
Download

RESEARCH ARTICLE
Antimicrobial activities of polyphenol-based metabolites present in lettuce and recent methods for their estimation
Jinghua Liu1, Zhidi Chen1,2†, Fengbo Ma1, Dongming Liu1, Li-Byarlay Hongmei3, Xuanzhe Chang1, Yanyan Zhang1, Xiangning Chen1, Xiuzhi Gao1*
1Beijing Key Laboratory of Agricultural Product Detection and Control of Spoilage Organisms and Pesticide Residue, Beijing Laboratory of Food Quality and Safety, Key Laboratory of Agricultural Product Processing and Quality Control (Co-construction by Ministry and Province), Ministry of Agriculture and Rural Affairs, Beijing University of Agriculture, Beijing, China;
2State Key Laboratory for Crop Stress Resistance and High-Efficiency Production, Shaanxi Key Laboratory of Agricultural and Environmental Microbiology, College of Life Sciences, Northwest A & F University, Yangling, China;
3Agricultural Research and Development Program, Central State University, Wilberforce, Ohio, USA
†Zhidi Chen contributed equally to this work.
Abstract
The tissue structure of fresh-cut lettuce is easy to be damaged during processing and transportation, which leads to the reduction of its edible quality and safety. Ultra-high-performance liquid chromatography (UHPLC) combined with time-of-flight tandem mass spectrometry (TMS) metabolomics was used to identify the differential metabolites related to the antibacterial activity in lettuce and explore their bacteriostatic mechanism. The experimental results revealed that the growth of Escherichia coli was quite different when different varieties of lettuce were used as the nutrient substrate. Between the varieties Beizisheng No. 3 (BZ3) and Shooter 101 (SS), 204 differential metabolites showed significant changes (P < 0.05), 86 metabolites showed changes between the varieties BZ3 and Beisansheng No. 1 (BS1). Among the metabolites, isoquercitrin was the upregulated differential metabolite in BZ3 compared to SS and BS1. The content of isoquercitrin in lettuce juice positively correlated with the antibacterial activity of polyphenol extract, but negatively correlated with the growth of E. coli. The bacteriostatic property of polyphenol extract destroys the morphological structure of E. coli. The higher the isoquercitrin content, the stronger the resistance of fresh-cut lettuce to E. coli.
Key words: antibacterial activity, Escherichia coli, lettuce, metabolomics
*Corresponding Author: Xiuzhi Gao, Beijing University of Agriculture, Beijing 102206, China. Email address: [email protected]
Received: 18 January 2024; Accepted: 10 September 2024; Published: 14 October 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Lettuce, also known as Lactuca sativa L., is a biennial herb of the family Asteraceae. It is a very popular ready-to-eat vegetable grown worldwide (López et al., 2014). Lettuce contains various nutrients, such as phenols, vitamin C, folic acid, carotenoids, and chlorophyll (Yang et al., 2022). It has a good nutritional value, prevents cardiovascular diseases (Shi et al., 2022), is antidiabetic (Cheng et al., 2014b), increases gastrointestinal motility to aid weight loss (Cheng et al., 2014a), and prevents cancer (Sularz et al., 2021). Lettuce is abundant in phytochemicals, including flavonoids, anthocyanins, and terpenes. Dannehl et al. (2016) found that red leaf lettuce contains one phenolic acid, five caffeic acid derivatives, three flavonols and their aglycones, and one anthocyanin.
Llorach et al. (2008) identified the phytochemicals in different lettuce varieties based on HPLC-DAD-MS/MS and found two compounds, quercetin and luteolin rhamnosyl hexoside, which were not reported in lettuce previously.
Plants contain a variety of secondary metabolites, among which the higher contents of polyphenols (El Moussaoui et al., 2019), flavonoids (Muna et al., 2022; Soberón et al., 2020), and terpenoids (Ketut Srie and Purnawati, 2023) in lettuce have been proved to have antibacterial activities. Pepe et al. (2014) found that leaf lettuce contains 16 polyphenol compounds all of which have anti-inflammatory and antioxidant activities. Moodi et al. (2021) showed that quercetin extracted from Ginkgo biloba combined with metal ions can significantly improve the inhibition of Staphylococcus aureus and Escherichia coli. Anthocyanins are also widely used in the antibacterial field. Li et al. (2022) showed that the lowest inhibitory concentrations of acylated anthocyanins in legumes against S. aureus and E. coli were 0.312 mg/mL and 4.000 mg/mL, respectively. Lettuce and Atractylodes macrocephala belong to the Asteraceae family. Zhaoyong et al. (2024) showed that the essential oil of A. macrocephala had various inhibitory effects on the growth of E. coli, Pseudomonas aeruginosa, Salmonella enteritidis, S. aureus, and Bacillus subtilis. van Treuren et al. (2018) determined the metabolic spectrum of phytochemicals in 150 lettuce samples. For many phytochemicals, the relative abundance was either positively or negatively correlated with available phenotypic data on resistance against pests and diseases, indicating their potential role in plant resistance.
Microorganisms have complex sources and easy reproduction characteristics, making them the primary risk factors for food raw materials, and processed and finished products (Zaman et al., 2024). In lettuce and its processed products, intestinal pathogens, such as E. coli (Tahir et al., 2022), Salmonella (Min et al., 2016), and Listeria monocytogenes (Shenoy et al., 2017), are the common causes of food poisoning. Mechanical damage during processing of fresh-cut lettuce often destroys the tissue, leading to microbial contamination (Lin et al., 2019). Microbial contamination of green leafy vegetables has caused at least 800 illnesses and eight deaths since 1993, particularly California-grown lettuce (iceberg, romaine, red leaf, and mesclun). Although cases of L. monocytogenes infection involving lettuce are rare, since 2010, eight recalls have been issued for green leafy vegetables infected with the microbe (Zeng et al., 2014). Due to the perishable nature of fresh-cut lettuce, air-conditioned packaging helps maintain its quality and safety (Islam et al., 2019). E. coli is a bacterium commonly found in the human gut and is a recognized marker of fecal contamination. It adheres to the surface of the lettuce through irrigation water (fertilizer), animal manure, or human pollution and reproduces through the plant juice lost during processing. The damages or wounds caused during processing invades the lettuce, causing contamination of the leaves as well as the entire processing line (Cuggino et al., 2023). The microorganisms also secrete toxins during their reproduction process, which could endanger the consumers’ health (Cui et al., 2018; Tang et al., 2017), causing diarrhea, urinary tract infections, arthritis, and septic infections (van Hoek et al., 2019). However, research indicates that the number of E. coli growing on the wounds vary in different lettuce varieties. Therefore, it is important to determine the reasons for the differences in the antibacterial properties of lettuce and find the varieties that are suitable as fresh-cut vegetables.
During the growth of biological cells and tissues, a variety of metabolites with very low molecular weight were generated, of which 92 could be qualitatively and quantitatively analyzed by metabolomics (Li et al., 2010; Lindon et al., 2006; Sun et al., 2011). Metabolomics, a new technology, is based on a widely targeted metabolome database (Sawada et al., 2009) and adopts a multiple reaction–monitoring mode to qualitatively and quantitatively detect metabolites in samples (Chen et al., 2013). In recent years, metabolomic analysis based on LC–MS technology has been widely used in species identification and nutritional science (Gauthier et al., 2015; Steinmeyer et al., 2015). Liquid chromatography (LC) is predominantly employed to segregate the target and the matrix components, thereby facilitating the qualitative or quantitative detection of trace components within a sample. Xue et al. (2022) used 50 kinds of pollutants from drugs and personal care products as unknown and unexpected contaminants, and analyzed the contaminated lettuce and corn substrates using UHPLC-MS. Various parameters including S-plot, permutation test, and VIP in OPLS-DA were used for screening and identification of marker compounds, which proved the effectiveness of the metabolomics method for nontargeted screening of various unknown and unexpected drugs and personal care products in plant-derived foods. Yang et al. (2018) combined GC×GC-TOF/MS and UPLC-IMS-QTOF/MS to conduct nontargeted metabolomics analysis of 30 lettuce cultivars. The results revealed that the metabolomics differences between different varieties of lettuce were related to secondary metabolism. van Treuren et al. (2018) determined the phytochemical metabolite profiles of 150 Lactuca samples. The research results indicated that many plant chemicals had species specificity, some of which were typically associated with traits related to plant health or nutritional value. For a large number of phytochemicals, the relative abundance was either positively or negatively correlated with the available phenotypic data on resistance to pests and diseases, indicating their potential role in plant resistance. Liu et al. (2021) used ultra-high-performance liquid chromatography–high resolution mass spectrometry (UHPLC-HRMS) to analyze the metabolites that may be related to browning in lettuce. Otify et al. (2023) used LC-QTOF-MS/MS metabolomics, assisted by molecular networking and integrated with metabolomics, to annotate 195 metabolites of six lettuce varieties, and evaluated the biological activity of lettuce against nonantibiotic and drug-resistant bacteria, contributing to lettuce breeding. Although several related studies have reported the metabolic profiles of lettuce polyphenols, they are still incomplete and lack relevant research on their antibacterial properties. This study aimed to screen lettuce varieties with high resistance to E. coli and adopt the broadly targeted metabolomics technology of UHPLC-MS. Unsupervised PCA, OPLS-DA, and other multivariate statistical analysis methods were used to identify the metabolites in different resistant lettuce varieties, determine the key factors related to their antibacterial properties, and provide a theoretical basis for the selection and comprehensive utilization of the different varieties.
Materials and Methods
Lettuce samples and chemicals
The lettuce cultivation facility is situated at the Crop Variety Test and Exhibition Base in Changping District, Beijing, positioned at a latitude of 40° North. The soil type of the facility is sandy loam, and the soil composition is as follows: 50% sandy soil, 20% clay, and 5% organic matter. The density of lettuce planting is 80 plants/bed, and the plant distance is 0.3 × 0.3 m (Zhong et al., 2009).
This study involved 10 lettuce varieties. For clear expression, the full names are abbreviated as follows: Beisheng No. 1: B1; Beisheng No. 2: B2; Beisheng No. 3: B3; Beisheng No. 4: B4; Shooter 101: SS; Beisansheng No. 1: BS1; Beisansheng No. 2: BS2; Beizisheng No. 1: BZ1; Beizisheng No. 2: BZ2; Beizisheng No. 3: BZ3. All chemicals used in this study were of LC–MS grade.
