
Download

Review
Substantiable Eruca sativa biomass: from mass spectrometry to insights for biopharmacotherapy
Azizah Abdulaziz Algreiby*
Department of Chemistry, College of Science, Qassim University, Buraydah, Saudi Arabia
Abstract
The use of medicinal plants to treat human ailments has received more attention recently. Eruca sativa (E. sativa; jarjeer) is a very popular edible plant of the Brassicaceae family with remarkable nutritional and therapeutic properties. In this frame, bioactive metabolites derived from E. sativa biomass have captured more attention in pharmacotherapy and biomedicine. This review focuses on recent advances in high-resolution mass spectrometry (HRMS), which have made the characterization of Eruca metabolites, such as glucosinolates and their derivatives, more feasible. Moreover, bio-pharmacological impact, viz. antiulcer, anticancer, antioxidant and antimicrobial, anti-obesity and hypoglycemic activities, cardiovascular benefits, and neuroprotective effects are discussed. This review concentrates all important research findings on these topics, and stresses upon the activities of chemistry and pharmacological areas. By stressing upon additional investigations, this paper emphasizes and appeals researchers and industrials to attempt a comprehensive knowledge of the pharmacological implications of Eruca species and improve their application in modern medicine.
Key words: Eruca sativa, phytochemicals, mass spectrometry measurements, biomedicine, pharmacotherapy potential
Corresponding Author: Azizah Abdulaziz Algreiby, Department of Chemistry, College of Science, Qassim University, Buraydah 51452, Saudi Arabia. Email: [email protected]
Received: 5 September 2024; Accepted: 7 October 2024; Published: 7 November 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Overusage of antibiotics in numerous areas, viz. agriculture and medicine, has caused propagation of drug-resistant bacteria, with extended human travel and reduced global hygiene practices (Bobate et al., 2023; Chatterjee and Haque, 2024). Presently, antimicrobial resistance (AMR) has become a global fear. Millions of people die annually because of opportunistic or major pathogens that develop resistance due to horizontal gene transfer (HGT) and/or biofilm formation (Cangui-Panchi et al., 2023). This has induced global approximation that in 2050 10 million people may die because of infections because of ineffective antibiotics and resistant bacteria (Ahmed et al., 2024).
On the other hand, doubts have arisen about the effects of synthetic chemical compounds on human health. Notwithstanding our incomplete awareness of their impacts on human health, such compounds are available and interconnected to health threats by virtue of their high toxicity and carcinogenic consequences (Cheke et al., 2022). For instance, over the past 40–50 years, the International Agency for Research on Cancer (IARC) has categorized more than 1,000 synthetic agents, with the majority being occupational chemicals and some complex compounds. The assessments have revealed that 50% of them are or possibly carcinogenic to humans, while the rest 50% are not identifiable because of deficient data (Madia et al., 2019). Generally, chemical carcinogens recognized by IARC mainly cover those listed as carcinogens in the European Chemicals Agency (ECHA) inventory, which embody ≈3% of the whole chemicals list (Madia et al., 2019).
Facing all these difficulties, researchers shifted their primary target toward original natural compounds (Badraoui et al., 2020). Lately, phytochemicals have captured more and more attention because of their widespread biological potential (Thiruvengadam et al., 2022). Reviews of some published literature demonstrate that myriad plant-based compounds have a great potential for human health. As a medicinal plant and pertaining to the Brassicaceae family, E. sativa is found globally (Zhu et al., 2021). E. sativa is a diploid annual/biennial herb and is a synonym of Eruca vesicaria subsp. sativa (miller). In the United Kingdom, it is designated as salad rocket, salatrauke (Germany), eruca or oruga (Spain), roquette (France), rucola (Italy), and in the United States, it is named arugula (Testai et al., 2022).
In the Mediterranean basin, E. sativa was extemporaneously cultivated as a leafy vegetable, while in Europe, Northern America, and Asia, it was industrialized for oilseeds (Wells 2024). E. sativa has been recognized for its impact on human health since the Roman period. E. sativa is known for its biological activities, notably stomachic, diuretic, carminative, and to relieve abdominal discomfort, enhance digestion and its aphrodisiac features (Park et al., 2024). In addition, E. sativa has abundant of bioactive compounds, such as antioxidants, vitamins C and E, flavonoids, and glucosinolates (GSLs), with positive effects on human health (Bell et al., 2020). Its protective effect to combat several diseases has been directly linked to their phytochemical composition (Bell et al., 2020). In fact, in response to stress, GSLs are decomposed into bioactive molecules known as isothiocyanates (ITCs), which have antioxidant, anti-inflammatory, and cardioprotective properties (Bell et al., 2020). Similarly, E. sativa reduces the release of inflammatory mediators, and had cytoprotective, antiulcer and antisecretory potential (Pagnotta et al., 2022). The Eruca species comprise GSLs, secondary metabolites that are fragmented by myrosinase enzymes into ITCs and numerous other breakdown compounds (Bell et al., 2020). Regarding these products, their characteristics depend on the specific core of GSL structures, their stability, and the conditions of extraction, such as pH, time, and temperature (Tian et al., 2024). Owing to the existence of high amount of GSL, flavonoids, and carotenoids, various extracts from E. sativa seeds were considered as an abundant source of antioxidants (El-Nwehy et al., 2023). All health benefits offered by E. sativa are attributed to the presence of different phytochemicals in its tissues. 4-Methylthiobutyl glucosinolate (glucoerucin) is the chief GSL extant in E. sativa extracts as well as its oxidized analogues, 4-methylsulfinylbutyl glucosinolate and glucoraphanin (GRP) (Testai et al., 2022).
