude fiber | 11.6 | |
| Coarse ash | 7.8 | |
| Calcium | 1.2 | |
| Phosphorus | 0.6 | |
| Lysine | 0.5 |
Note: Mineral vitamin premix provides per kilogram of diet: vitamin A, 4,000 IU; vitamin K3, 3,500 IU; vitamin B12, 2,500 IU; vitamin D3, 1,500 IU; vitamin E, 35 mg; zinc, 35 mg; iron, 35 mg; manganese, 20 mg; copper, 12 mg; iodine, 1.4 mg; and cobalt, 0.2 mg.
Animal sample collection
Both groups of lambs were transported on the same day to a slaughter house, 50 km from the farm (Grassland Hengtong Foods Co. Ltd., Bayannur, Inner Mongolia) and slaughtered by regular commercial halal methods. Before slaughtering Sunit lambs, the animals fasted for 24 h, their live weights were recorded, blood samples were collected and stored in etheylenediaminetetraacetic acid (EDTA) tubes, and immediately sent to the laboratory for centrifugation at low temperature (-4°C) at 3000×g for 10 min; the plasma was stored at -80°C for determining blood parameters.
After slaughtering the animals, the abdominal cavity was immediately opened and colonic contents were collected (0.5 g) and preserved at -80°C in 15-mL enzyme-free sterile centrifuge tubes for SCFAs analysis. The colonic epithelial tissues were trimmed into 2 × 1 cm2 (L × W), and one part was stored in 4% neutral formaldehyde solution for observation of colonic morphology while the other part was stored in 15 mL centrifuge tubes at -80°C for expression of fatty acid transport genes in the intestinal epithelium. The 1 × 0.5 × 0.5 cm3 (L × W × H) longissimus dorsi muscle blocks were clipped and collected in three tubes per lamb, and two blocks in each tube were frozen in 2-mL gene-freezing tubes at -80°C to determine muscle fatty acid metabolism gene expression. In addition, the longissimus dorsi (50 g) muscle on the 12th and 13th intercostal spine of each lamb was collected for analysis of fatty acid content and stored in self-sealing bags at -20°C.
Chemical analysis
Intestinal short-chain fatty acids
Weigh 0.5 g of colon contents into 15-mL centrifuge tubes, add 2 mL of ice-cold saline and 1 mL of 50% H2SO4 solution, and then vortex to mix. After centrifugation for 5 min (4°C, 10,000 r/min) while adding 2 mL of ether to mix, centrifuge for 5 min (4°C, 10,000 r/min). The supernatant was filtered through a 0.22-μm organic filter into a sample vial for gas chromatographic analysis following stratification. Following were the conditions for gas chromatography (Wang et al., 2021): DB-5 column (30 m × 0.25 mm, 0.25 μm), helium carrier gas flow rate of 1.2 mL/min, 250°C injection port temperature, 1-μL injection volume, 10:1 split ratio, and the flame ionization detector (FID). The following temperature programs was adopted: (1) the initial temperature was 100°C, which was maintained for 0.5 min; (2) the rate of 8°C/min was increased to 180°C and held for 1 min; (3) the rate of 20°C/min was increased to 200°C and maintained for 5 min. Conditions for mass spectrometry: ion source temperature of 250°C, transmission line temperature of 250°C, and mass scanning range of 40–450 m/z. Seven standard SCFAs were purchased from Aladdin Biochemical Technology Co (Shanhai, China), with >99.5% purity.
Intestinal mucosal morphology
The colon tissues were dried and fixed in paraffin to create 6-μm-thick paraffin slices. After staining with hematoxylin and eosin (H&E), the tissues were observed under light microscopy. The Leica Qwin V3 image analysis and processing system was used to measure the depth of crypt and mucosal thickness of the colon within the field of view. At least five fields of view in each section were selected for measurement, and the mean value was calculated as the measurement data.
Relative gene expression
Real-time quantitative polymerase chain reaction (RT-PCR) was used to determine the genes that regulate fatty acids in the colon and longissimus dorsi muscles: (1) stearovl-coenzyme a desaturase (SCD), fatty acid desaturases (FADS1 and FADS2), and fatty acid-binding protein 4 (FABP4); (2) intestinal regulation of fatty acid absorption and transport-related genes: fatty acid-binding protein 2 (FABP2), fatty acid transport protein 4 (FATP4), peroxisome proliferators activate receptors α (PPARα), monocarboxylate transporter 1 (MCT1), and the relative expression of sodium-coupled monocarboxylate transporter 1 (SMCT1) mRNA. Table 2 shows the sequences of gene primers whereas β-actin is the internal reference gene. The design and synthesis of gene primer sequences were completed by Shanghai Sangon Bioengineering Co. Ltd (Shanghai, China).
Table 2. Primers of RT-PCR.
| Genes | Primer sequence 5’-3’ | Fragment size (bp) |
|---|---|---|
| β-actin | F: 5’-CTCACGGAGCGTGGCTACA-3’ | 107 |
| R: 5’-GCCATCTCCTGCTCGAAGTC-3’ | ||
| FABP2 | F: 5’-CCTCACAGACGGAACTGAACTC-3’ | 123 |
| R: 5’-TCGCCACCTATAATTTCTCGGACAG-3’ | ||
| FATP4 | F: 5’-CATGGAGAACCGCAATGAGTTTGTG-3’ | 122 |
| R: 5’-GAGAGGAGGTCAGGCAGTGGAG-3’ | ||
| MCT1 | F: 5’-TCCCACTGAAGACACAGGCT-3’ | 139 |
| R: 5’-ACAGCACTGCAACACGAGAG-3’ | ||
| SMCT1 | F: 5’-TCTCGTGGGACTCTGGGTGA-3’ | 114 |
| R: 5’-TGGTCCGGTGCAGACACTTT-3’ | ||
| PPARα | F: 5’-CACAGCAGCCGTGTCGATTC-3’ | 98 |
| R: 5’-ACCCTCCCTGGACACCTCTT-3’ | ||
| SCD | F: 5’-GAGTACCGCTGGCACATCAA-3’ | 103 |
| R: 5’-CTAAGACGGCAGCCTTGGAT-3’ | ||
| FADS1 | F: 5’-CTGCTGTACCTGCTGCACAT-3’ | 161 |
| R: 5’-ACGGACAGGTGTCCAAAGTC-3’ | ||
| FADS2 | F: 5’-TGCCAACTGGTGGAACCATCGC-3’ | 188 |
| R: 5’-GCGGCCCGATCAGGAAGAAGTAC-3’ | ||
| FABP4 | F: 5’-AAGAAGTGGGTGTGGGCTTT-3’ | 91 |
| R: 5’-ATGTTGACCACATCCCCATT-3’ |
The samples kept at -80°C were put in a clean, pre-chilled mortar and pounded into powder. RNA extraction was conducted by the Hou method. The obtained RNA samples were extracted using 1% agarose gel electrophoresis to determine extraction quality, and OD values at A230, A260, and A280 nm were simultaneously measured to guarantee RNA’s purity. The ultimate concentration of RNA in each sample was 500 ng/μL. The extracted RNA was reverse-transcribed into complementary DNA (cDNA) using PrimeScript™ RT reagent kit with genomic deoxyribonucleic acid (gDNA) eraser. Finally, the obtained cDNA was stored at -20°C for use. RT-PCR was performed using a fluorescence quantitative kit (TB GreenTM Premix Ex TaqTM II). PCR reaction conditions for genes in the longissimus dorsi muscle were as follows: pre-denaturation (95°C, 30 s); denaturation (95°C, 5 s); annealing (58°C, 30 s); and extension (72°C, 30 s), for 45 cycles. PCR reaction conditions for intestinal genes were as follows: pre-denaturation (95°C, 30 s); denaturation (95°C, 5 s); annealing (58°C, 30 s); extension (72°C, 30 s), for 50 cycles. Reverse-transcription was performed using a Biometra instrument, followed by RT-PCR using a LightCycler® 96 (Roche, Basel, Switzerland).