Strains
The gram-negative bacteria E. coli YS MN153456 used throughout the study was separated from the lettuce; the strains were stored in the Food Microbiology Laboratory (Beijing University of Agriculture, Beijing, China), and the bacteria were activated in nutrient broth (AOBOX, Beijing, China) at 37°C in shaker for 48 h. The prepared bacterial suspension was subcultured in Luria–Bertani (LB) broth (AOBOX, Beijing, China) at 37°C for 12 h and harvested in sterile saline water. The concentration of the bacterial suspension was monitored by its transmittance at OD600 nm using a T6 spectrophotometer (Beijing Puxi General Instrument Co., Ltd., Beijing, China).
Screening of lettuce varieties with high resistance to E. coli
Fresh lettuce juice was centrifuged at 8000 rpm for 15 min in a large, refrigerated centrifuge (Thermo Scientific), and filtered through a 0.22 μm filter to obtain 50 mL of filtrate. The filtrate was stored in a sterile conical flask at 4°C for use. The cultured E. coli YS was diluted to 1 × 102 colony-forming units (CFU)/mL, inoculated into the lettuce juice of 2% inoculum, and cultured at 37 ± 1°C for 48 h. The E. coli YS was then counted in five parallel groups for each sample.
Lettuce metabolomics
Extraction of metabolites
According to a slightly modified method of Doppler et al. (2016) and Chen et al. (2013), 2 g of fresh lettuce leaves was taken, wrapped in tin foil, marked, and quickly put into liquid nitrogen for 15 min and subjected to vacuum freeze-drying for 24 h. The freeze-dried samples were then crushed with a mixer mill for 30 s at 45 Hz. A 50 mg aliquot of each sample was precisely weighed and transferred to an Eppendorf tube, after which 800 μL of extract solution was added (methanol/water 3:1, precooled at −40°C, containing the internal standard). The samples were vortexed for 30 s, homogenized at 35 Hz for 4 min, and sonicated for 5 min in an ice water bath. The homogenization and sonication processes were repeated thrice. The samples were then extracted overnight at 4°C on a shaker and centrifuged at 12,000 rpm for 15 min at 4°C. The supernatant was carefully filtered through a 0.22 μm microporous membrane. 40 μL was taken from each sample and pooled as QC samples, which were stored at −80°C until the UHPLC-MS analysis. Four samples per group were tested in parallel.
UHPLC-MS analysis
Chromatographic conditions: The UHPLC separation was carried out in ExionLCTM System (Sciex) with a Waters Acquity UPLC HSS T3 column (2.1 × 100 mm, 1.8 μm). Mobile phase A and B were 0.1% formic acid in water and acetonitrile, respectively. The column and the autosampler temperatures were set at 40°C and 4°C, respectively, and the injection volume was 2 μL.
In the initial state, the mobile phase A and B were 98% formic acid aqueous solution and 2% acetonitrile, respectively. A flow rate of 400μL/min was maintained throughout the operation. Within 0.5–10 min after initiation, the aqueous solution of formic acid in mobile phase A decreased by 50%, while acetonitrile in mobile phase B increased by 50%. Subsequently, within the first 11–13 min, the aqueous solution of formic acid in mobile phase A reached 5%, while the acetonitrile in mobile phase B reached 98% and remained stable. Within the first 13–15 min, the aqueous solution of formic acid in mobile phase A recovered to 98%, and the acetonitrile in mobile phase B recovered to 2%, and eventually remained stable.
Mass spectrometry conditions: Sciex QTrap 6500 + instrument (Sciex Technologies) was used for data analysis in multireaction monitoring mode. Typical ion source parameters were used: ion spray voltage +5500/−4500 V; curtain gas 35 psi; temperature 400°C; ion source gas 1: 60 psi; ion source gas 2: 60 psi; DP ±100 V.
Antibacterial susceptibility tests
Antibacterial tests were performed using the Oxford cup method (Zhu et al., 2019). The extract was redissolved in ultrapure water to obtain a final concentration of 200 mg/mL. 100 μL of the prepared 1 × 106 CFU/mL E. coli YS strain was added to 100 mL of nutrient agar medium (AOBOX, Beijing, China) at approximately 50°C and shaken evenly, and 10 mL of the mixture was poured into a sterile petri dish. After solidification, 200 μL of the prepared extract solution was implanted on the inoculated agar through an Oxford cup. The DIZ was measured after 24 h of incubation at 37°C.
Effect of lettuce polyphenol extracts on the morphological structure of E. coli
Polyphenol extracts from lettuce were procured utilizing ultrasonic-assisted extraction techniques (Zhidi et al., 2022). E. coli was cultured in LB broth at 37°C for 8 h and washed off with PBS buffer to prepare E. coli suspension with a concentration of 106 CFU/mL. The polyphenol extract was added for 6 h, and the suspension was centrifuged. The precipitated cells of E. coli were washed twice with 0.1 M PBS (pH 7.4) and fixed overnight at 4°C with 2.5% (v/v) glutaraldehyde in 0.1 M PBS. The cells were then dehydrated by sequential exposure to ethanol with concentrations ranging from 30 to 100%, and the ethanol was finally replaced with tert-butanol. The centrifuged cells were dried at a “critical point” in liquid CO2 at 95 bar pressure, and the samples were covered with gold by cathodic spray. Finally, the morphology of the bacterial cells was observed on a scanning electron microscope (Prysese Instrument).
Determination of the isoquercitrin content
The isoquercitrin content in the extracts was measured using HPLC (Agilent, Santa Clara, CA). The HPLC conditions were as follows: ZORBAX SB C18 column (250 × 4.6 mm, 5 µm); mobile phase A: 0.1% formic acid; mobile phase B: methanol; gradient elution: ~0–0.5 min, 11% B; ~0.5–5 min, ~11–25% B; ~5–6 min, ~25–50% B; ~6–20 min, 50% B; ~20–30 min, ~50–11% B; flow rate: 0.5 mL/min; injection volume: 5 μL; and column temperature: 30°C. Chromatographic data were scanned in the 360 nm wavelength range. Purified lettuce polyphenols were dissolved in methanol at a concentration of 1 mg/mL. The reference substance of isoquercitrin was accurately weighed and placed in a brown vial, fully dissolved in methanol. The reference substance reserve solutions were prepared at concentrations 0.01, 0.05, 0.1, 0.15, and 0.2 mg/mL, and the peak area was detected by HPLC. The standard curve was drawn with peak area as ordinate and concentration as abscissa. Isoflavone glycoside standard curve: y = 2E+07x-39366, R2 = 0.9993 is a good linear relationship.
Data analysis
SCIEX Analyst Work Station software (version 1.6.3) was used for multiple reaction–monitoring data acquisition and processing. The raw MS data (.wiff) files were converted to TXT format using an MS converter. An in-house R program and database were applied for peak detection and annotation. SIMCA software (version 16.0.2, Sartorius Stedim Data Analytics AB, Umea, Sweden) was used to perform a multivariate statistical analysis on the UHPLC-MS data, including unsupervised PCA and OPLS-DA.
The PCA was mainly used to study the sample distribution, deviation characteristics, and common trends. The OPLS-DA was used to classify samples, identify the most discriminant variables, and verify them based on the goodness of fit (R2Y) and goodness of prediction (Q2Y) for their classification and prediction ability. The model was tested for overfitting by permutation test (n = 200) and the negative value of the intercept (Q2 intercept) was obtained, indicating the robustness of the model. The VIP score showed the contribution of each variable to the model. The VIP scores in the predicted components were analyzed, and only those metabolites with a VIP score of ≥1 were considered categories between which there was discrimination. Finally, the metabolite information obtained was screened according to the VIP value, the P-value of Student’s t-test, and the FC, with the log value of 2 as the base, so as to obtain the metabolites with significant differences.
Statistical analysis
All assays were analyzed in quadruplicate. A one-way analysis of variance was performed on the number of E. coli colonies, the DIZ, and the content of isoquercitrin. Significance of P ≤ 0.05 and P ≤ 0.01 levels was determined using SPSS 16.0 software.
Results
Screening of antibacterial lettuce varieties
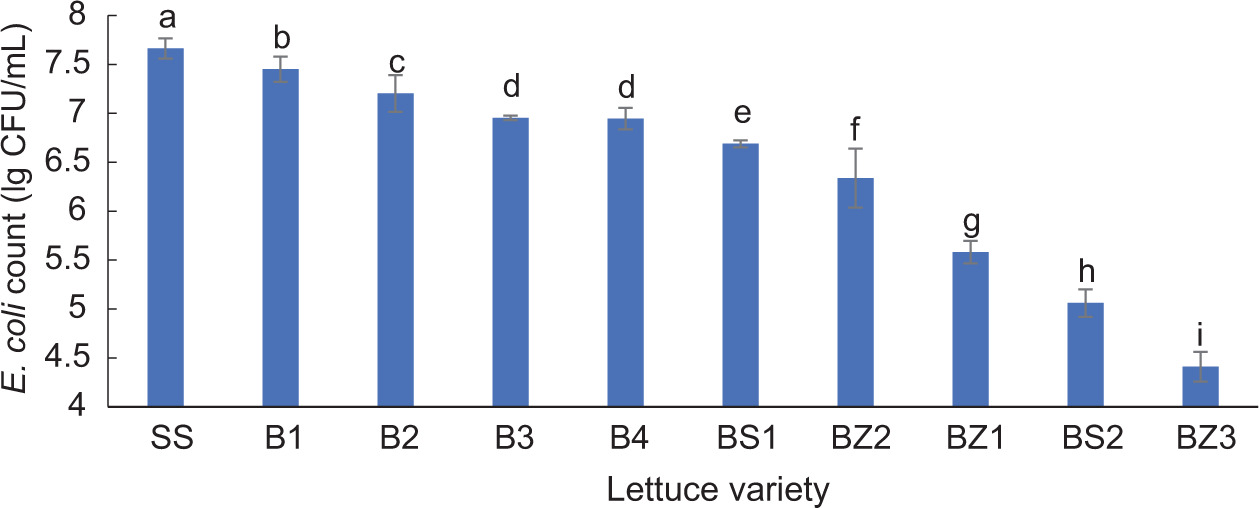
After E. coli YS inoculation, the counts for different varieties of lettuce juice varied significantly. Figure 1 shows the results of E. coli YS growth in juice of different lettuce varieties. The number of E. coli colonies in SS was the highest, at 7.66 lg CFU/mL, and that in BZ3 was the lowest, with only 4.41 lg CFU/mL. The results showed that BZ3 had the highest inhibitory effect on E. coli and the strongest antibacterial activity, whereas SS had the poorest performance for both. At the same time, BS1, which ranked in the middle, was selected as the control for the metabolomic analysis.
Figure 1. Escherichia coli count results from different varieties of lettuce juice. Different letters in the figure indicate significant differences (P < 0.05).