Recently, the high analytical power triggered by the new mass spectrometry instruments could characterize plant metabolites more feasible. As an illustration, in recent decades, a rapid progress is observed in mass spectrometric (MS) systems permitting analysis and characterization of Eruca natural products. Although the analysis of Eruca natural products has been explored for several years, no review in literature focused on the prospective multifunctional roles and their pharmacological potential, and the useful perspective of high-resolution mass spectrometry (HRMS). Here, we provide information about these topics and their advances and applications that could be interesting for analytical chemistry, natural product communities, and pharmacological commodities.
Literature Review Methodology
Inclusion, exclusion criteria, and relevant screening of studies
The evaluation of original papers was established through screening of the (a) title, (b) abstract, and (c) full text.
Title screening
Thanks to intensive search approach and good numbers of assembled manuscripts, title screening was the first step to eliminate noticeably unsuitable articles.
Abstract screening
In this phase, abstract of articles, which matched subsequent criteria, were chosen. To avoid probable errors in language translation, published studies in English were included. Similarly, the data focusing on “E. sativa,” “mass spectrometry,” and “health benefits” were only selected. Here, unpublished documents were not involved, and the data were assembled for scientific articles only.
Screening of full text
Full texts of available papers, for which abstract screening was completed, were considered and unattainable manuscripts were excluded. Only articles with available full texts were selected for data extraction and analysis. Article was selected if it fulfilled the following criteria: (a) studies involving experiments with E. sativa, mass spectrometry (part 1), and health benefits (part 2); (b) available full-text research articles; (c) original research papers; (d) judging of positive samples and the exact total size; (e) representative type of examined E. sativa; (f) MS as an analytical method; and (g) health benefit. Furthermore, to restrict any faux pas in translation, only English language papers were incorporated, as reported by the earlier meta-analysis studies in analytical chemistry and medical sciences. Papers not meeting the above-mentioned criteria were excluded. In addition, research articles with distinct E. sativa, analytical chemistry by MS, and medicinal capacity, were included. However, the prior condition triggered an extreme limit in pertinent citation numbers. Therefore, the manuscripts were comprised if only E. sativa was assessed in the research paper. To attain references, the EndNote X7® software (Thomson Reuters, Toronto, Canada) was used.
The inclusion criteria reflected that included in part 1, ensuring the identification of articles with experimental designs and publications in English. Regarding part 1, the careful selection process yielded 45 articles that met the established criteria. For part 2, the search identified 37 articles that met the established criteria.
Data extraction
The data were extended and well checked by the author of this work. The collected data from each study were illustrated as study structures corresponding as the first author, year of publication, E. sativa, MS (analyte, quantification, and detection), and health-promoting potential.
In order to search and recoup the research papers associated to “E. sativa”, “mass spectrometry,” and “health benefits,” the array of key collection of online Scopus database was explored, and only these criteria were selected. Studies pertaining to conference abstracts, letters, patents, or review articles were not accepted.
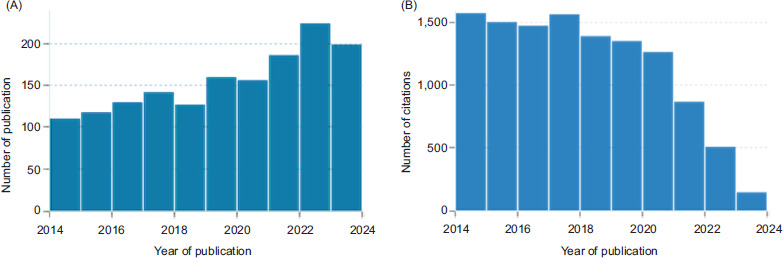
Between 2014 and 2024, the number of publications augmented exponentially. Slow but stable growth achieved its first maximum in 2023, with n = 220 articles cited, and c = 1,500 times in 2014 and 2018. This was clarified by the fact that consumers preferred “Natural” for preventatives than for curatives (Figures 1A and B).