Relative gene expression
Spectrophotometry was used to measure total cholesterol (TC), triglyceride (TG), low-density lipoprotein (LDL), and high-density lipoprotein (HDL) in the blood according to the instruction manual of the assay kit provided by the Nanjing Jiancheng Bioengineering Institute (Nanjing, Jiangsu, China).
Muscle long-chain fatty acids
A total of 5.0 g of homogenized longissimus dorsi muscle were weighed; a 2:1 combination of chloroform and methanol was added, stirred magnetically for 2 h, soaked for 10 h, and filtered using a G3 funnel. After adding a 5-mL mass fraction of 20% NaCl solution to the filtrate, the bottom chloroform layer (containing the fat extract) was preserved after standing. Anhydrous Na2SO4 was added for dehydration and rotational evaporation at 40°C to obtain fat concentration. Then, 5 mL of 20-g/L sodium hydroxide–methanol solution was added, refluxed at 70°C for 5 min for saponification, and 5 mL of boron trifluoride ether solution was added, and refluxed at 70°C for 2 min for methyl esterification. Finally, 2 mL of chromatographically pure hexane was added, the mixture was refluxed at 70°C for 1 min, 5 mL of saturated NaCl solution was added, and the mixture was allowed to stand for 10 min. The top layer of n-hexane was withdrawn by using a 1-mL syringe, filtered it using a 0.22-μm organic filter into the injection bottle to conduct gas chromatography analysis.
Improvements were made based on Wang et al. (2021). Gas chromatography was done using the SP-2560 gas chromatography column (100 m × 250 μm, 0.2 μm). The carrier gas was nitrogen, the injection port temperature was 250°C, and a volume of 1 L was injected at a split ratio of 25:1. The temperature program consisted of the following three steps: (1) the initial temperature was 100°C, after holding for 13 min, the temperature was increased to 180°C at 10°C/min and held for 6 min; (2) after the temperature was increased to 200°C at a rate of 1°C/min, it was stabilized for 20 min; and (3)heating was continued to 230°C at a rate of 4°C/min, and the temperature rise ended after holding for 20.5 min.
Slaughter performance
Live weight: after fasting for 24 h, the lambs were weighed before slaughter.
Carcass weight: fully exsanguinated after slaughter. The fur, head and hoof, lymph, and internal organs were removed and weighed after standing period of 30 min:
Net meat weight: remove carcass bones and the remaining net meat was weighed.
Carcass depth: measuring tape was used to measure vertical distance from end of the rib to the spine.
Back fat thickness: back fat thickness was measured immediately after slaughtering using a calliper.
Statistical analysis of data
Analysis of variance (ANOVA) test and Pearson’s correlation analysis were performed using the SPSS 23.0 software. Values were expressed as mean ± standard error (S.E.). The 2-ΔΔCt method was used to calculate the relative expression of gene mRNA. Histograms were plotted using the R (v4.1.1) and OriginPro (v9.8.0.200), and beautified using Adobe Illustrator. This resulted in six sample points for each group for the following parameters: slaughtering performance, SCFAs content, mRNA expression of colonic transporter genes, haematological parameters, lipid metabolism genes, and fatty acid content of the longest dorsal muscle. For the observation of the colonic epithelium, we selected five fields of view for each section and five positions for each field of view to record the depth of crypts and mucosal thickness. The average of the five positions was used to express the depth of crypts and mucosal thickness in that field of view. Therefore, there were 30 data points in each group of colonic epithelium data.
Results
Effects of exercise on intestinal metabolites and structure in Sunit lamb
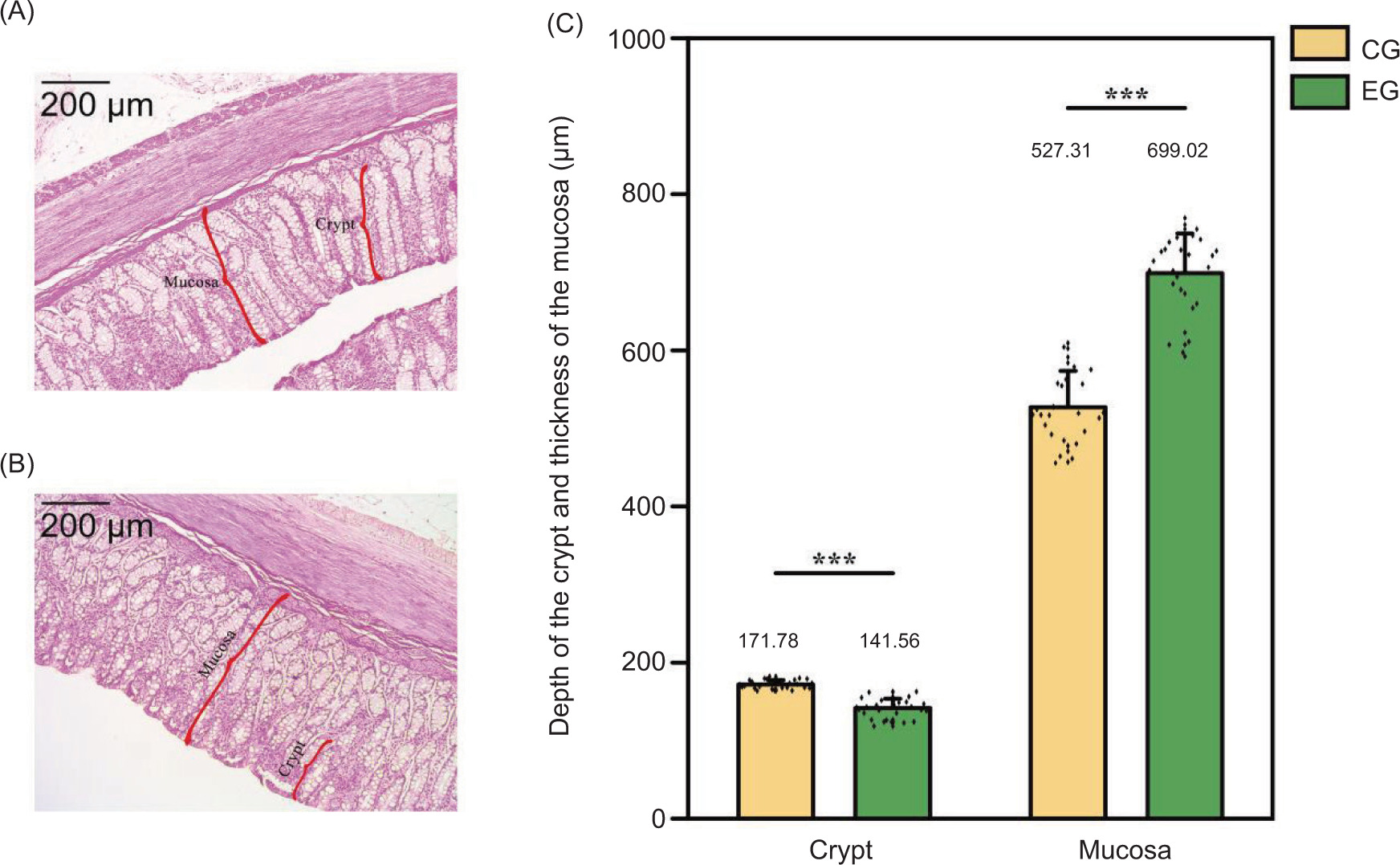
The levels of SCFAs in the colons of Sunit lamb under two feeding regimes were assessed, and the results are presented in Figure 1A. No significant differences in isovaleric acid and caproic acid were observed between the two groups (P > 0.05). However, the EG exhibited significantly lower concentrations of acetic acid, butyric acid, isobutyric acid, and valeric acid, compared to CG (P < 0.05), while the CG had significantly higher level of propionic acid (P < 0.05). Additionally, Sunit lambs in the EG had a significantly lower colonic pH (P < 0.05) (Figure 1B). The exercise regime also impacted the structure of the colonic mucosa as shown in Figures 2A–2C. Specifically, the EG exhibited a reduction in the depth of the intestinal crypt and a significant increase in mucosal thickness (P < 0.05).