Metabolomic analysis of lettuce
Analysis of the QC samples
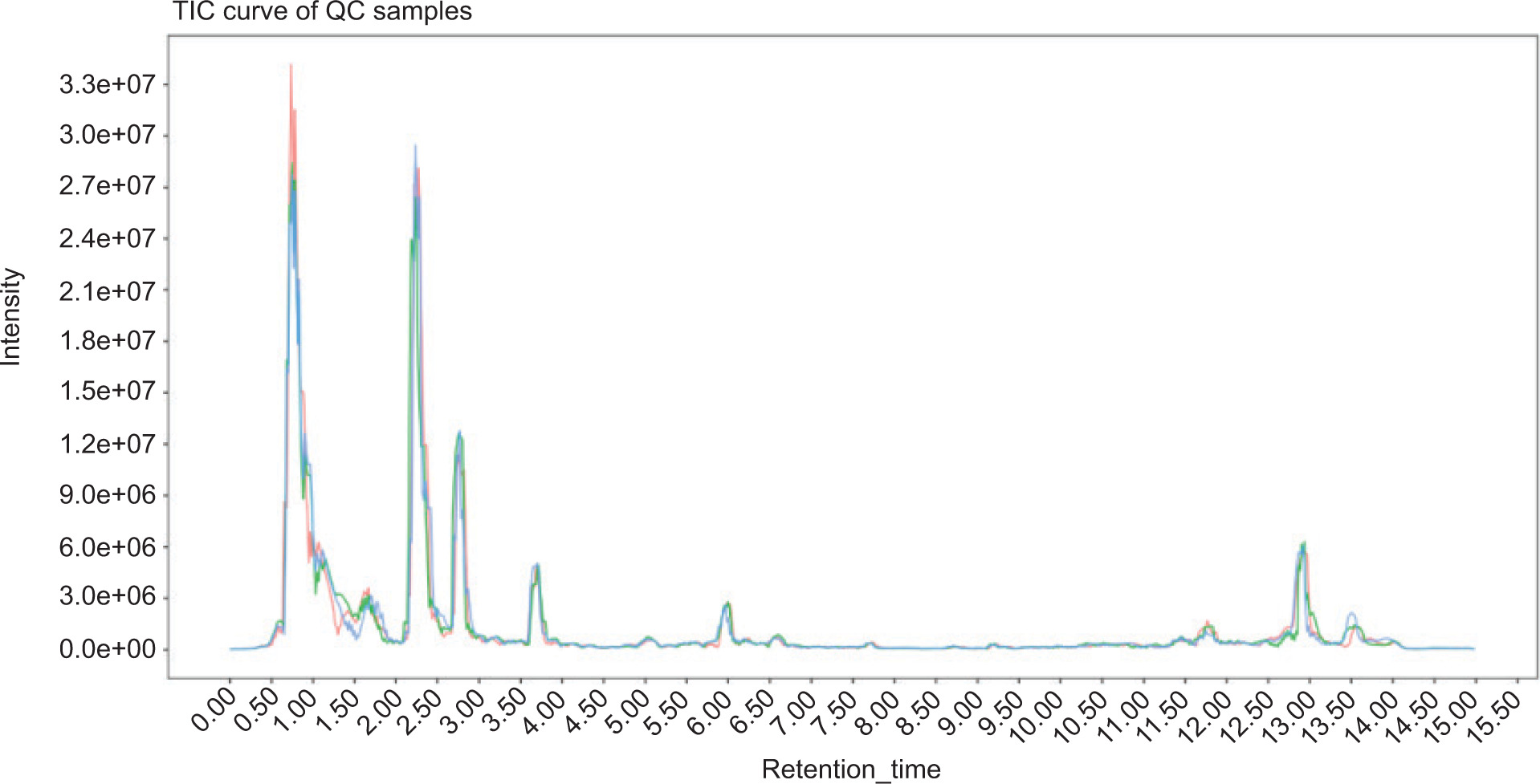
Quality control samples were used to evaluate the stability and repeatability of the system during the test. As illustrated in Figure 2, results showed that the curves of the total ion current for metabolite detection had a high overlap, that is, the retention time and peak intensity were consistent, indicating that the signal stability was better when the mass spectrometer detected the same sample at different times.
Figure 2. Overlay of the TIC of quality control (QC) samples by mass spectrometry detection.
Multivariate statistical analysis of metabolites in lettuce
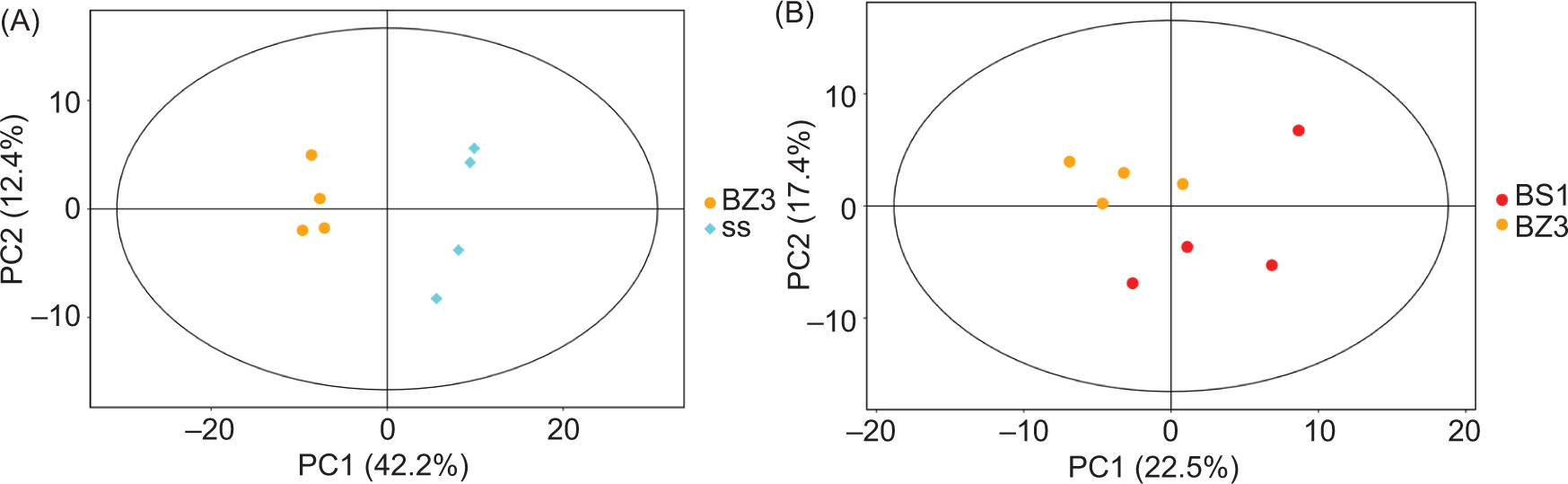
To analyze the difference in metabolites between the two groups of lettuce samples, PCA analysis and OPLS-DA were used. The PCA score between the two groups is shown in Figure 3, and the OPLS-DA analysis and its model replacement test are summarized in Figure 4.
Figure 3. Principal components analysis for groups. Group A is BZ3 versus SS; group B is BZ3 versus BS1.
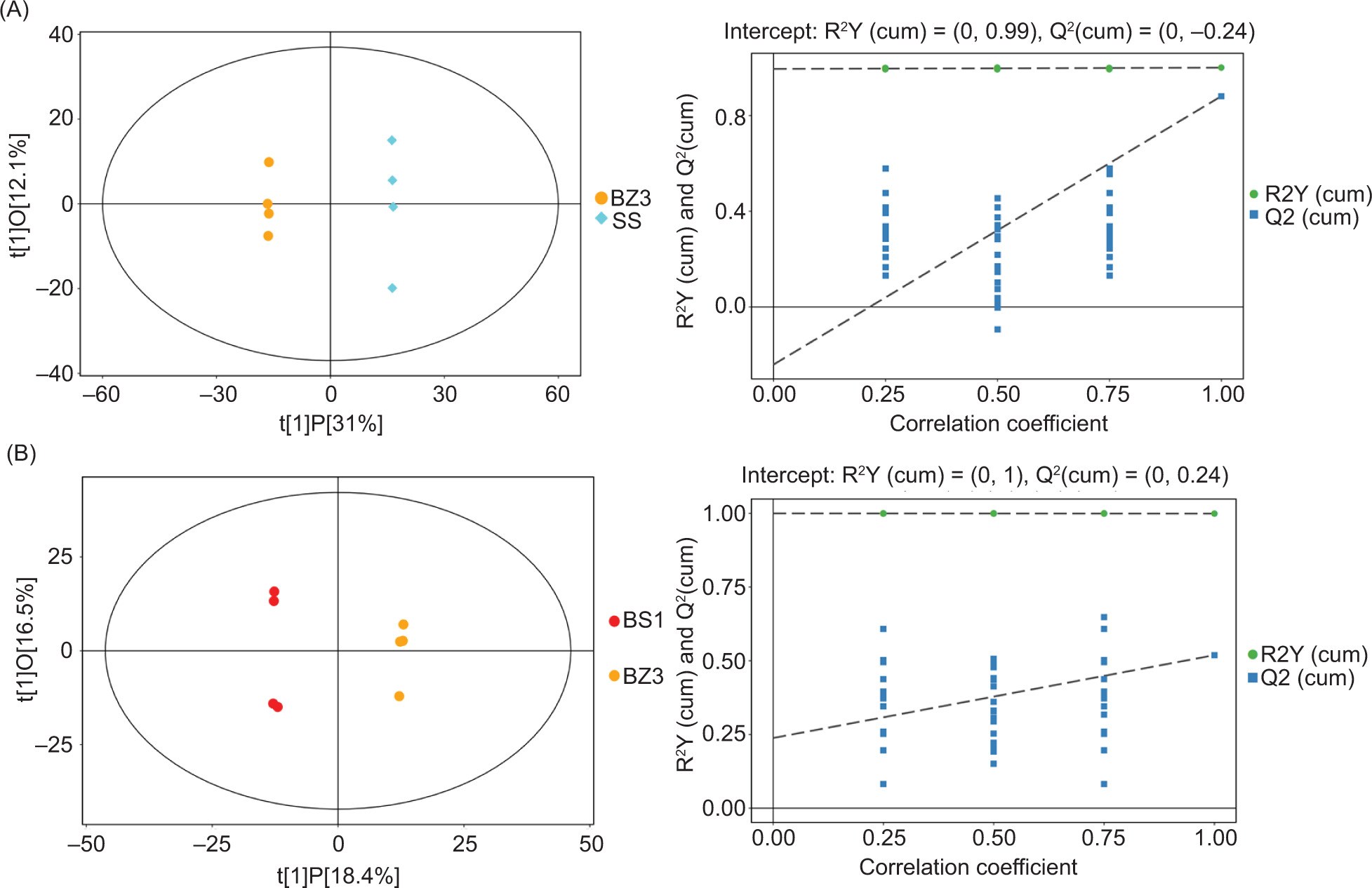
Figure 4. Orthogonal partial least squares discriminant analysis for groups. Groups A and A' are BZ3 versus SS; groups B and B' are BZ3 versus BS1.