Figure 1. (A) Number of publications and (B) citations of E. sativa, mass spectrometry, and health benefits between 2014 and 2024. Source: www.lens.org. Accessed on 26 June 2024.
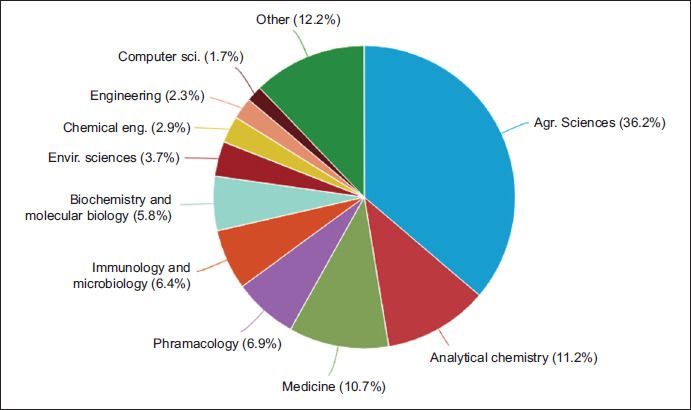
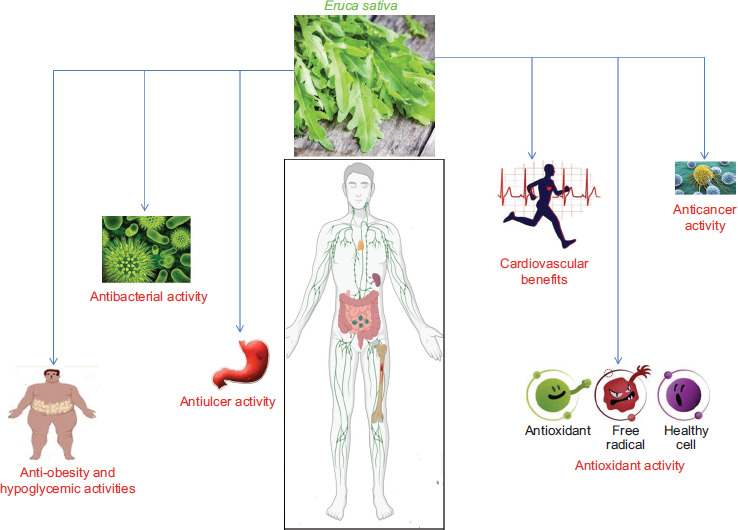
From the utilized 876 publications, at least 87.8% were explicitly about E. sativa, MS, and medicinal potential. The research areas were evaluated according to the Journal Citation Report (JCR) assignments. The most allocated JCR subject category was (i) agricultural sciences, (ii) analytical chemistry, and (iii) medicine and pharmacology, with 65% of the total of topics (Figure 2). The increasing attention to E. sativa in medicine is linked to various factors, such as beneficial impact on various diseases (e.g. antiulcer, anticancer, antioxidant, antimicrobial, anti-obesity, and hypoglycemic activities, and cardiovascular and neuroprotective benefits) as well as their phytochemical characterization by suitable analytical chemistry tools (such as MS measurements).
Figure 2. Journal Citation Report (JCR) subject categories related to the E. sativa, mass spectrometry, and medicinal potential. The data were obtained from Scopus database on 26 June 2024.
A varied collection of scholarly literature is within the area of science and archived in numerous databases, which were examined extensively. Here, a comprehensive inspection approach was engaged through international databases. In view of its position as an authoritative and extensively employed database that includes worldwide citations and research publications, the author of this review decided to apply Web of Science (WoS), considered as the largest accessible citation database. The search was subdivided into two parts. Part I concentrated on an in-depth collective expertise on the analytical techniques employed in identification and characterization of phenolics from E. sativa. To guarantee robust study design, the used keywords in this search were “E. sativa,” “analytical chemistry,” and “mass spectrometry.” Part II of the search strategy examined the health-promoting potential of the Eruca species. The corresponding keyword search engaged terms such as “Eruca extract,” “medicinal, pharmaceutical, therapeutic,” and “health benefits.”
A bibliometric analysis of the Lens (www.lens.org) displays that between 2014 and 2024 (26 June 2024), a total of 1,820 peer-reviewed scholarly works were published on “E. sativa,” OR “mass spectrometry,” OR “health benefits” (Figure 3).
Figure 3. Publication subject linked to E. sativa, OR mass spectrometry, OR health benefits. Source: www.lens.org. Accessed on 26 June 2024.