Figure 1. Effects of exercise on the content of SCFAs in the colon of Sunit lamb (mg/100 g). CG: control group (basal feed), EF: exercise group (basal feed + exercise). *Significant difference between the two groups (P < 0.05); **significant difference between the two groups (P < 0.01); ***significant difference between the two groups (P < 0.001). (A) The content of SCFAs (mg/100 g) between both CG and EG groups; (B) difference in pH value between both CG and EG groups. SCFAs: short-chain fatty acids.
Figure 2. Effects of exercise on the colon structure of Sunit lamb. CG: control group (basal feed), EG: exercise group (basal feed + exercise). ***Significant difference between the two groups (P < 0.001). (A) Colonic structure of lamb in the captive group; (B) colonic structure of lamb in EG; (C) differences in the crypt and mucosa in the intestinal structure of both CG and EG groups.
Effect of exercise on the expression of fatty acid absorption and transport genes in the colon of Sunit lamb
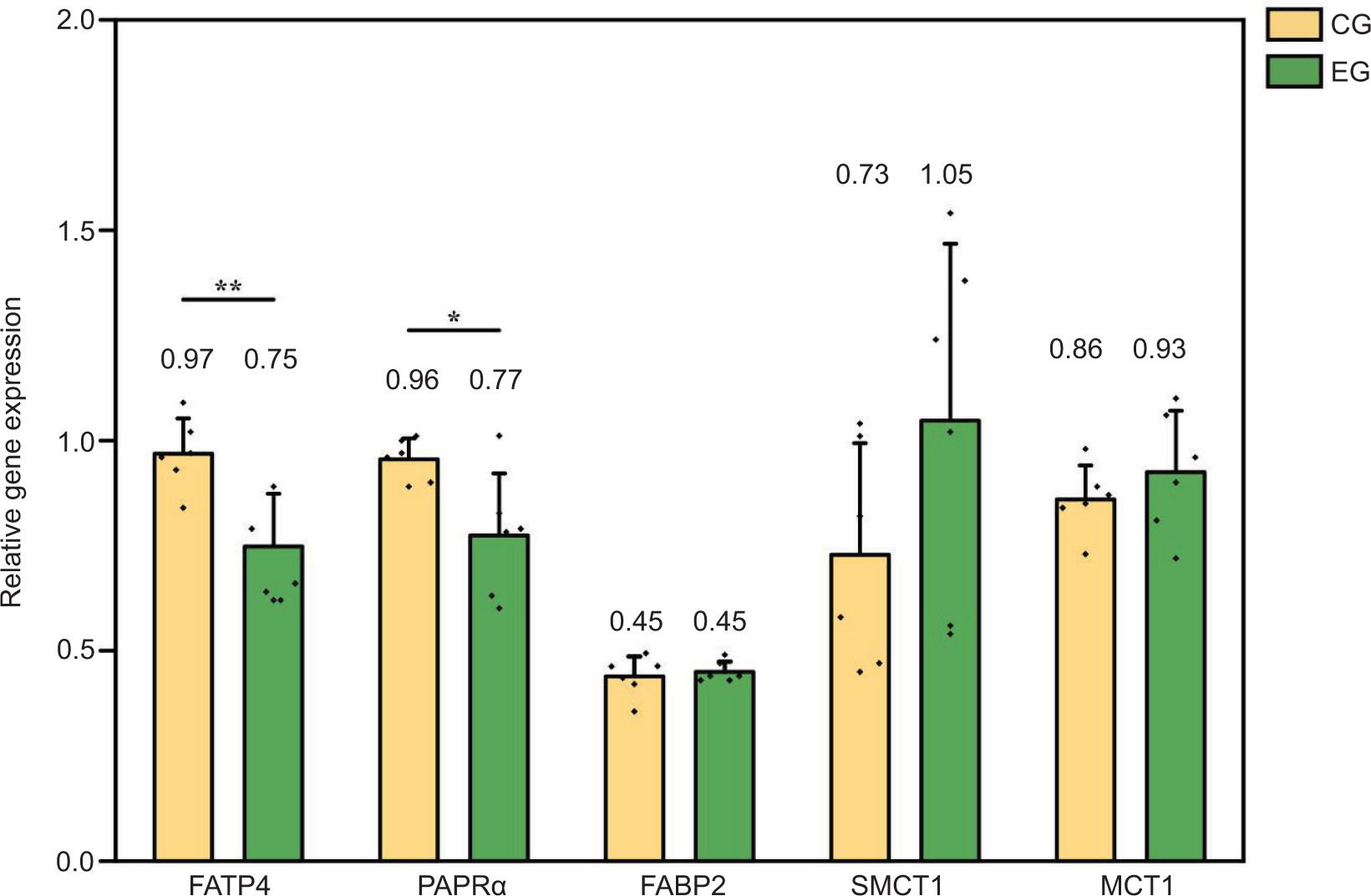
The exercise was found to have a significant impact on the expression of genes related to fatty acid absorption and transport in the colonic epithelium of Sunit lamb (Figure 3). The mRNA expression of FATP4 and PPARα genes was significantly higher in the CG, compared to the EG (P < 0.05). However, no significant differences were observed in the mRNA expression of colonic FABP2, SMCT1, and MCT1 between the two groups (P > 0.05). Exercise may affect the regulation of fatty acid absorption and transport in Sunit lamb, potentially impacting their nutritional value.
Figure 3. Effects of exercise on the expression of a fatty acid absorption transporter gene in the colon of Sunit lamb. CG: control group (basal feed), EF: exercise group (basal feed + exercise). *Significant difference between the two groups (P < 0.05); **significant difference between the two groups (P < 0.01).
Effect of exercise on blood parameters in Sunit lamb
Table 3 illustrates the observed differences between the EG and the CG concerning various lipid parameters. Notably, the EG displayed significantly higher levels of HDL, compared to the CG (P < 0.05). Moreover, plasma levels of TG, TC, and LDL were significantly lower in the EG by 23.50%, 9.52%, and 5.97%, respectively, compared to the CG (P < 0.05). Exercise can improve lipid metabolism and potentially reduce the risk of cardiovascular disease in Sunit lamb.
Table 3. Effect of exercise on blood parameters (mmol/L).
| Parameter | CG | EG |
|---|---|---|
| TG | 3.83±0.51a | 2.93±0.54b |
| TC | 7.88±0.42a | 7.13±0.38b |
| HDL | 1.16±0.15b | 1.59±0.22a |
| LDL | 0.67±0.06 | 0.63±0.03 |
a,bSignificant differences between feeding probiotics (P < 0.05).
LDL: low-density lipoprotein; HDL: high-density lipoprotein; TG: triglycerides; TC: total cholesterol.