The PCA modeling method was used for the analysis because the antibacterial activities of BZ3 and SS were very different: when principal component (PC) 1 was 42.2%, PC2 was 12.4%, and BZ3 and SS could clearly be separated as having differences in metabolites. However, due to the small differences in antibacterial activity of BZ3 and BS1 with those of BZ3 and SS, the differences in metabolites of the former duo were smaller.
For the OPLS-DA model diagrams, it was often necessary to use permutation tests to verify the effectiveness of the model to prevent the overfitting phenomenon, which would lead to deviations. R2Y represents the interpretation level of the model in the Y-axis direction, while Q2 indicates its predictive ability (Xue et al., 2022). The closer R2Y (cum) and Q2 are to 1, the more stable and reliable the model is. Q2 > 0.5 indicates better predictive ability of the model. In Figures 4A and B, the R2Y (cumulative) and Q2 values of BZ3 versus SS were 1 and 0.881, respectively, and the R2Y (cumulative) and Q2 values of BZ3 versus BS1 were 0.999 and 0.519, respectively, which demonstrated that the two OPLS-DA models were effective and there was no overfitting. Therefore, it could accurately analyze the differences in metabolites between the different antibacterial lettuce varieties. As shown in Figures 4A and B, the two different varieties of lettuce could clearly be distinguished in the score chart, which demonstrated that intermediate differences were present in the types and contents of metabolites.
Screening of differential metabolites in lettuce
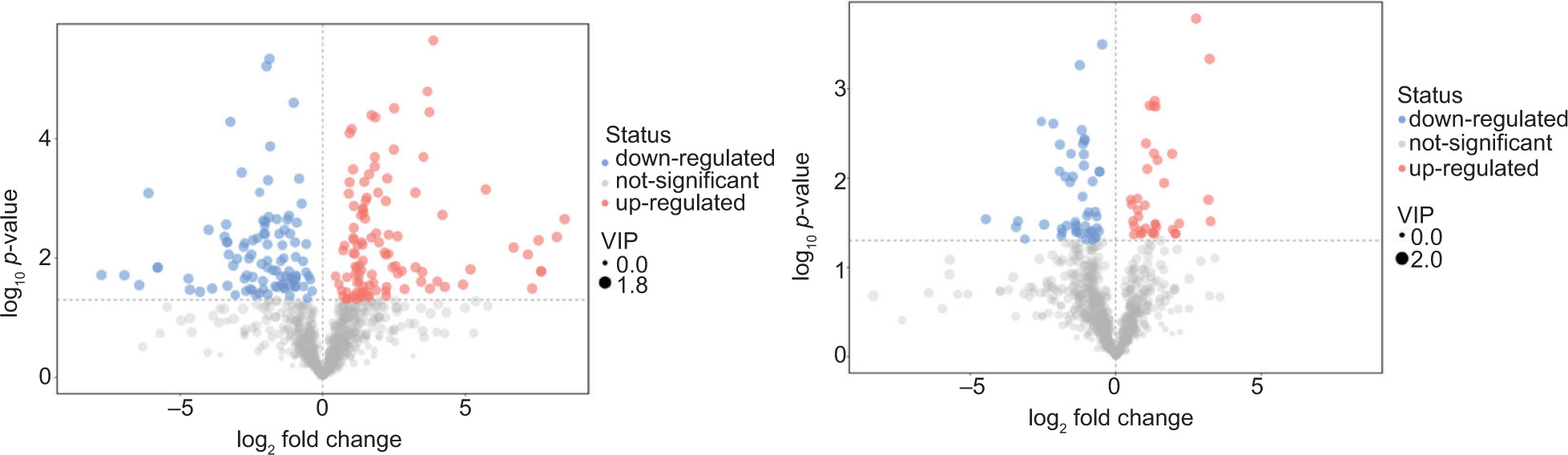
The different metabolites between the groups were screened according to the VIP value of PC1 generated by OPLS-DA and the P-value of the t-test, (VIP ≥ 1 and P < 0.05). The higher the VIP value, the more it indicated the strength of the influence of differential metabolites on the antibacterial activity of different varieties of lettuce. As shown in Figure 5, 204 differential metabolites were identified in BZ3 and SS, 106 of which were upregulated metabolites, including 16 alkaloids, 11 terpenes, 6 phenols, 22 flavonoids, and 3 flavonoids. A total of 98 metabolites were downregulated, including 12 alkaloids, 7 phenols, and 21 amino acids and derivatives. Eighty-six differential metabolites were identified in BZ3 and BS1, of which 36 were upregulated and 50 were downregulated.
Figure 5. Volcano plot of different metabolite screening between groups. Group A is BZ3 versus SS; group B is BZ3 versus BS1.
Cluster analysis of differential metabolites
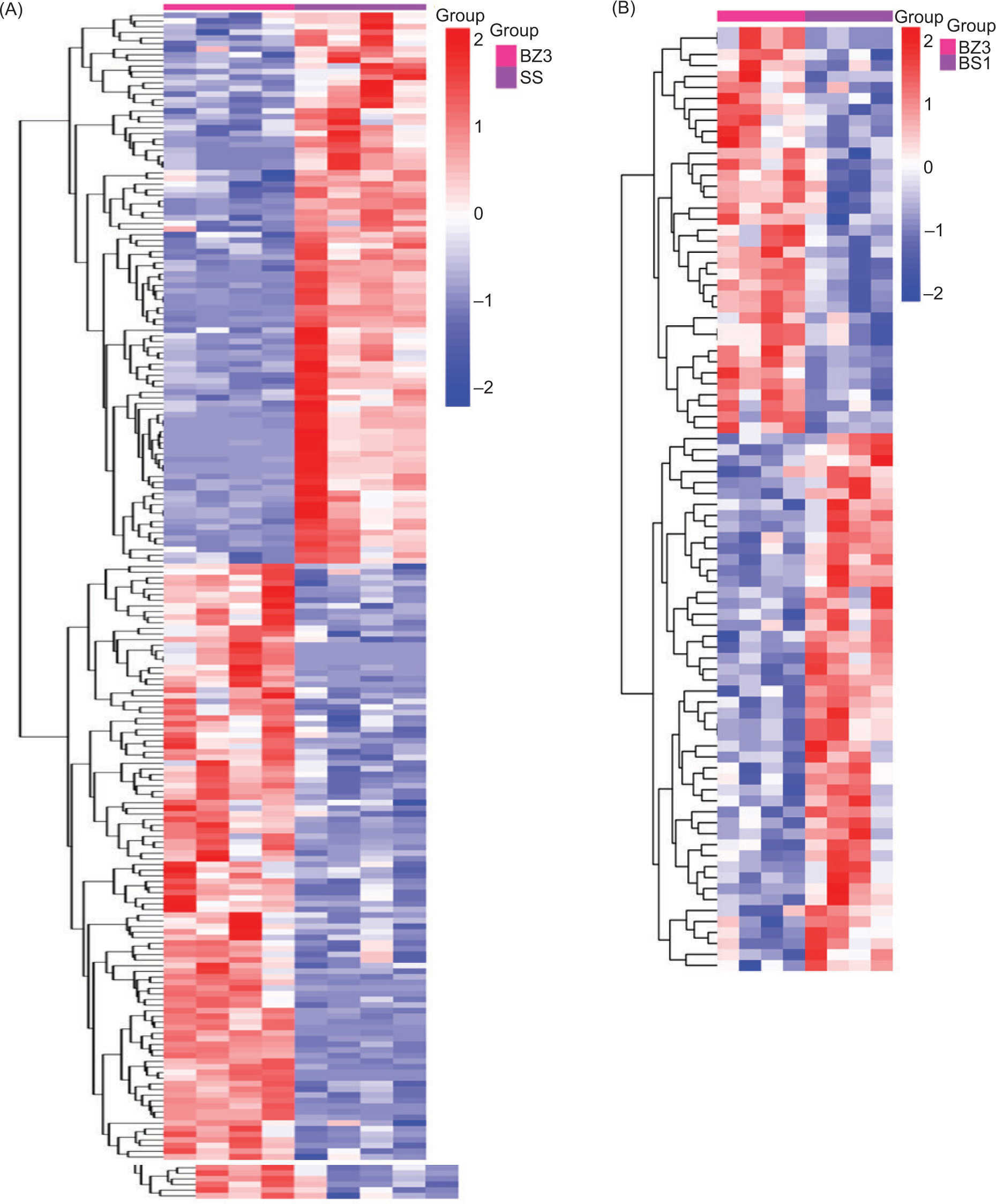
A heatmap of the hierarchical clustering analysis method was used to analyze the differential metabolites in BZ3 versus SS and BZ3 versus BS1. The results are shown in Figure 6. A significant difference was found in the amounts of metabolites between the two groups, with the difference between the BZ3 and SS groups being more pronounced. The higher contents of BZ3 included cynaroside, delphinidin-3-O-glucoside, isoquercitrin, and glycitein methyl hesperidin, while adenosine 2',3'-cyclic phosphate, guanosine 3',5'-cyclic monophosphate, and (−)-cinchonidine were found in higher amounts in BS1. In contrast, l-glutamine, d-glutamine, and l-asparagine were higher in SS.
Figure 6. Heatmap of the hierarchical clustering analysis for differential metabolites. Group A is BZ3 versus SS; group B is BZ3 versus BS1. The color indicates the content: red indicates high expression, and blue indicates low expression.