An Insight into Qualitative and Quantitative Profiles of Phenolic Compounds in E. sativa
Kumar et al. (2022) explored the phytochemical composition and anti-oxidant benefits of E. sativa grown under different environment conditions in India: high mountain Ladakh region (3,524 m, Leh–Ladakh) vs. low-altitude region (321 m, Chandigarh). The solubility of polyphenols in polar solvents is more than that in nonpolar solvents, suggesting that polar solvents extract large amounts of phenolic compounds (Kaczorová et al., 2021; Palaiogiannis et al., 2023). In this way, the cited authors indicated that the efficiency of solvents for extraction of phenolic compounds was in the following order: aqueous > 70% methanol (MeOH) > chloroform (CHCl3) > Hex. Compared to Chandigarh plants, aqueous extract of Leh–Ladhak-grown plants showed highest values of total phenolic content (TPC) (~32 μg gallic acid equivalent [GAE]/mg) and total flavonoid content (TFC) (~33 µM RE/mg) whereas 70% MeOH extract showed higher value of total antioxidant capacity (TAC) (~120 μg AAE/mg). Samples grown at these two different altitudes were statistically significant. Regarding TPC, E. sativa grown in Leh–Ladhak had a significantly higher levels of aqueous extract (32 μg/mg), 70% MeOH (29 μg/mg), chloroform (10 μg/mg), and Hex (8.05 ± 0.30 μg/mg) compared to the Chandigarh-grown E. sativa. Kumar et al. (2022) confirmed that great altitude displayed higher TPC than lower altitude.
It is reported that exposure of plants to ultraviolet-B (UV-B) radiation enhances the concentration of some phenolic compounds, including secoiridoids and hydrocinnamic acid, which protect plants from the deleterious effects of UV-B stress (Dias et al., 2019). Several studies demonstrated that plants grown at higher altitudes are revealed to intense UV-B radiation, leading to great consequences of plant growth, physiology, and morphology. The defense mechanism of plants is activated by intense UV-B radiation and therefore a higher production of polyphenol compounds (Testai et al., 2022).
Elahdef et al. (2020) revealed that significant differences were observed in the phytochemical contents among E. sativa populations collected from seven natural Tunisian geographic locations. The cited authors reported that environmental conditions, such as altitude, average annual precipitation (AAT), average annual temperature, and relative humidity, influenced phytochemical contents. Hence, altitude and average annual precipitation had a positive impact on TFC, and any increase in average annual precipitation was directly linked to TPC, TFC, and TAC.
Zidorn and Stuppner (2001) collected Leontodon (Cichorieae) in Austria from diverse altitudes (1013–3855 m) and demonstrated a significant relationship between altitudes of collection sites and TFC. Dong et al. (2011) investigated variation of TFC in Eucommia ulmoides leaves grown at six different altitudes (550–1,180 m) and showed that the altitude was correlated to TFC. Ghasemi et al. (2011) discovered a good coefficient of correlation between TFC and altitudes (750–2,465 m) in MeOH extract of walnut green husks (Juglans regia L.) obtained from 11 different geographical locations with differing climatic conditions in Iran. Likewise, Pandey et al. (2018) perceived comparable conclusions for Thalictrum foliolosum extracts, and demonstrated that samples from higher altitudes had more phenolic compounds than those from lower altitudes. Kumar et al. (2007) also reported variation of TFC in E. sativa extracts from the samples grown at two different altitudes. Similar trends were reported by Kumar et al. (2022), which E. sativa grown in Leh and Chandigarh were comparable to those for 32 Eruca accessions, with an average value of 2353 mg/100 g DW, and a range that varied from 999 to 3139 mg/100 g DW (Pasini et al., 2012). Additionally, TFC of fresh E. sativa showed a broader range because its content is influenced remarkably by exposure to UV light and cyclical variations, in addition to soil growth conditions (Bennett et al., 2006).
Sadiq et al. (2014) assessed the phytochemical profile of aqueous and methanolic extracts of E. sativa stems, leaves, flowers, and seeds. Remarkably, findings evidenced that all parts of E. sativa plant were abundant in steroids, terpenoids, tannins, diterpenes and glycosides, polyphenols, alkaloids, flavonoids, and phytosterols. In addition, TPC of all aerial parts revealed that they had a good quantity of phenolics, particularly in seeds (27 μg GAE/mg) and leaves (23 μg GAE/mg). Table 1 summarizes selected outcomes of phenolic contents reported in Eruca sativa in recent years.
Table 1. Some examples of phenolic contents reported in Eruca sativa.