Effect of gene expression of fatty acid metabolism in the longissimus dorsi muscle
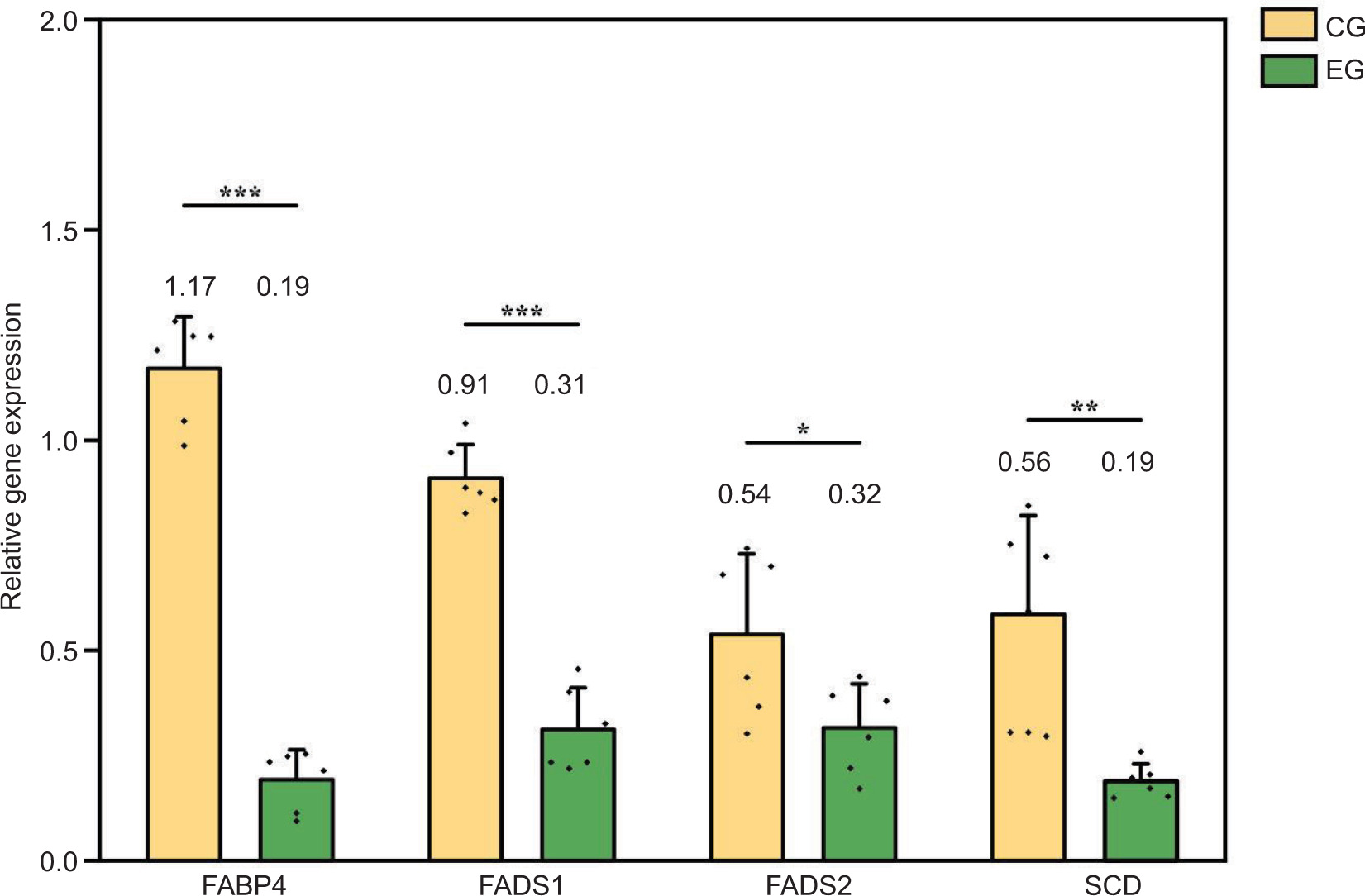
The present study aimed to investigate the effects of exercise on gene expression of fatty acid metabolism in the longissimus dorsi muscle of Sunit lamb, as shown in Figure 4. There had significant differences in the mRNA expression of FABP4, FADS1, FADS2, and SCD between both EG and CG. In the longissimus dorsi muscle of the EG, the mRNA expression of these genes was significantly lower than that of the CG (P < 0.05). Exercise may influence the regulation of fatty acid metabolism in Sunit lamb.
Figure 4. Effects of exercise on gene expression of fatty acid metabolism in the longissimus dorsi muscle of Sunit lamb. CG: control group (basal feed), EG: exercise group (basal feed + exercise). *Significant difference between the two groups (P < 0.05); **significant difference between the two groups (P < 0.01); and ***significant difference between the two groups (P < 0.001).
Fatty acid composition of longissimus dorsi muscle
The present study aimed to investigate the effect of exercise on fatty acid content in the longissimus dorsi muscle of Sunit lamb as presented in Table 4. In both EG and CG, stearic acid (C18:0) and palmitic acid (C16:0) accounted for 85–89% of SFA in the longissimus dorsi muscle. The main source of mutony taste was C18:0, which showed no significant difference in its content between the two groups (P > 0.05). However, the total SFA content of the longissimus dorsi muscle was significantly higher in the EG than in the CG (P < 0.05). Notably, lauric acid (C12:0), myristic acid (C14:0), pentadecanoic acid (C15:0), and palmitic acid (C16:0) were significantly increased, while arachidic acid (C20:0) was significantly decreased in the EG (P < 0.05).
Table 4. Effect of exercise on fatty acids of the longissimus dorsi muscle of Sunit lamb (%).
| Name | Structure | CG | EG | |
|---|---|---|---|---|
| SFA (saturated fatty acids) | Decanoic acid | C10:0 | 0.14±0.01b | 0.16±0.01a |
| Lauric acid | C12:0 | 0.11±0.01b | 0.15±0.01a | |
| Myristic acid | C14:0 | 2.19±0.22b | 2.49±0.18a | |
| Pentadecanoic acid | C15:0 | 0.32±0.02b | 0.56±0.08a | |
| Palmitic acid | C16:0 | 23.79±0.38b | 25.11±0.86a | |
| Stearic acid | C18:0 | 13.30±0.48 | 13.76±1.21 | |
| Arachidic acid | C20:0 | 0.12±0.01a | 0.10±0.01b | |
| Henicosanoic acid | C21:0 | 0.21±0.01a | 0.04±0.01b | |
| Behenic acid | C22:0 | 2.69±0.11 | 2.55±0.45 | |
| total | - | 42.86±0.32b | 44.93±2.11a | |
| MUFA (monounsaturated fatty acids) | Myristoleic acid | C14:1 | 0.35±0.02a | 0.21±0.02b |
| Palmitoleic acid | C16:1 | 2.19±0.34b | 2.69±0.05a | |
| Nervonic | C24:1 | 0.48±0.03a | 0.26±0.04b | |
| Elaidic acid | C18:1n9t | 1.36±0.09b | 1.95±0.18a | |
| Oleic acid | C18:1n9c | 42.44±0.50a | 41.49±0.49b | |
| total | - | 46.83±0.33 | 46.59±0.42 | |
| PUFA (polyunsaturated fatty acids) | Linolelaidic acid | C18:2n6t | 0.32±0.04a | 0.18±0.05b |
| Linoleic acid | C18:2n6c | 6.89±0.48 | 6.79±1.12 | |
| γ-linolenic acid | C18:3n6 | 0.36±0.01 | 0.34±0.03 | |
| α-linolenic acid | C18:3n3 | 0.73±0.07a | 0.42±0.08b | |
| cis-11,14-Eicosadienoic acid | C20:2 | 0.25±0.03 | 0.23±0.04 | |
| cis-8,11,14-Eicosatrienoic acid | C20:3n6 | 0.41±0.04a | 0.21±0.02b | |
| cis-11,14,17-Eicosatrienoic acid | C20:3n3 | 0.24±0.03 | 0.19±0.06 | |
| Arachidonic acid (AA) | C20:4n6 | 0.33±0.04a | 0.17±0.01b | |
| Eicosapentaenoic acid (EPA) | C20:5n3 | 0.36±0.02a | 0.28±0.01b | |
| Docosahexaenoic acid (DHA) | C22:6n3 | 0.17±0.01 | 0.15±0.02 | |
| total | - | 10.06±0.51 | 8.98±1.16 | |
| n-3 PUFA | - | 1.49±0.08a | 1.05±0.09b | |
| n-6 PUFA | - | 8.31±0.51 | 7.69±1.15 | |
| n-6/n-3 | - | 5.57±0.41b | 7.35±1.04a |
a,bSignificant differences between feeding probiotics (P < 0.05).