Analysis of the FC in the metabolite difference
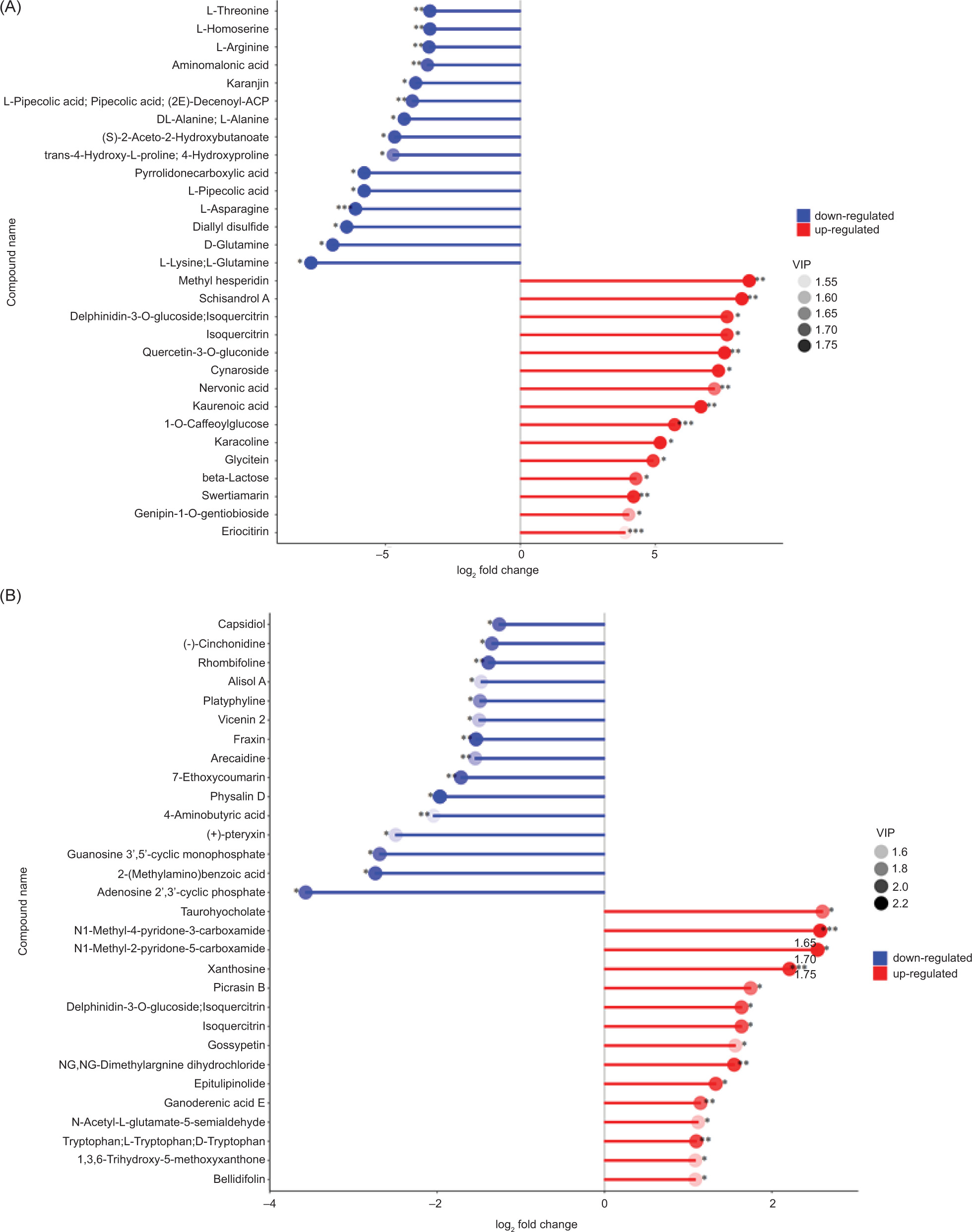
By calculating the corresponding proportions of quantitative values for different metabolites, their significant changes are identified. To show the different metabolites with a large degree of change, we calculated the corresponding ratio of the quantitative value of the metabolites, expressed as FC, and took the logarithmic conversion with base as 2. The top 15 differential metabolites according to their FC values are shown in Table S1. Figure 7 is a matchstick diagram of the two groups with significantly different metabolites. Because the metabolite contents in samples BZ3 and SS were quite different, the FC value was relatively large. Among the contents were 10 metabolites with log2FC ≥ 4, whereas in BZ3 and BS1, the FC value was small and the log2FC value was less than 4. Delphinidin-3-O-glucoside and isoquercitrin were the upregulated differential metabolites shared by the two groups, indicating that the contents of both metabolites in BZ3 were higher than those in BS1 and SS.
Figure 7. Matchstick analysis for significantly different metabolites. Group A is BZ3 versus SS; group B is BZ3 versus BS1. The abscissa shows the multiples of change after logarithmic transformation, and the shades of the dots represent the variable importance in projection (VIP) value. An asterisk (*) represents significance: *0.01 < P < 0.05, **0.001 < P < 0.01, ***P < 0.001.
Analysis of the KEGG metabolic pathway
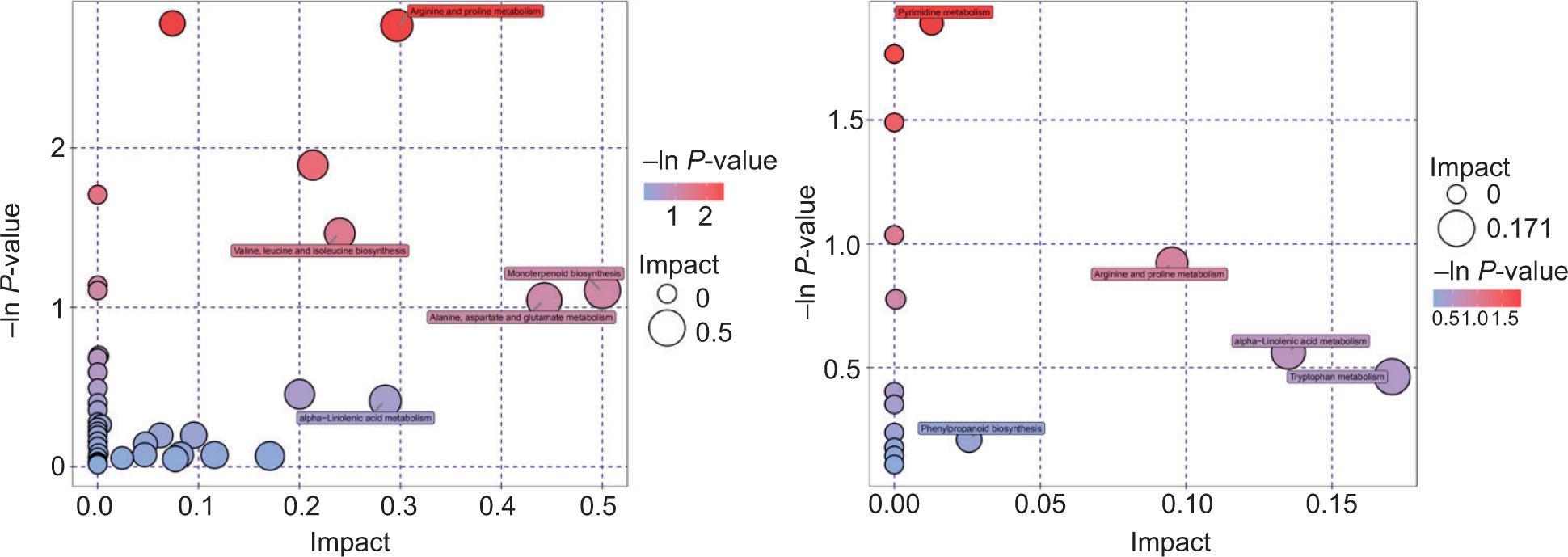
The screened-out differential metabolites were analyzed for metabolic pathway enrichment according to the KEGG data platform, and the results of the metabolic pathway analysis are displayed in a bubble chart (Figure 8). Forty-one metabolic pathways were enriched by BZ3 and SS. Among the metabolic pathways that were significantly enriched were monoterpenoid biosynthesis; alanine, aspartate and glutamate metabolism; valine, leucine, and isoleucine biosynthesis; α-linolenic acid metabolism; and arginine and proline metabolism. In contrast, 15 metabolic pathways were enriched by BZ3 and BS1. The pathways that were significantly enriched were tryptophan metabolism, α-linolenic acid metabolism, arginine and proline metabolism, phenylpropanoid biosynthesis, and pyrimidine metabolism.
Figure 8. Pathway analysis for the two groups. Group A is BZ3 versus SS; group B is BZ3 versus BS1. Each bubble in the bubble chart represents a metabolic pathway. The abscissa of the bubble and the size of the bubble indicate the size of the enrichment and size of the influence factor of the pathway in the topology analysis. The larger the value, the greater the impact factor. The ordinate of the bubble and the bubble color indicate the P-value of the enrichment analysis (by taking the negative natural logarithm, i.e., −ln(p)). The darker the color, the smaller the P-value and the more significant the degree of enrichment.
Bacteriostatic activity
The polyphenol extracts of each variety had inhibitory effects on E. coli, and significant differences were found in their antibacterial effects (P < 0.05). The inhibition zone diameters of BZ3, BS1, and SS polyphenol extracts against E. coli were 18.61,16.33, and 11.23 mm, respectively. The inhibitory ability of BZ3 polyphenol extract to E. coli was significantly higher than that of other varieties.
Effect of lettuce polyphenol extracts on the morphological structure of E. coli
The micromorphology of E. coli in the control group and the lettuce polyphenol extract treatment group is shown in Figure 9. The morphology of E. coli in the control group (A) showed a complete and smooth rod-like structure. Upon introduction of lettuce polyphenol extract (B), there was a reduction of approximately 25% in the average bacterial length. Furthermore, the bacterial surface exhibited a rough and incomplete morphology, while the cellular surface was markedly depressed and wrinkled. The overall morphology of the bacteria was compromised, leading to cell disruptions and dissolution of their contents.
Figure 9. The effect of polyphenol extractson the morphological structure of E. coli. The cells indicated by the red arrow are those with folded morphology.
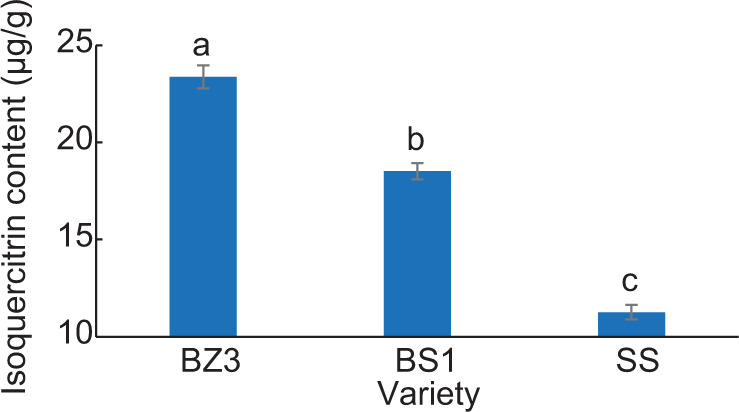
Figure 10. Contents of isoquercitrin in different varieties of lettuce. Different letters in the figure indicate significant differences (P < 0.05).