| Source | Plant Part | Solvent Extraction | Analyte | Method/Detection | Quantification | Reference | |
|---|---|---|---|---|---|---|---|
| Leh–Ladakh (India) | Whole plant | Hexane Chloroform Water 70% Methanol |
TPC (GAE μg/mg of DPE) | Folin–Ciocalteu (765 nm) | 8.05 10.01 31.9 28.79 |
Kumar et al., 2022 | |
| TFC (RE [µM/mg of DPE]) | Aluminium chloride colorimetric technique (415 nm) | 14.6 14.77 35.54 21.6 | |||||
| Chandigarh (India) | Hexane Chloroform Water 70% Methanol |
TPC (GAE μg/mg of DPE) | Folin–Ciocalteu (765 nm) | 6.62 8.59 16.82 25.48 | |||
| TFC (RE [µM/mg of DPE]) | Aluminium chloride colorimetric technique (415 nm) | 3.21 7.54 19.38 15.04 | |||||
| Islamabad (Pakistan) | Stem Leaf Flower Seed | 85% Methanol | TPC (mg GAE/g) | Folin–Ciocalteu (640 nm) | 13.55 23.07 19.9 27.1 |
Sadiq et al., 2014 | |
| Kedarkantha (India) | Whole plant | 95% EtOH 50% EtOH Water |
TPC (GAE/g extract) | Folin–Ciocalteu (765 nm) | 26.57 18.4 1.54 |
Pandey et al., 2018 | |
| TFC (RE/g extract) | Aluminium chloride colorimetric technique (415 nm) | 8.28 6.36 5.64 | |||||
| Berberine content (μg/mg) of crude sample | HPTLC mobile phase, isopropanol:formic acid:water (9:2:1) | 7.37 6.84 5.1 | |||||
| Taluka (India) | 95% ethanol, 50% ethanol Water |
TPC (GAE/g extract) | Folin–Ciocalteu (765 nm) | 25.42 16.8 1.4 | |||
| TFC (RE/g extract) | Aluminium chloride colorimetric technique (415 nm) | 7.96 6.02 5.28 | |||||
| Berberine content (μg/mg) of crude sample | HPTLC mobile phase, isopropanol:formic acid:water (9:2:1) | ||||||
| Sankri (India) | 95% ethanol, 50% ethanol Water |
TPC (GAE/g extract) | Folin–Ciocalteu (765 nm) | 24.7 15.62 1.24 | |||
| TFC (RE/g extract) | Aluminium chloride colorimetric technique (415 nm) | 7.2 5.63 5.09 | |||||
| Berberine content (μg/mg) of crude sample | HPTLC mobile phase, isopropanol:formic acid:water (9:2:1) | 9.76 9.42 6.80 | |||||
| Bhowali (India) | 95% ethanol, 50% ethanol Water |
TPC (GAE/g extract) | Folin–Ciocalteu (765 nm) | 21.27 12.52 10.01 | |||
| TFC (RE/g extract) | Aluminium chloride colorimetric technique (415 nm) | 6.94 5.24 4.82 | |||||
| Berberine content (μg/mg) of crude sample | HPTLC mobile phase, isopropanol: formic acid: water (9:2:1) | 11.1 10.8 7.84 | |||||
| Iran | Leaves | Water | TPC (mg TAE/g DW) | Folin–Ciocalteu (765 nm) | 28–42 3.76–5.11 |
Nikzad et al., 2023 | |
| TFC (mg QE/g DW) | Aluminium chloride colorimetric technique (510 nm) | ||||||
| Ethiopia | Leaves | Water | TFC (mg of CE/g) | Aluminium chloride colorimetric technique (510 nm) | 14.03 0.25 |
Keyata et al., 2021 | |
| TAC (mg of cyanidin-3-glucoside per gram) | pH dependence of color change of anthocyanins (520 and 700 nm) | ||||||
| Egypt | Leaves | ethanol | TPC (mg GAE/g) | Folin–Ciocalteu (725 nm) | 12.522 9.938 |
El-Gayar et al., 2022 | |
| TFC (mg CE/g) | Aluminium chloride colorimetric technique (510 nm) | ||||||
TPC: Total polyphenolic content, GAE: gallic acid equivalent, TFC: Total flavonoid content, RE: rutin-trihydrate equivalent.
Advanced Analytical Methods for Detection of Phenolic Compounds in Eruca Species
Establishing the richness of bioactive compounds at any particular point of Eruca life cycle and crop management could provide understanding of how metabolic variations may impact the health. Since these biological compounds are quiet challenging to acquire, and their isolation and extraction are expensive, it is generally decided that MS is a major requirement for their identification and characterization. It is equal to employ tandem mass spectrometry (MS/MS) or nuclear magnetic resonance (NMR) studies, which permit total confirmation and revelation of molecular structures (Bouanani et al., 2023). Through high-performance liquid chromatography–diode array detection–electrospray ionization/tandem mass spectrometry (HPLC-DAD-ESI-MS/MS), Pasini et al. (2012) identified GSLs in 32 Italian E. sativa accessions. Eight desulfo-glucosinolates (DS-GSLs) were marked in all extracts. Findings exposed four aliphatic-derived compounds (glucoraphanin, glucoalyssin, glucoerucin, and progoitrin/epiprogoitrin), one aromatic GSL (glucosinalbin), two indole-derived compounds (4-OH-glucobrassicin and glucobrassicin), and two structurally related compounds covering one intermolecular disulfide linkage (4-[β-D-glucopyranosyldisulfanyl] butyl-GSL and dimeric 4-mercaptobutyl-GSL).