CG: control group (basal feed); EF: exercise group (basal feed + exercise).
Regarding monounsaturated fatty acids (MUFA), palmitoleic acid (C16:1) and elaidic acid (C18:1n9t) were significantly higher in the EG than in the CG (P < 0.05), while myristic acid (C14:1) and oleic acid (C18:1n9c) were significantly lower in the EG (P < 0.01). However, there was no significant difference in the total MUFA content of the longissimus dorsi muscle between the two groups (P > 0.05).
Polyunsaturated fatty acids (PUFA) were classified into n-3 and n-6 series based on the position of unsaturated bond. N-6 series mainly includes γ-linolenic acid (C18:3 n-6), cis-8,11,14-eicosatrienoic acid (C20:3 n-6), and arachidonic acid (AA). The total PUFA content of the longissimus dorsi muscle of Sunit lamb had no significant difference between the two groups (P > 0.05). The percentage of n-3 PUFA was significantly higher in the CG than in the EG (P < 0.05), but there was no significant change in the percentage of n-6 PUFA between the two groups (P > 0.05). Additionally, the n-6:n-3 ratio in the unsaturated fatty acids (UFA) was significantly higher in the EG, compared to the CG (P < 0.05). Exercise may influence the fatty acid composition of the longissimus dorsi muscle in Sunit lamb.
Correlation analysis of fatty acid metabolism-related genes and fatty acids
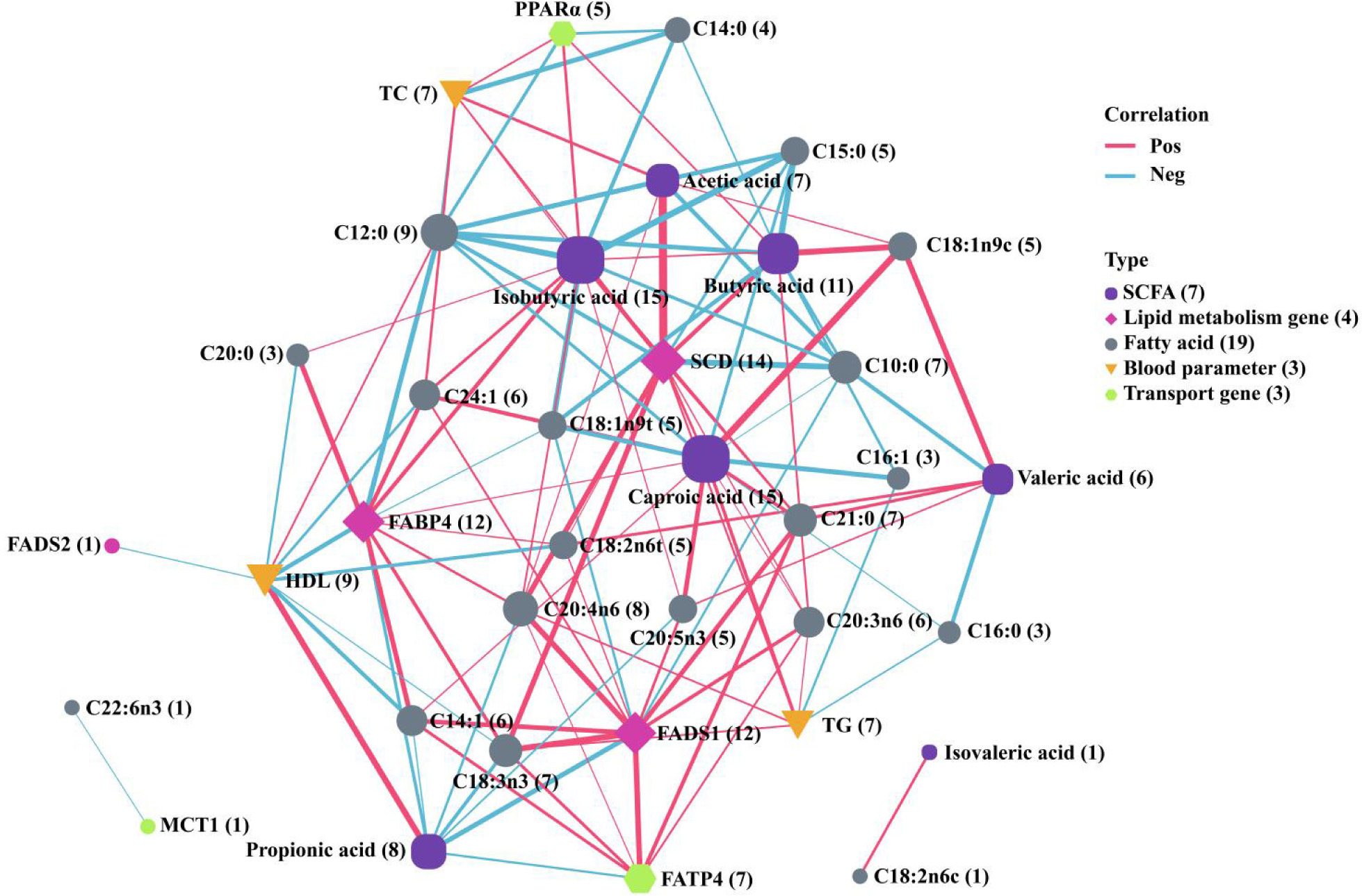
Figure 5 shows a network diagram of the correlation analysis between colonic SCFAs, intestinal epithelial fatty acid absorption and transport genes, blood parameters, longissimus dorsi muscle lipid metabolism genes, and longissimus dorsi muscle fatty acids. Only results of correlation analysis with |Spearman’s r| > 0.6 and P < 0.05 were retained. A total of seven SCFAs, four lipid metabolism genes, 19 fatty acids, three blood parameters, and three fatty acid uptake and transport genes were included. Six metrics were found correlated with more than 10 other metrics. These metrics are hexanoic acid, isobutyric acid, SCD, FABP4, FADS1, and butyric acid. Additionally, 15 other metrics were found correlated with both hexanoic acid and isobutyric acid. Of all the fatty acids, C12:0 was found to be the most susceptible, showing significant correlations with nine different metrics. These included four medium- and short-chain fatty acids (acetic acid, propionic acid, hexanoic acid, and isobutyric acid), two lipid metabolism genes (FABP4 and SCD), two blood parameters (HDL and TC), and PPARα.
Figure 5. Correlation analysis of SCFAs, fatty acid absorption and transport genes, lipid metabolism genes, blood parameters, and fatty acids. Correlations were analysed using Spearman’s correlation. Only correlations with |Spearman’s r| > 0.6 and P < 0.05 were retained and presented in the figure. Size of the dots in the graphs indicates the number of indicators associated with them (numbers in parentheses), and the shape and colour of the dots indicate their type. The thickness of the line indicates the magnitude of |Spearman’s r|: the thicker the line, the larger the |Spearman’s r|. Line colour indicates the positivity or negativity of Spearman’s r. Red line indicates a positive correlation between the two indicators in case Spearman’s r > 0, while a blue line indicates a negative correlation if Spearman’s r < 0.
Effect of exercise on slaughter performance of Sunit lamb
Table 5 presents the impact of exercise on the slaughter performance of Sunit lamb. There was no significant difference in carcass weight, carcass height, and net meat weight between the two groups of Sunit lamb (P > 0.05). However, the EG demonstrated significantly higher values for carcass depth and dressing percentage (P < 0.05), while back fat thickness was significantly lower, compared to the captive group (P < 0.05). These results may have important implications for the husbandry practices and management of Sunit lamb.