Content of isoquercitrin in the extract
The isoquercetin content in different varieties of lettuce was determined. There was a significant difference in its content in BZ3, BS1, and SS (P < 0.05), with BZ3 having the highest content, at 23.38 μg/g, and SS the lowest (11.26 μg/g). The content of BS1 fell in between the two (18.52 μg/g).
Correlation analysis
A correlation analysis of each index was carried out, and the results are shown in Table 1. The isoquercitrin content in lettuce samples positively correlated with the antibacterial activity of polyphenol extracts (r = 0.995) and negatively correlated with the growth of E. coli in lettuce juice (r = −1, P < 0.01). The results showed that the higher the isoquercitrin content, the stronger the antibacterial activity of the lettuce polyphenol extract, the less E. coli grew in the lettuce juice, and the stronger the lettuce antibacterial activity. Therefore, the isoquercitrin content was closely related to the antibacterial activity of the lettuce.
Table 1. Correlation analysis of each index.
| Isoquercitrin content | Bacteriostatic activity | E. coli counts in lettuce juice | |
|---|---|---|---|
| Isoquercitrin content | 1 | ||
| Bacteriostatic activity | 0.995 | 1 | |
| E. colicounts in lettuce juice | −1** | −0.993 | 1 |
**Indicates significant difference, P < 0.01.
Discussion
Lettuce is rich in nutrients and has a crisp texture, which is greatly enjoyed by consumers in daily life. As a common fresh vegetable, its microbial safety has also attracted much attention. During the growth process of lettuce, metabolites are abundant, and there are interspecies differences as well. The types and contents of the metabolites are closely related to their antibacterial activities. Broadly targeted metabolomics has been widely used in profiling plant metabolites in recent years.
During the experiment, we found that the growth of E. coli YS differed when the bacterium was inoculated into lettuce juice samples. Therefore, a broadly targeted metabolome of lettuce metabolites was used to analyze their differences in different lettuce varieties. The PCA results of unsupervised pattern recognition showed significant differences in the metabolite content of different lettuce varieties. Up on comparison between BZ3 and SS, 204 differential metabolites were detected, and 86 differential metabolites were detected and between BZ3 and BS1. The results showed that metabolites differed across different varieties of lettuce, which were consistent with the findings by van Treuren et al. (2018). In addition, the study by Viacava et al. (2017) has shown that purple leaf lettuce has more phenolic compounds than the green leaf variety. Consistent with their experimental results, in our research, BZ3, a purple leaf lettuce, was richer in phenolic compounds than BS1 and SS. For example, in BZ3 and BS1, the FC value of isoquercitrin content was 3.11, while in SS, it was 201.65.
In the comparative analysis of BZ3 and SS, 106 upregulated differential metabolites were detected, most of which were flavonoids, alkaloids, and terpenes. Among them, 11 had log2FC > 4. In addition, 98 downregulated differential metabolites were detected. The content of antibacterial active substances in BZ3 was significantly higher than that in SS; thus, the antibacterial activity of the former was stronger than that of latter. The difference in antibacterial activity between BZ3 and BS1 was small, thus the number of different metabolites between them was significantly less than that between BZ3 and SS. Assefa et al. (2021) analyzed the metabolites of 113 red lettuce samples and found that the content of metabolites in them varied greatly. Cyanidin 3-O-(6″-O-malonyl) glucoside, 2,3-di-O-caffeoyltartaric acid, and quercetin 3-O-(6″-O-malonyl) glucoside predominated in red lettuce samples, and high levels of anthocyanins, flavonoids, and hydroxycinnamoyl derivatives were detected at the leaf incisions. The comparative analysis of BZ3 versus SS and BZ3 versus BS1 revealed delphinidin-3-O-glucoside and isoquercitrin as the significantly upregulated metabolites in the two groups. The isoquercitrin content of BZ3 was significantly higher than that of SS and BS1, and it had the greatest antibacterial effect. The results of correlation analysis showed that isoquercitrin content positively correlated with the antibacterial activity of lettuce, a key factor in the resistance of lettuce against E. coli. Zhang et al. (2024) extracted isoquercitrin from the antibacterial component of the fruit of Amomum villosum Lour. According to Yun et al. (2018), isoquercitrin is a flavonoid with good antibacterial activity. Tang et al. (2023) extracted isoquercitrin from the key antibacterial ingredient of an edible herb Potentilla kleiniana Wight et Arn. Fei et al. (2023) analyzed the metabolites of 50 kinds of lettuce by UPLC-Q-ToF-MS. The study revealed a positive correlation between the total phenols and total flavonoids content in lettuce and its antioxidant capacity. Furthermore, it was established that quercetin serves as a characteristic index for assessing the antioxidant activity of lettuce. In this study, only the resistance levels of 23 lettuce varieties to E. coli were compared and analyzed. However, it is imperative to broaden the scope of this research to encompass a more comprehensive analysis of the variability and correlation of lettuce metabolites. The analysis of correlational data pertaining to the characteristics of various lettuce varieties can offer valuable theoretical insights for the selection and breeding processes.
This study explored the impact of lettuce polyphenol extract on the cellular structure of E. coli. This study has demonstrated that the inhibition of polyphenol extracts on E. coli is achieved through the disruption of cellular structure, resulting in cell shrinkage and rupture. This process serves to inhibit the growth and reproduction of bacteria. Similar observation was reported by Jeyakumar and Lawrence (2021) with changes in the surface morphology of cells when treated with eugenol. It has been reported that flavonoids (Xu et al., 2019) and phenols (Bonechi et al., 2018) are secondary metabolites with antibacterial activity. Yu (2021) found that mulberry leaf polyphenols could significantly inhibit the motility of E. coli and the production of extracellular polymers. Birru et al. (2021) found that apple polyphenols can inhibit the growth of pathogenic bacteria and relieve the symptoms of chronic obstructive pulmonary disease. Taleb et al. (2016) showed that jujube pulp polyphenols inhibited the growth of E. coli and S. aureus by inducing oxidative stress in bacteria to produce hydrogen peroxide.
In the present study, lettuce polyphenol extracts had antibacterial effects on E. coli, and isoquercitrin content positively correlated with the antibacterial effect of the extract. The higher the content of isoquercitrin, the greater the antibacterial effect, which is consistent with the conclusion of bamboo leaf flavonoids inhibiting E. coli (Xiao et al., 2023). This finding will facilitate further investigation into the comprehensive utilization of lettuce. Moreover, lettuce polyphenols have good antioxidant activity (Xiao et al., 2015). Therefore, the selection of lettuce varieties with high polyphenol content as food raw materials can delay cell senescence and reduce the risks of cardiovascular disease and cell carcinogenesis to a certain extent (Hehuan et al., 2022). Consequently, the selection of lettuce varieties with elevated isoquercitrin content, such as BZ3, as raw materials in vegetable foods, such as salads and sandwiches, will significantly improve food safety.
Conclusion
Lettuce is a popular raw vegetable. Significant differences exist in the antimicrobial activity of and the contents and types of metabolites in different varieties of lettuce (P < 0.05). Purple leaf lettuce and loose-leaf lettuce have more metabolites than that of the green leaf variety and head lettuce, respectively. The greater the difference in the antibacterial activity of the lettuce, the greater the difference in the isoquercitrin content. The E. coli growth negatively correlated with the isoquercitrin content. The higher the isoquercitrin content, the stronger the antibacterial activity of the lettuce polyphenol extract, and the better the antibacterial activity of the lettuce.
Ethics Approval and Consent to Participate
Not applicable.
Consent for publication
Not applicable.
Competing Interests
The authors declare that there are no competing interests.
Funding
This work was supported by Urban Agriculture and Forestry Interdisciplinary Platform Construction Project of Beijing Agricultural University and 2023 Degree and graduate Education Reform and development project of Beijing University of Agriculture. HLB was supported by the USDA NIFA funding.
Availability of Data and Materials
The data sets used and analyzed during the present study are available from the corresponding author on reasonable request.
Author Contributions
JL and ZC carried out the experiments and drafted a manuscript. FM, DL, and XC participated in its design and coordination. HL-B and YZ improved the writing of the manuscript. XG designed the experimental scheme and carried out the overall planning and improvement of the manuscript. All authors have read and approved the final manuscript.
Acknowledgments
Technical support from Shanghai Biotree Biotech Co., Ltd. on the metabolomics test is appreciated.
AI declaration statement
The author solemnly declares that the content of this article is original and has not been assisted by any artificial intelligence AI tools, including but not limited to ChatGPT, nor has it used software or online services such as automatically generated text.
REFERENCES
Assefa, A.D., Hur, O., Hahn, B., Kim, B., Ro, N.Y., and Rhee, J.H. 2021. Nutritional metabolites of red pigmented lettuce (Lactuca sativa) germplasm and correlations with selected phenotypic characters. Foods 10(10): 2504. 10.3390/foods10102504
Birru, R.L., Bein, K., Bondarchuk, N., Wells, H., Lin, Q., Di, Y.P., et al. 2021. Antimicrobial and anti-inflammatory activity of apple polyphenol phloretin on respiratory pathogens associated with chronic obstructive pulmonary disease. Frontiers in Cellular and Infection Microbiology 11: 652944. 10.3389/fcimb.2021.652944
Bonechi, C., Donati, A., Tamasi, G., Leone, G., Consumi, M., Rossi, C., et al. 2018. Protective effect of quercetin and rutin encapsulated liposomes on induced oxidative stress. Biophysical Chemistry 233: 55–63. 10.1016/j.bpc.2017.11.003
Chen, W., Gong, L., Guo, Z., Wang, W., Zhang, H., Liu, X., et al. 2013. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Molecular Plant 6: 1769–1780. 10.1093/mp/sst080
Cheng, D.M., Pogrebnyak, N., Kuhn, P., Krueger, C.G., Johnson, W.D., and Raskin, I. 2014a. Development and phytochemical characterization of high polyphenol red lettuce with anti-diabetic properties. PLoS One 9(3): e91571. 10.1371/journal.pone.0091571
Cheng, D.M., Pogrebnyak, N., Kuhn, P., Poulev, A., Waterman, C., Rojas-Silva, P., et al. 2014b. Polyphenol-rich Rutgers Scarlet lettuce improves glucose metabolism and liver lipid accumulation in diet-induced obese C57BL/6 mice. Nutrition 30(7–8): S52–S58. 10.1016/j.nut.2014.02.022
Cuggino, S.G., Possas, A., Posada-Izquierdo, G.D., Theumer, M.G., and Pérez-Rodríguez, F. 2023. Unveiling fresh-cut lettuce processing in argentine industries: Evaluating salmonella levels using predictive microbiology models. Foods 12(21): 3999. 10.3390/foods12213999
Cui, H., Bai, M., Sun, Y., Abdel-Samie, M.A.S., and Lin, L. 2018. Antibacterial activity and mechanism of Chuzhou chrysanthemum essential oil. Journal of Functional Foods 48: 159–166. 10.1016/j.jff.2018.07.021
Dannehl, D., Becker, C., Suhl, J., Josuttis, M., and Schmidt, U. 2016. Reuse of organomineral substrate waste from hydroponic systems as fertilizer in open-field production increases yields, flavonoid glycosides, and caffeic acid derivatives of red oak leaf lettuce (Lactuca sativa L.) much more than synthetic fertilizer. Journal of Agricultural and Food Chemistry 64(38): 7068–7075. 10.1021/acs.jafc.6b02328
Doppler, M., Kluger, B., Bueschl, C., Schneider, C., Krska, R., Delcambre, S., et al. 2016. Stable isotope-assisted evaluation of different extraction solvents for untargeted metabolomics of plants. International Journal of Molecular Sciences 17(7): 1017. 10.3390/ijms17071017
El Moussaoui, A., Jawhari, F.Z., Almehdi, A.M., Elmsellem, H., Fikri Benbrahim, K., Bousta, D., et al. 2019. Antibacterial, antifungal and antioxidant activity of total polyphenols of Withania frutescens L. Bioorganic Chemistry 93: 103337. 10.1016/j.bioorg.2019.103337
Fei, Y., Xiaoyan, Z., Changyan, Z., Shiwei, W., Hengchao, E., and Xiaobei, L. 2023. Comparison and antioxidant activity of polyphenols and flavonoids in different varieties of lettuce. Food Science 13: 17–25.