Pasini et al. (2012) reported that all Eruca accessions displayed a large variation in TFC, that is, from 23 to 31 g/kg DW. The ampler flavonoid group was characterized by kaempferol derivatives, with kaempferol-3,4-diglucoside being the main flavonoid in all samples, ranging from 8 to 23 g/kg DW. The second abundant flavonoid was isorhamnetin-3,4-diglucoside, with 9–16% TPC. Bell et al. (2015) reported that the total average flavonol content in Eruca sample ranged from 0.5 to 3.8 g/kg DW. It was demonstrated by liquid chromatography–mass spectrometry (LC-MS) that Eruca accumulated kaempferol glucosides, and myricetin was the predominant glucoside.
For GSL, the monomeric and dimeric forms of glucosativin (4-mercaptobutyl-GSL) were identified and quantified separately. Fechner et al. (2018) reported that glucosativin and 3-thiazepane-2-thione (sativin), its hydrolysis product, are liable for the distinctive flavor and aroma of Rocket “Eruca sativa L” (EER). Bell et al. (2017) carried out the broadest sensory analysis of seven E. sativa accessions by grouping with numerous chemical analyses. This elucidated some of the associations between specific traits of EER, such as GSLs hydrolytic products (GHPs) and volatile sulfur compounds with mustard and hotness flavors. Other relation detected between green leaf volatiles and free amino acids was a decrease of apparent pungency, and GSLs/GHPs were reported to reduce bitterness and hotness perceptions. A study conducted by Raffo et al. (2018) reported that sativin was the main cause of flavor and aroma in EER.
Kim and Ishii (2006) identified two DS-GSLs, DS-glucoraphanin, and DS-glucoerucin by HPLC-ESI-MS. These molecules were confirmed to be the principal GSLs in E. sativa seeds and roots, while three DS-GSLs (DS-glucoraphanin, DS-glucoerucin, and DS-dimeric 4-mercaptobutyl GSL) were the core compounds detected in leaves. By using LC/ESI-quadrupole ion-trap mass spectrometry (QIT-MS), a study conducted by Cataldi et al. (2007) revealed 12 species of GSLs in E. sativa. Seven aliphatic compounds, named glucoraphanin (m/z 436), glucoerucin (m/z 420), 4-mercaptobutyl-GLS (m/z 406), progoitrin/epiprogoitrin (m/z 388), sinigrin (m/z 358), 4-methylpentyl-GSL, and n-hexyl-GSL (m/z 402). In addition, two indole GSLs (4-(β-D-glucopyranosyldisulfanyl)butyl-GLS [m/z 600] and dimeric 4-mercaptobutyl-GLS [m/z 811]) were detected. It was an MS2 fragmentation experiment and chromatographic parting from 4-hydroxyindol-3-ylmethyl-GSL.
Abd-Elsalam et al. (2021) determined chemical components of the ethanolic extract of E. sativa L. seeds by LC-MS. The chemical profile revealed the existence of numerous classes as sulfur-containing compounds, including derivative compounds of glucosinalates, viz. glucoerucin, glucoalyssin, desulfated glucoraphanin, desulfated sinidrin, and desulfated (glucosyl-disulfonyl)-butyl glucosinolate, and flavonoids that fit to O-glycosides and their aglycones. In addition, the corresponding profile revealed some phenolic compounds, such as caffeoyl-O-hexoside and chlorogenic acid and fatty acids (e.g. tri-hydroxy-octadecenoic, octadecatetraenoic, linoleic, arachidic, and erucic acids).
Martínez-Sánchez et al. (2007) examined the chemical profile of E. vesicaria leaves. The MS/MS study endorsed the elucidation of quercetin mono- and diacyl-tri-O-glucosides and flavonoid glycosylation. The cited authors proved that E. vesicaria included kaempferol derivatives as principal compounds. In order to quantify glucosinolates concurrently, and the corresponding ITCs in E. sativa defatted seed meals (DSM), Franco et al. (2016) developed a new high-pressure liquid chromatography–electrospray ionization–tandem MS (HPLC-ESI-MS/MS) method. Interestingly, the new approach was suitable to quantify glucosinolates and ITC derivatives jointly in biomasses and bakery products. Sharma et al. (2017) evolved a simple and rapid ultra performance liquid chromatography (RP-UPLC) to synchronously quantify erucin (4-methylthiobutyl isothiocyanate; ER), allyl isothiocyanate, and benzyl isothiocyanate in E. sativa oil. Erucin was detected as the main component (~29%) in E. sativa oil, while allyl isothiocyanate and benzyl isothiocyanate were observed at very low levels (0.06% and 0.07%, respectively). Chemically, in the presence of myrosinase, glucoerucin breaks down into erucin and other active hydrolytic products, such as 4-methylthiobutyl thiocyanate and 4-methylthiobutyl nitrile (Maycotte et al., 2024). Sharma et al. (2017) confirmed the presence of eight compounds in methanolic E. sativa extract by using UPLC-MS/MS. Glucoerucin was the only detected isothiocyanate at m/z 421, with perceived caffeoyl glucose (at m/z 203), 3-caffeoylquinic acid (at m/z 191), and sinapicglucoside (at m/z 223. In addition, apigenin-7-O-glucoside, isorhamonetin-3-O-rutinoside, kaempferol-3-O-glucuronide, and Isorhamonetin-3-O-(3”-acetylglucoside) were detected as four flavonoids derivatives in the methanolic extract of E. sativa.