Table 5. Effects of exercise on slaughter performance and carcass quality of Sunit lamb.
| Index | CG | EG |
|---|---|---|
| Live weight (kg) | 36.78±2.37 | 35.40±3.18 |
| Carcass weight (kg) | 15.73±1.81 | 17.10±1.91 |
| Net meat weight (kg) | 12.22±2.48 | 13.86±1.40 |
| Carcass height (cm) | 73.50±2.17 | 72.67±1.86 |
| Carcass depth (cm) | 17.83±0.41b | 19.17±0.75a |
| Back fat thickness (mm) | 5.70±0.70a | 4.42±0.93b |
| Dressing percentage (%) | 44.48±4.28b | 50.38±2.24a |
a,bSignificant differences between feeding probiotics (P < 0.05).
CG: control group (basal feed); EF: exercise group (basal feed + exercise).
Discussion
The colon plays a crucial role in degrading undigested nutrients from the rumen and small intestine, resulting in higher concentrations of SCFA. The concentration of SCFAs in the colon reflects the extent of fermentation occurring in the animal’s hindgut (Carey and Montag, 2021). Exercise is one factor that influences the metabolites of intestinal flora, such as intestinal endocrine cells, which in turn affect intestinal digestion, barrier function, and host metabolism (Wrzosek et al., 2013). Previous studies have shown that exercise increases intestinal butyrate content in mice (Matsumoto et al., 2008), which is consistent with the findings of our investigation (P < 0.05). Butyrate has been shown to reduce intestinal inflammation and improve intestinal mucosal function (Hu et al., 2020). Our study demonstrated that exercise increased the thickness of colonic epithelial mucosa (P < 0.001) and SCFA concentration in the intestine while reducing the depth of colonic epithelial crypts (P < 0.001). This suggests that appropriate exercise promotes hindgut fermentation in Sunit lamb. The shallower the intestinal epithelial crypt, the better the maturation and secretory function of intestinal epithelial cells, while increased mucosal thickness improves intestinal immune function and promotes animal growth. An increase in the concentration of intestinal flora metabolites (SCFAs) may promote the synthesis and secretion of glucagon-like peptide 1 by colorectal epithelial cells, which in turn affects insulin secretion done by pancreatic B cells to regulate muscle cell glycogen metabolism (Tolhurst et al., 2012). Ultimately, this influences the muscular growth and development of Sunit lamb in EG. A decrease in intestinal pH in the EG (P < 0.05) may also inhibit the growth and reproduction of pathogenic Aspergillus bacteria. In conclusion, our study shows that exercise improves the absorption capacity of colon bacteria in Sunit lamb, which has a positive impact on both intestinal and the overall health of the organism.
The expression of lipid metabolism-related genes in Sunit lamb intestinal epithelial cells was found to regulate fatty acid metabolism and absorption. Among these genes, PPARα (P < 0.05), a key player in lipid metabolism, is involved in various biological processes, such as fatty acid transport and oxidation (Dias et al., 2022). Knockdown of intestinal PPARα in mice fed with a high-fat diet has been shown to limit body fat accumulation (Stojanović et al., 2021). FATP4 is a family of intestinal fatty acid transporters that regulates the absorption and transport of long-chain fatty acids (more than 18 carbons) by intestinal epithelial cells (Larqué et al., 2006). The mRNA expression of colonic FATP4 was found to be decreased in the EG (P < 0.01), inhibiting the absorption and transport of PUFA. SMCT1 (P > 0.05) and MCT1 (P > 0.05), on the other hand, are involved in the transport and absorption of nutrients, such as intestinal SCFA and amino acids (Izuddin et al., 2019). Therefore, the present study suggests that exercise regulates the expression of genes related to fatty acid metabolism in the colon of Sunit lamb, affecting the absorption and transport of long-chain fatty acids.
Blood parameters, such as TC, TG, HDL, and LDL are essential indicators of lipid metabolism in livestock. Abnormal lipid metabolism is often associated with increased levels of plasma TC and TG. Moderate exercise intervention has been shown to improve inflammatory cytokines and regulate lipid levels in mice fed with a high-fat diet (Semeraro et al., 2022). The current investigation found that exercise reduced plasma TC (P < 0.05) and TG (P < 0.05) levels in Sunit lamb, indicating that exercise improves lipid metabolism in animals and potentially benefit human nutritional health.
Significant correlations (P < 0.05) were observed between the seven SCFAs and other indicators throughout the correlation network. Each Hexanoic acid (15 indicators), isobutyric acid (15), and butyric acid (11) was correlated with more than 10 indicators. Acetic acid was found to be significantly different between the two groups (P < 0.01). Correlation analysis showed a significant positive correlation between acetic acid and TC, which was also significantly reduced in EG. Elevated levels of acetic acid are considered an indicator of obesity (Li et al., 2023), which leads to increased blood TC. Exercise has been shown to have a positive effect on lowering TC (Mann et al., 2014), which is consistent with the results of the present study. Therefore, it is suggested that exercise helps to improve the health status of Sunit lamb. A significant positive correlation (P < 0.05) was found between acetic acid and SCD, a key enzyme in the desaturation of fatty acids (Fu et al., 2021). The gene expression of SCD was significantly reduced in the dorsal longest muscle of the EG (P < 0.01), resulting in a reduced conversion of saturated to UFAs in the muscle of Sunit lambs. As a result, the proportion of SFAs increased in the EG. The correlation between acetic acid and fatty acids was also verified. Acetic acid exhibited a significant negative correlation with C18:0, C12:0, and C15:0, and a significant positive correlation with UFAs, such as C18:1n9c and C20:4n6.
In this study, it was found that levels of propionic acid were significantly higher in the colon of EG (P < 0.05) and were positively correlated with levels of HDL (P < 0.05). Additionally, levels of HDL were significantly higher in the EG group (P < 0.05), which is consistent with the findings of Zha et al. (2023) and Wang et al. (2023). It has been found that HDL can play a protective role in the body (von Eckardstein et al., 2023). HDL is able to transport excess cholesterol from peripheral tissues or cells to the liver for breakdown (Teng et al., 2020), which improves the health of Sunit lamb. Research has shown that propionic acid inhibits the expression of Npc1l1, a crucial cholesterol transporter in the intestine. This is achieved by affecting the number of T cells and the level of IL10 in the intestinal microenvironment (Haghikia et al., 2022). This pathway shows how propionic acid affects cholesterol metabolism in the body. Propionic acid was negatively correlated with the genes involved in lipid metabolism in the muscle (FABP4 and FADS1) and a lipid transporter gene in the gut (FATP4) (P < 0.05). FABP4 is associated with fat deposition (Michal et al., 2006), while the FADS family regulates PUFA synthesis (Hayashi et al., 2021), and FATP4 is responsible for the transport and uptake of long-chain fatty acids in the intestine (more than 18 carbons) (Stahl, 2004). In this study, it was found that the expression of all three genes was significantly down-regulated. The fatty acids that were correlated with these genes were mostly correlated positively. Additionally, both of these fatty acids were significantly reduced in the longest dorsal muscle. Based on these findings, we hypothesize that propionic acid can influence the meat quality of Sunit lamb by affecting absorption of intestinal cholesterol and composition of muscle fatty acid.
The study found a significant reduction in butyric acid in the EG (P < 0.05). Additionally, a positive correlation was observed between the transporter gene PPARα and lipid metabolism genes in the intestine (P < 0.05). PPARα is involved in various biological processes, including lipid metabolism, fatty acid transportation, and oxidation. It has been discovered that knocking down of intestinal PPARα in mice on a high-fat diet can inhibit dilation of the intestinal crypts and shortening of the villi, which in turn limits increase of body fat (Stojanović et al., 2021). This finding suggests a potential role for PPARα in regulating body fat. Similarly, a decrease in colonic PPARα expression in EG Sunit lamb has been found to limit body fat deposition and restrict cecal crypt expansion.