Gauthier, L., Atanasova-Penichon, V., Chereau, S., and Richard-Forget, F. 2015. Metabolomics to decipher the chemical defense of cereals against Fusarium graminearum and deoxynivalenol accumulation. International Journal of Molecular Sciences 16(10): 24839–24872. 10.3390/ijms161024839
Hehuan, P., Meifeng, Z., Yangchen, Q., Yangchen, Q., and Qi, L. 2022. Research progress on pharmacological effects of polyphenolic compounds. China Pharmaceuticals. 23: 124–127.
Islam, M.Z., Lee, Y.T., Mele, M.A., Choi, I.L., Jang, D.C., Ko, Y.W., et al. 2019. Effect of modified atmosphere packaging on quality and shelf life of baby leaf lettuce. Quality Assurance and Safety of Crops & Foods 11(8): 749–756. 10.3920/QAS2019.1626
Jeyakumar, G.E., and Lawrence, R. 2021. Mechanisms of bactericidal action of eugenol against Escherichia coli. Journal of Herbal Medicine 26: 100406. 10.1016/j.hermed.2020.100406
Ketut Srie, M.J., and Purnawati, A. 2023. Phytochemical compounds and antibacterial activity to Escherichia coli of green macro algae. IOP Conference Series. Earth and Environmental Science 1131(1): 12012. 10.1088/1755-1315/1131/1/012012
Li, H., Sun, L., Mittapalli, O., Muir, W.M., Xie, J., Wu, J., et al. 2010. Bowman-birk inhibitor affects pathways associated with energy metabolism in Drosophila melanogaster. Insect Molecular Biology 19(3): 303-313. 10.1111/j.1365-2583.2009.00984.x
Li, L., Zhou, P., Wang, Y., Pan, Y., Chen, M., Tian, Y., et al. 2022. Antimicrobial activity of cyanidin-3-O-glucoside–lauric acid ester against Staphylococcus aureus and Escherichia coli. Food Chemistry 383: 132410. 10.1016/j.foodchem.2022.132410
Lin, Y., Hu, J., Li, S., Hamzah, S.S., Jiang, H., Zhou, A., et al. 2019. Curcumin-based photodynamic sterilization for preservation of fresh-cut Hami melon. Molecules 24(13): 2374. 10.3390/molecules24132374
Lindon, J.C., Holmes, E., and Nicholson, J.K. 2006. Metabonomics techniques and applications to pharmaceutical research & development. Pharmaceutical Research 23(6): 1075–1088. 10.1007/s11095-006-0025-z
Liu, Z., Sun, J., Teng, Z., Luo, Y., Yu, L., Simko, I., et al. 2021. Identification of marker compounds for predicting browning of fresh-cut lettuce using untargeted UHPLC-HRMS metabolomics. Postharvest Biology and Technology 180: 111626. 10.1016/j.postharvbio.2021.111626
Llorach, R., Martínez-Sánchez, A., Tomás-Barberán, F.A., Gil, M.I., and Ferreres, F. 2008. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem 108(3): 1028–1038. 10.1016/j.foodchem.2007.11.032
López, A., Javier, G., Fenoll, J., Hellín, P., and Flores, P. 2014. Chemical composition and antioxidant capacity of lettuce: Comparative study of regular-sized (Romaine) and baby-sized (Little Gem and Mini Romaine) types. Journal of Food Composition and Analysis 33(1): 39–48. 10.1016/j.jfca.2013.10.001
Min, S.C., Roh, S.H., Niemira, B.A., Sites, J.E., Boyd, G., and Lacombe, A. 2016. Dielectric barrier discharge atmospheric cold plasma inhibits Escherichia coli O157:H7, Salmonella, Listeria monocytogenes, and Tulane virus in Romaine lettuce. International Journal of Food Microbiology 237: 114–120. 10.1016/j.ijfoodmicro.2016.08.025
Moodi, Z., Bagherzade, G., and Peters, J. 2021. Quercetin as a precursor for the synthesis of novel nanoscale Cu (II) complex as a catalyst for alcohol oxidation with high antibacterial activity. Bioinorganic Chemistry and Applications 2021: 8818452. 10.1155/2021/8818452
Muna, A.A., Famuyide, I., Wooding, M., McGaw, L.J., and Mühling, K.H. 2022. Secondary metabolite profile and pharmacological opportunities of lettuce plants following selenium and sulfur enhancement. Pharmaceutics 14(11): 2267. 10.3390/pharmaceutics14112267
Otify, A.M., ElBanna, S.A., Eltanany, B.M., Pont, L., Benavente, F., and Ibrahim, R.M. 2023. A comprehensive analytical framework integrating liquid chromatography-tandem mass spectrometry metabolomics with chemometrics for metabolite profiling of lettuce varieties and discovery of antibacterial agents. Food Research International 172: 113178. 10.1016/j.foodres.2023.113178
Pepe, G., Sommella, E., Manfra, M., De Nisco, M., Tenore, G.C., Scopa, A., et al. 2014. Evaluation of anti-inflammatory activity and fast UHPLC–DAD–IT-TOF profiling of polyphenolic compounds extracted from green lettuce (Lactuca sativa L.; var. Maravilla de Verano). Food Chem 167: 153–161. 10.1016/j.foodchem.2014.06.105
Sawada, Y., Akiyama, K., Sakata, A., Kuwahara, A., Otsuki, H., Sakurai, T., et al. 2009. Widely targeted metabolomics based on large-scale MS/MS data for elucidating metabolite accumulation patterns in plants. Plant & Cell Physiology 50(1): 37–47. 10.1093/pcp/pcn183
Shenoy, A.G., Oliver, H.F., and Deering, A.J. 2017. Listeria monocytogenes internalizes in romaine lettuce grown in greenhouse conditions. Journal of Food Protection 80(4): 573–581. 10.4315/0362-028X.JFP-16-095
Shi, M., Gu, J., Wu, H., Rauf, A., Talha, B.E., Khan, Z., et al. 2022. Phytochemicals, nutrition, metabolism, bioavailability, and health benefits in Lettuce—A comprehensive review. Antioxidants 11(6): 1158. 10.3390/antiox11061158
Soberón, J.R., Sgariglia, M.A., Carabajal Torrez, J.A., Aguilar, F.A., Pero, E.J.I., Sampietro, D.A., et al. 2020. Antifungal activity and toxicity studies of flavanones isolated from Tessaria dodoneifolia aerial parts. Heliyon 6(10): e05174. 10.1016/j.heliyon.2020.e05174
Steinmeyer, S., Lee, K., Jayaraman, A., and Alaniz, R.C. 2015. Microbiota metabolite regulation of host immune homeostasis: A mechanistic missing link. Current Allergy and Asthma Reports 15(5): 24. 10.1007/s11882-015-0524-2
Sularz, O., Koronowicz, A., Smoleń, S., Kowalska, I., Skoczylas, Ł., Liszka-Skoczylas, M., et al. 2021. Anti-and pro-oxidant potential of lettuce (Lactuca sativa L.) biofortified with iodine by KIO 3, 5-iodo-and 3, 5-diiodosalicylic acid in human gastrointestinal cancer cell lines. RSC Advances 11(44): 27547–27560. 10.1039/D1RA04679A
Sun, L., Li, H.M., Seufferheld, M.J., Walters, K.R. Jr., Margam, V.M., Jannasch, A., et al. 2011. Systems-scale analysis reveals pathways involved in cellular response to methamphetamine. PLoS One 6(4): e18215. 10.1371/journal.pone.0018215
Tahir, U., Zameer, M., Zahra, N., Mazhar, M., Rafique, A., Alyas, S., et al. 2022. Isolation and characterization of Shiga-toxigenic Escherichia coli isolated from various food samples. Journal of Food and Nutrition Research 10(1): 19–25. 10.12691/jfnr-10-1-3
Taleb, H., Maddocks, S.E., Morris, R.K., and Kanekanian, A.D. 2016. The antibacterial activity of date syrup polyphenols against S. aureus and E. coli. Frontiers in Microbiology 7: 198. 10.3389/fmicb.2016.00198
Tang, C., Xie, B., and Sun, Z., 2017. Antibacterial activity and mechanism of B-type oligomeric procyanidins from lotus seedpod on enterotoxigenic Escherichia coli. Journal of Functional Foods 38: 454–463. 10.1016/j.jff.2017.09.046
Tang, Y., Pan, Y., and Chen, L. 2023. Identification of antibacterial components and modes in the methanol-phase extract from a herbal plant Potentilla kleiniana wight et arn. Foods 12(8): 1640. 10.3390/foods12081640
van Hoek, A.H.A.M., van Veldhuizen, J.N.J., Friesema, I., Coipan, C., Rossen, J.W.A., Bergval, I.L., et al. 2019. Comparative genomics reveals a lack of evidence for pigeons as a main source of stx2f-carrying Escherichia coli causing disease in humans and the common existence of hybrid Shiga toxin-producing and enteropathogenic E. coli pathotypes. BMC Genomics 20(1): 271. 10.1186/s12864-019-5635-z
van Treuren, R., van Eekelen, H.D.L.M., Wehrens, R., and de Vos, R.C.H. 2018. Metabolite variation in the lettuce gene pool: Towards healthier crop varieties and food. Metabolomics 14(11): 1–14. 10.1007/s11306-018-1443-8
Viacava, G.E., Roura, S.I., Berrueta, L.A., Iriondo, C., Gallo, B., and Alonso-Salces, R.M. 2017. Characterization of phenolic compounds in green and red oak-leaf lettuce cultivars by UHPLC-DAD-ESI-QToF/MS using MSE scan mode. Journal of Mass Spectrometry 52(12): 873–902. 10.1002/jms.4021
Xiao, L., Daodong, P., Xuan, W., and Yongfang, Z. 2023. Study on the antibacterial activity and mechanism of bamboo leaf flavonoids on E. coli. China Food Additives. 12: 197–204. 10.19804/j.issn1006-2513.2023.12.026
Xiao, Y., Zhi, Y., Li-wei, G., and Dan-feng, H. 2015. Research progress of antioxidants and its bioactivity in lettuces. China Vegetables 02: 17–24.