In a study conducted by Crescenzi et al. (2023), it was reported that through LC-ESI-Quadrupole(Q)-Exactive-MS/MS, the methanol and hydroalcoholic profiles of E. sativa extracts exhibited a good number of bioactive compounds, such as glycosylated flavonoids (quercetin 3,30,40-O-triglucoside, quercetin-3,40-O-diglucoside-30-O-(6-sinapoyl-glucoside), and quercetin diglucoside). Besides, E. sativa cultivated in the Piana del Sele (Salerno, Italy) presented the peak value of glucosinolates, with a talented aptitude to produce erucin by its myrosinase conversion. Crescenzi et al. (2023) separated different geographical E. sativa origins by using the archived biomarker metabolites profiling of hydroalcoholics. Glucosinolates and flavonols were controlled by harvest time, culture type, and production cycle (Buitrago-Villanueva et al., 2023). These differences established the conditions to select culture system and E. sativa crops. Herr and Büchler (2010) reported that glucosinolates levels could vary gradually depending on environmental conditions. Furthermore, E. sativa cultivated in Piana del Sele showed the highest level of Glucoerucin (GER), with a promising ability to generate erucin through its conversion by myrosinase action. Sut et al. (2018) extracted glucosinolates by using supercritical carbon dioxide (SCO2) as a combination of water. Sut et al. (2018) pointed out that maximum glucosinolate found in E. sativa leaves was in the dimeric form of 4-mercaptobutylglucosinolate (DMB) glucosativin and glucoerucin.
Ramazzina et al. (2022) investigated the impact of plasma-activated water (PAW) on polyphenolic profile of E. sativa. In this way, the cited authors explored qualitative/quantitative polyphenolic profile by ultra-high-performance liquid chromatography (UHPLC)-MS/MS analysis. It was performed on the MeOH extracts acquired after revealing matrix to PAW for 20 min (PAW-20). Detected polyphenol correlated to glycosylated flavonols (kaempferol, isorhamnetin, and quercetin). From E. sativa seeds, Khaliq et al. (2021) defined napin (EsNap), a low molecular mass protein that fits to prolamin super family. EsNap was isolated by 70% (NH4)2SO4 precipitation and size-exclusion chromatography. The nano LC-MS/MS analysis generated two peptides (IYQTATHLPK10 and QQQGQQGQQLQQVISR16) that included 26 residues. It should be noted that EsNap family is characteristically rich in Arg, Lys, and Cys amino acid residues.
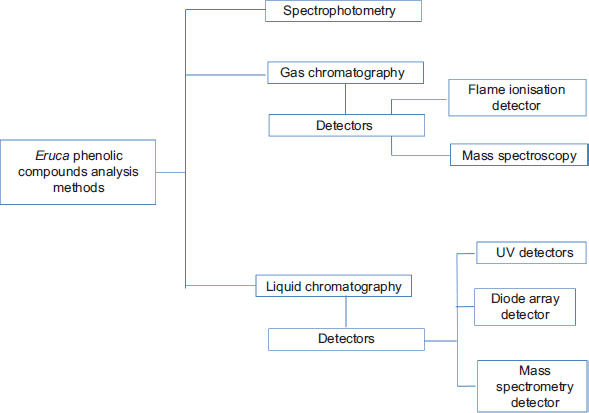
Fagerlund et al. (2021) examined anti-listerial activity in E. sativa, and concluded that the active fraction was retained in polar chromatographic conditions and comprised several metabolites. The elucidation of this fraction, using LC-MS/MS, led to identification of compounds containing 19 nucleosides and amino acids: 11 amino acids, a dipeptide, a quaternary ammonium compound, an amine, three nucleosides, and two nucleobases. In summary, Table 2 presents some frequently used methods to quantify and identify phenolic compounds (chromatographic conditions; mobile phase and gradient; quantification and detection; and analytical method) from different extracts of Eruca species. As shown in Figure 4, many dependable quali-quantitative approaches are available for the measurement and characterization of phenolic contents/compounds from Eruca species.