Isobutyric acid showed significant correlation with 15 indicators and was significantly reduced in the EG (P < 0.05). Additionally, it was significantly positively correlated with PPARα, FABP4, and SCD (P < 0.05). In our study, we found that isobutyric acid was significantly correlated with 11 fatty acids, including six SFAs and five UFAs. Among the six SFAs, isobutyric acid was significantly negatively correlated with four of them (P < 0). Among UFAs, isobutyric acid showed significant positive correlation with four of them (P < 0.05). Additionally, one study found a significant positive correlation between isobutyric acid and UFA/SFA. It is important to note that all evaluations presented are objective and based on empirical evidence (Ye et al., 2020).
The intervention of exercise in this study was found to reduce dorsal fat deposition as measured by a decrease in back fat thickness in the EG. Lambs with higher carcass weights are known to be more productive and competitive in the market (Volpi-Lagreca et al., 2021).
This study examines the impact of exercise on the meat quality of Sunit lamb, specifically investigating the effects on SCFAs, fatty acids, and slaughter performance. While SCFAs were analysed in the colon of Sunit lamb in this experiment, the factors influencing their content remain unclear. Therefore, in next experiments, we plan to perform metagenome sequencing of the colonic flora of Sunit lamb to investigate changes in colonic flora and function. We also plan to monitor the intake of Sunit lamb to observe the effect of exercise on intake in order to clarify the reasons for the effect of SCFAs content. Additionally, we want to investigate the effects of exercise on meat quality and nutritional indexes as well as slaughtering performance and muscle fatty acids.
Conclusions
Exercise had a significant effect on the content of SCFAs in the colon of Sunit lamb. It also lowered the pH of the colon and increased the thickness of the mucosa while lowering the depth of colonic crypts. These changes had a positive effect on the digestion and absorption of Sunit lamb. Furthermore, exercise significantly impacted the lipid metabolism of Sunit lamb. Specifically, the fatty acid absorption and transfer genes in the intestines, lipid metabolism genes in the longissimus dorsi muscle of Sunit lamb, and blood parameters were generally reduced. This is also reflected in changes in fatty acid content of the longissimus dorsi muscle of Sunit lamb. Exercise significantly increases the dressing percentage of Sunit lamb, with a positive impact on growth of the livestock industry and income. Additionally, exercise improves dressing percentage and health of Sunit lamb. However, the effect of exercise on fatty acids can further impact the nutritional value and flavour of mutton.
REFERENCES
Carey, R.A., and Montag, D., 2021. Exploring the relationship between gut microbiota and exercise: short-chain fatty acids and their role in metabolism. BMJ Open Sport & Exercise Medicine 7(2): e000930. 10.1136/bmjsem-2020-000930
Dias, B.V., Gomes, S.V., da Cruz Castro, M.L., Carvalho, L.C.F., Breguez, G.S., de Souza, D.M.S., et al. 2022. EPA/DHA and linseed oil have different effects on liver and adipose tissue in rats fed with a high-fat diet. Prostaglandins & Other Lipid Mediators 159: 106622. 10.1016/j.prostaglandins.2022.106622
Du, E., Guo, W., Zhao, N., Chen, F., Fan, Q., Zhang, W., et al. 2022. Effects of diets with various levels of forage rape (Brassica napus) on growth performance, carcass traits, meat quality and rumen microbiota of Hu lambs. Journal of the Science of Food and Agriculture 102(3): 1281–1291. 10.1002/jsfa.11466
Dunne, P.G., Monahan, F.J., and Moloney, A.P., 2011. Current perspectives on the darker beef often reported from extensively managed cattle: does physical activity play a significant role? Livestock Science 142(1–3): 1–22.
Fu, J., Wang, Y., Tan, S., and Wang, J., 2021. Effects of banana resistant starch on the biochemical indexes and intestinal flora of obese rats induced by a high-fat diet and their correlation analysis. Frontiers in Bioengineering and Biotechnology 9: 575724. 10.3389/fbioe.2021.575724
Gangnat, I.D.M., Leiber, F., Dufey, P.A., Silacci, P., Kreuzer, M., and Berard, J., 2017. Physical activity, forced by steep pastures, affects muscle characteristics and meat quality of suckling beef calves. Cambridge University Press, Cambridge UK, p. 2.
Haghikia, A., Zimmermann, F., Schumann, P., Jasina, A., Roessler, J., Schmidt, D., et al. 2022. Propionate attenuates atherosclerosis by immune-dependent regulation of intestinal cholesterol metabolism. European Heart Journal 43(6): 518–533. 10.1093/eurheartj/ehab644
Hayashi, Y., Lee-Okada, H.-C., Nakamura, E., Tada, N., Yokomizo, T., Fujiwara, Y., et al. 2021. Ablation of fatty acid desaturase 2 (FADS2) exacerbates hepatic triacylglycerol and cholesterol accumulation in polyunsaturated fatty acid-depleted mice. FEBS Letters 595(14): 1920–1932. 10.1002/1873-3468.14134
Hou, Y., Su, L., Su, R., Luo, Y., Wang, B., Yao, D., et al. 2020. Effect of feeding regimen on meat quality, MyHC isoforms, AMPK, and PGC-1α genes expression in the biceps femoris muscle of Mongolia sheep. Food Science & Nutrition 8(5): 2262–2270. 10.1002/fsn3.1494
Hu, B., Ye, C., Leung, E.L.-H., Zhu, L., Hu, H., Zhang, Z., et al. 2020. Bletilla striata oligosaccharides improve metabolic syndrome through modulation of gut microbiota and intestinal metabolites in high fat diet-fed mice. Pharmacological Research, 159: 104942. 10.1016/j.phrs.2020.104942
Hughes, R. L., and Holscher, H. D., 2021. Fueling gut microbes: a review of the interaction between diet, exercise, and the gut microbiota in athletes. Advances in Nutrition 12(6): 2190–2215. 10.1093/advances/nmab077
Izuddin, W.I., Loh, T.C., Samsudin, A.A., Foo, H.L., Humam, A.M., and Shazali, N., 2019. Effects of post-biotic supplementation on growth performance, ruminal fermentation and microbial profile, blood metabolite and GHR, IGF-1 and MCT-1 gene expression in post-weaning lambs. BMC Veterinary Research 15(1): 315. 10.1186/s12917-019-2064-9
Jang, H.J., Kim, D.M., Kim, K.B., Park, J.W., Choi, J.Y., Oh, J.H., et al. 2017. Analysis of metabolomic patterns in thoroughbreds before and after exercise. Asian-Australasian Journal of Animal Sciences 30(11): 1633–1642. 10.5713/ajas.17.0167
Joo, S.T., Kim, G.D., Hwang, Y.H., and Ryu, Y.C., 2013. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Science, 95(4): 828–836. 10.1016/j.meatsci.2013.04.044
Koh, A., De Vadder, F., Kovatcheva-Datchary, P., and Bäckhed, F., 2016. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 165(6): 1332–1345. 10.1016/j.cell.2016.05.041
Larqué, E., Demmelmair, H., Klingler, M., De Jonge, S., Bondy, B., and Koletzko, B., 2006. Expression pattern of fatty acid transport protein-1 (FATP-1), FATP-4 and heart-fatty acid binding protein (H-FABP) genes in human term placenta. Early Human Development 82(10): 697–701. 10.1016/j.earlhumdev.2006.02.001