Xu, Z., Li, H., Qin, X., Wang, T., Hao, J., Zhao, J., et al. 2019. Antibacterial evaluation of plants extracts against ampicillin-resistant Escherichia coli (E. coli) by microcalorimetry and principal component analysis. AMB Express 9(1): 1–10. 10.1186/s13568-019-0829-y
Xue, W., Yang, C., Liu, M., Lin, X., Wang, M., and Wang, X. 2022. Metabolomics approach on non-targeted screening of 50 PPCPs in lettuce and maize. Molecules 27(15): 4711. 10.3390/molecules27154711
Yang, X., Gil, M.I., Yang, Q., and Tomás-Barberán, F.A. 2022. Bioactive compounds in lettuce: Highlighting the benefits to human health and impacts of preharvest and postharvest practices. Comprehensive Reviews in Food Science and Food Safety 21(4–45): 4–45. 10.1111/1541-4337.12877
Yang, X., Wei, S., Liu, B., Guo, D., Zheng, B., Feng, L., et al. 2018. A novel integrated non-targeted metabolomic analysis reveals significant metabolite variations between different lettuce (Lactuca sativa. L) varieties. Horticulture Research 5: 1–14. 10.1038/s41438-018-0050-1
Yu, L. 2021. Study on the preservation effect of mulberry leaf polyphenols on fresh-cut fruits and vegetables and the mechanism of controlling the adhesion of food-borne pathogenic bacteria. Doctoral dissertation, Southwest University.
Yun, J., Woo, E.-R., and Lee, D.G. 2018. Effect of isoquercitrin on membrane dynamics and apoptosis-like death in Escherichia coli. Biochimica et Biophysica Acta–Biomembranes 1860(2): 357–363. 10.1016/j.bbamem.2017.11.008
Zaman, S., Aziz, A., Siddique, M.A., Khaleque, M.A., and Bari, M.L. 2024. Use of non-chlorine sanitizers in improving quality and safety of marketed fresh salad vegetables. Processes 12(5): 1011. 10.3390/pr12051011
Zeng, W., Vorst, K., Brown, W., Marks, B.P., Jeong, S., Pérez-Rodríguez, F., et al. 2014. Growth of Escherichia coli O157:H7 and Listeria monocytogenes in packaged fresh-cut romaine mix at fluctuating temperatures during commercial transport, retail storage, and display. Journal of Food Protection 77(2): 197–206. 10.4315/0362-028X.JFP-13-117
Zhang, K., Cao, F., Zhao, Y., Wang, H., and Chen, L. 2024. Antibacterial ingredients and modes of the methanol-phase extract from the fruit of Amomum villosum Lour. Plants 13(6): 834. 10.3390/plants13060834
Zhaoyong, L., Xiaoli, Y., Jinglei, L., Boning, L., Keying, Z., and Shuling, W. 2024. Research progress on the chemical composition and pharmacological effects of volatile oil from Atractylodes macrocephala. Chinese Archives of Traditional Chinese Medicine 03: 881–889. https://link.cnki.net/urlid/21.1546.r.20240122.1416.027
Zhidi, C., Sinning, Z., Fan, X., Lianquan, Z., Xinxin, Y., and Xiuzhi, G. 2022. Optimization of extraction technology of polyphenols in leaf lettuce. Journal of Beijing University of Agriculture 1: 111–116. 10.13473/j.cnki.issn.1002-3186.2022.0119
Zhong, L.Q., Wang, L.F., and Qi, C.H. 2009. Talking about the propagation technology of strawberry seedlings. Strawberry Research Progress 3: 269–272.
Zhu, H.L., Chen, G., Chen, S.N., Wang, Q.R., Wan, L., and Jian, S.P. 2019. Characterization of polyphenolic constituents from Sanguisorba officinalis L., and its antibacterial activity. European Food Research and Technology 245(7): 1487–1498. 10.1007/s00217-019-03276-2
Supplementary
Table S1. The top 15 differential metabolites according to the fold change (FC) values.
| BZ3 versus SS | BZ3 versus BS1 | dem | ||||||
|---|---|---|---|---|---|---|---|---|
| Compound name | VIP | FC | log2FC | Compound name | VIP | FC | log2FC | |
| Eriocitrin | 1.5478 | 14.6929 | 3.8770 | Bellidifolin | 1.5718 | 2.1222 | 1.0856 | up |
| Genipin-1-O-gentiobioside | 1.6151 | 16.1411 | 4.0127 | 1,3,6-Trihydroxy-5-methoxyxanthone | 1.5718 | 2.1222 | 1.0856 | up |
| Swertiamarin | 1.7632 | 18.3111 | 4.1946 | Tryptophan | 1.9918 | 2.1383 | 1.0965 | up |
| β-Lactose | 1.7029 | 19.3753 | 4.2761 | N-Acetyl-l-glutamate 5-semialdehyde | 1.5892 | 2.1717 | 1.1188 | up |
| Glycitein | 1.7403 | 30.2168 | 4.9173 | Ganoderenic acid E | 1.8969 | 2.2145 | 1.1470 | up |
| Karacoline | 1.7779 | 36.2173 | 5.1786 | Epitulipinolide | 2.0084 | 2.5130 | 1.3294 | up |
| 1-O-Caffeoylglucose | 1.7549 | 52.5024 | 5.7143 | NG,NG-Dimethylarginine dihydrochloride | 2.0537 | 2.9279 | 1.5499 | up |
| Kaurenoic acid | 1.7836 | 103.1271 | 6.6883 | Gossypetin | 1.5906 | 2.9542 | 1.5627 | up |
| Nervonic acid | 1.6701 | 145.8884 | 7.1887 | Isoquercitrin | 1.9579 | 3.1111 | 1.6374 | up |
| Cynaroside | 1.7707 | 162.0642 | 7.3404 | Delphinidin-3-O-glucoside | 1.9579 | 3.1111 | 1.6374 | up |
| Quercetin-3-O-glucuronide | 1.7900 | 189.2416 | 7.5641 | Picrasin B | 1.8347 | 3.3547 | 1.7462 | up |
| Isoquercitrin | 1.7447 | 201.6518 | 7.6557 | Xanthosine | 2.0793 | 4.6279 | 2.2104 | up |
| Delphinidin-3-O-glucoside | 1.7447 | 201.6518 | 7.6557 | N,1-Methyl-2-pyridone-5-carboxamide | 2.1440 | 5.8412 | 2.5463 | up |
| Schisandrol A | 1.7920 | 295.4953 | 8.2070 | N,1-Methyl-4-pyridone-3-carboxamide | 2.2080 | 5.9725 | 2.5783 | up |
| Methyl hesperidin | 1.7938 | 356.4401 | 8.4775 | Taurohyocholate | 1.8537 | 6.0823 | 2.6046 | up |
| l-Lysine; l-glutamine | 1.7885 | 0.0046 | −7.7596 | Adenosine 2′,3′-cyclic phosphate | 1.9159 | 0.0842 | −3.5709 | down |
| d-Glutamine | 1.7711 | 0.0081 | −6.9547 | 2-(Methylamino)benzoic acid | 2.0001 | 0.1499 | −2.7380 | down |
| Diallyl disulfide | 1.7780 | 0.0116 | −6.4337 | Guanosine 3′,5′-cyclic monophosphate | 1.8678 | 0.1554 | −2.6859 | down |
| l-Asparagine | 1.7885 | 0.0145 | −6.1112 | (+)-Pteryxin | 1.5186 | 0.1774 | −2.4946 | down |
| l-Pipecolic acid | 1.7742 | 0.0181 | −5.7878 | 4-Aminobutyric acid | 1.4646 | 0.2427 | −2.0429 | down |
| Pyrrolidonecarboxylic acid | 1.7742 | 0.0181 | −5.7878 | Physalin D | 2.1688 | 0.2559 | −1.9664 | down |
| trans-4-Hydroxy-l-proline | 1.6564 | 0.0382 | −4.7115 | 7-Ethoxycoumarin | 1.9572 | 0.3047 | −1.7147 | down |
| (S)-2-Aceto-2-hydroxybutanoate | 1.7498 | 0.0396 | −4.6586 | Arecaidine | 1.6429 | 0.3429 | −1.5439 | down |
| dl-Alanine; l-alanine | 1.7611 | 0.0506 | −4.3040 | Fraxin | 2.0829 | 0.3451 | −1.5349 | down |
| l-Pipecolic acid | 1.7301 | 0.0623 | −4.0035 | Vicenin 2 | 1.5794 | 0.3541 | −1.4979 | down |
| Karanjin | 1.7538 | 0.0680 | −3.8786 | Platyphylline | 1.7442 | 0.3565 | −1.4880 | down |
| Aminomalonic acid | 1.7217 | 0.0921 | −3.4411 | Alisol A | 1.5144 | 0.3600 | −1.4740 | down |
| l-Arginine | 1.7564 | 0.0958 | −3.3844 | Rhombifoline | 1.9713 | 0.3827 | −1.3857 | down |
| l-Homoserine | 1.7583 | 0.0980 | −3.3507 | (−)-Cinchonidine | 1.8790 | 0.3938 | −1.3445 | down |
| l-Threonine | 1.7583 | 0.0980 | −3.3507 | Capsidiol | 1.8739 | 0.4183 | −1.2574 | down |
dem, differentially expressed metabolites; VIP, variable importance in projection.