Ma, Y., Han, L., Zhang, S., Zhang, X., Hou, S., Gui, L., et al. 2023. Insight into the differences of meat quality between Qinghai white Tibetan sheep and black Tibetan sheep from the perspective of metabolomics and rumen microbiota. Food Chemistry: X 19: 100843. 10.1016/j.fochx.2023.100843
Mann, S., Beedie, C., and Jimenez, A., 2014. Differential effects of aerobic exercise, resistance training and combined exercise modalities on cholesterol and the lipid profile: review, synthesis and recommendations. Sports Medicine 44(2): 211–221. 10.1007/s40279-013-0110-5
Matsumoto, M., Inoue, R., Tsukahara, T., Ushida, K., Chiji, H., Matsubara, N., et al. 2008. Voluntary running exercise alters microbiota composition and increases n-butyrate concentration in the rat cecum. Bioscience, Biotechnology and Biochemistry, 72(2): 572–576. 10.1271/bbb.70474
Michal, J.J., Zhang, Z.W., Gaskins, C.T., and Jiang, Z., 2006. The bovine fatty acid binding protein 4 gene is significantly associated with marbling and subcutaneous fat depth in Wagyu×Limousin F2 crosses. Animal Genetics 37(4): 400–402. 10.1111/j.1365-2052.2006.01464.x
Quiroga, R., Nistal, E., Estébanez, B., Porras, D., Juárez-Fernández, M., Martínez-Flórez, S., et al. 2020. Exercise training modulates the gut microbiota profile and impairs inflammatory signaling pathways in obese children. Experimental & Molecular Medicine 52(7): 1048–1061. 10.1038/s12276-020-0459-0
Rebec, G.V., Koceja, D.M., and Bunner, K. D., 2022. Measuring movement in health and disease. Brain Research Bulletin 181: 167–174. 10.1016/j.brainresbull.2022.01.021
Samuel, B.S., Shaito, A., Motoike, T., Rey, F.E., Backhed, F., Manchester, J.K., et al. 2008. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proceedings of the National Academy of Sciences 105(43): 16767–16772. 10.1073/pnas.0808567105
Semeraro, M.D., Almer, G., Kaiser, M., Zelzer, S., Meinitzer, A., Scharnagl, H., et al. 2022. The effects of long-term moderate exercise and western-type diet on oxidative/nitrosative stress, serum lipids and cytokines in female Sprague Dawley rats. European Journal of Nutrition 61(1): 255–268. 10.1007/s00394-021-02639-4
Spence, J.R., Lauf, R., and Shroyer, N.F., 2011. Vertebrate intestinal endoderm development. Developmental Dynamics 240(3): 501–520. 10.1002/dvdy.22540
Stahl, A., 2004. A current review of fatty acid transport proteins (SLC27). Pflügers Archives 447(5): 722–727. 10.1007/s00424-003-1106-z
Stojanović, O., Altirriba, J., Rigo, D., Spiljar, M., Evrard, E., Roska, B., et al. 2021. Dietary excess regulates absorption and surface of gut epithelium through intestinal PPARα. Nature Communications 12(1): 7031. 10.1038/s41467-021-27133-7
Tazoe, H., Otomo, Y., Kaji, I., Tanaka, R., and Kuwahara, A., 2008. Roles of short-chain fatty acids receptors, GPR41 and GPR43 on colonic functions. Journal of Physiology & Pharmacology 59(Suppl 2): 251–262.
Teng, Y., Wang, Y., Tian, Y., Chen, Y.Y., Guan, W.Y., Piao, C.H., et al. 2020. Lactobacillus plantarum LP104 ameliorates hyperlipidemia induced by AMPK pathways in C57BL/6N mice fed high-fat diet. Journal of Functional Foods 64: 103665. 10.1016/j.jff.2019.103665
Tolhurst, G., Heffron, H., Lam, Y.S., Parker, H.E., Habib, A.M., Diakogiannaki, E., et al. 2012. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the g-protein–coupled receptor FFAR2. Diabetes 61(2): 364–371. 10.2337/db11-1019
Volpi-Lagreca, G., Gelid, L.F., Alende, M., Bressan, E.R., Pordomingo, A.B., and Pordomingo, A.J., 2021. Effect of placement weight and days on feed on feedlot cattle performance and carcass traits. Livestock Science 244: 104392. 10.1016/j.livsci.2020.104392
von Eckardstein, A., Nordestgaard, B.G., Remaley, A.T., and Catapano, A.L., 2023. High-density lipoprotein revisited: biological functions and clinical relevance. European Heart Journal 44(16): 1394–1407. 10.1093/eurheartj/ehac605
Wang, B., Luo, Y., Wang, Y., Wang, D., Hou, Y., Yao, D., et al. 2021. Rumen bacteria and meat fatty acid composition of Sunit sheep reared under different feeding regimens in China. Journal of the Science of Food and Agriculture 101(3): 1100–1110. 10.1002/jsfa.10720
Wang, D., Tang, G., Yu, J., Li, Y., Feng, L., Liu, H., et al. 2023. Microbial enterotypes shape the divergence in gut fermentation, host metabolism and growth rate of young goats. Microbiology Spectrum 11(1): e04818–e04822. 10.1128/spectrum.04818-22
Wen, K., Zhao, M.M., Liu, L., Khogali, M.K., Geng, T.Y., Wang, H.R., et al. 2021. Thiamine modulates intestinal morphological structure and microbiota under subacute ruminal acidosis induced by a high-concentrate diet in Saanen goats. Animal 15(10): 100370. 10.1016/j.animal.2021.100370
Wrzosek, L., Miquel, S., Noordine, M.-L., Bouet, S., Chevalier-Curt, M.J., Robert, V., et al. 2013. Bacteroides thetaiotaomicron and Faecalibacterium prausnitziiinfluence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biology 11(1): 61. 10.1186/1741-7007-11-61
Wang, Y.C., Wangm X., Li, J.Z., Huang, P.F., Li, Y.L., Ding, X.Q., Huangm J., et al., 2023. The impact of lactating Hu sheep’s dietary protein levels on lactation performance, progeny growth and rumen development. Animal Biotechnology 34(6): 1919–1930. 10.1080/10495398.2022.2058006
Yang, J.J., Pham, M.T., Rahim, A.R., Chuang, T.-H., Hsieh, M.-F., and Huang, C.-M., 2020. Mouse abdominal fat depots reduced by butyric acid-producing leuconostoc mesenteroides. Microorganisms 8(8): 1180. 10.3390/microorganisms8081180
Yao, D., Su, R., Zhang, Y., Wang, B., Hou, Y., Luo, Y., et al. 2022. Impact of dietary Lactobacillus supplementation on intramuscular fat deposition and meat quality of Sunit sheep. Journal of Food Biochemistry 46(8): e14207.
Ye, Z., Cao, C., Li, Q., Xu, Y.J., Liu, Y., 2020. Different dietary lipid consumption affects the serum lipid profiles, colonic short chain fatty acid composition and the gut health of Sprague Dawley rats. Food Res Int. 132: 109117. 10.1016/j.foodres.2020.109117.
Li, Z.C., Liu, D.H., Gu, R.C., Qiao, Y., Jin, Q., Zhang, Y.J., et al. 2023. Fecal microbiota transplantation in obesity metabolism: A meta analysis and systematic review. Diabetes Res Clin Pract 202: 110803. 10.1016/j.diabres.2023.110803.
Zha, A., Tan, B., Wang, J., Qi, M., Deng, Y., Li, R., et al. 2023. Dietary supplementation modified attapulgite promote intestinal epithelial barrier and regulate intestinal microbiota composition to prevent diarrhea in weaned piglets. International Immunopharmacology 117: 109742. 10.1016/j.intimp.2023.109742
Zheng, L., Kelly, C.J., Battista, K.D., Schaefer, R., Lanis, J.M., Alexeev, E.E., et al. 2017. Microbial-derived butyrate promotes epithelial barrier function through IL-10 receptor–dependent repression of claudin-2. Journal of Immunology 199(8): 2976–2984. 10.4049/jimmunol.1700105
Zimmermann, P., and Curtis, N., 2019. The effect of antibiotics on the composition of the intestinal microbiota–a systematic review. Journal of Infection 79(6): 471–489. 10.1016/j.jinf.2019.10.008