Table 2 Selected high-resolution mass spectroscopy (HRMS) in the characterization and identification of phenolic compounds in Eruca species.
| Source | Analyte | Chromatographic conditions | Quantification | Detection | References | |
|---|---|---|---|---|---|---|
| Mobile phase | Gradient | |||||
| Italy | Glucosinolates and phenolic compounds | Water (mobile phase A)–acetonitrile (mobile phase B) | 99% A, linear gradient to 75% A at min 17.5, linear gradient to 99% A at min 20; total analysis time: 35 min | HPLC-DAD–ESI-MS | 229 nm | Pasini et al., 2012 |
| UK | Glucosinolate and flavonol | Ammonium formate (0.1%) (mobile phase A)–acetonitrile (mobile phase B) | Isocratic gradient: 95% and 5% | LC-MS | 229 and 330 nm, respectively | Bell et al., 2015 |
| Egypt | Sulfur-containing compounds, flavonoids, phenolic acid, and fatty acids | Water with 0.1% formic acid (mobile phase A) and methanol with 0.1% formic acid (mobile phase B) | – | LC-ESI-MS | Between 200 nm and 400 nm | Abd-Elsalam et al., 2021 |
| Italy | Desulfo-glucosinolates, phenolic acids, and flavonoids | Phosphoric acid 103M (mobile phase A)–acetonitrile 95:5 (mobile phase B) | B: 0–2 min, 5% B (isocratic); (2–4.5 min) at 10% B; at 5.5 min, 15% B; at 9 min, 35% B; at 11 min, 55% B; from 12 min to 13 min at 70% B (isocratic) | HPLC | 229, 280, and 320 nm, respectively | Testai et al., 2022 |
| Italy | Glucosinolate (GSL) and desulfo-glucosinolate | 0.5% Formic acid (mobile phase A)–acetonitrile with 0.5% formic acid 5 (mobile phase B) | B: 10 min at 5%, 4 min at 24%, 4 min at 50%, 7 min at 80%, 10 min at 5% | HPLC–ES-MS/MS | 229 and 365 nm, respectively | Franco et al., 2016 |
| India | Erucin, allyl isothiocyanate, and benzyl isothiocyanate | 0.1% HCOOH in water acid (mobile phase A)–0.1% HCOOH in methanol (mobile phase B) and pure acetonitrile (mobile phase C) | - | UPLC-DAD and UPLC-ESI-QTOF | 280 nm | Sharma et al., 2017 |
| Italy | Glucosinolates, glycosylated flavonoids, fatty acids, and lipids | Water + 0.1% formic acid (mobile phase A)–acetonitrile + 0.1% formic acid (mobile phase B) | B: 0–23 min, from 5 to 40%; 23–45 min, from 40 to 95%; and then back to 5% for 10 min | UHPLC-Q-Exactive-Orbitrap-MS/MS | 280 nm | Crescenzi et al., 2023 |
| Italy | Glucosinolates | Water with 0.1% TFA (mobile phase A)–MeOH with 0.1% TFA (mobile phase B) | 0–10 min: 100%:0% (A:B) linear change to 80%:20% (A:B, v/v); 10–15 min:50:50; 16–20 min increased to 100% B; 21–25 min: change to solvent A 100% | Electrospray ionization (ESI)–ion trap mass spectrometry (ITMS) (ESI-ITMS) | Between 200 nm and 400 nm | Cataldi et al., 2007 |
| India | Seed oil: isothiocyanates and free fatty acids | – | Temperature programming was from 50°C (hold 5 min) to 290°C at 6 C/min; the transfer line and ion trap were at 180°C | HS/SPME/GC–MS | Between 200 nm and 400 nm | Khoobchandani et al., 2010 |
| Saudi Arabia | Vitamins, fatty acids, alkaloids, flavonoids, terpenoids, and phenols | 1% Formic acid in deionized water (mobile phase A) and acetonitrile (mobile phase B) | – | HR-LC/MS | Between 200 nm and 400 nm | Awadelkareem et al., 2022a |
| India | Vitamins, fatty acids, alkaloids, flavonoids, terpenoids, and phenols | Methanol (mobile phase A)–water (0.05% formic acid) (mobile phase B) | – | HPTLC | 366 nm | Awadelkareem et al., 2022b |
| Italy | Phenolic compounds | Water + 0.2% formic acid (mobile phase A) and acetonitrile + 0.2 formic acid (mobile phase B) | A 9-min linear gradient of 2–20% B in 0.2% A | UHPLC-ESI-MS/MS | Between 200 nm and 400 nm | Ramazzina et al., 2022 |
| Colombia | Glucosinolates | 0.05% Formic acid (mobile phase A) and 0.05% HCOOH in acetonitrile (mobile phase B) | 0 min 5% B, then B was progressively raised to 95% at 55 min and kept at this value for 5 min, and finally, B 5% kept till 65 min | LC-MS | 270 nm | Buitrago-Villanueva et al.,2023 |
| Iran | Phenolic compounds | Formic acid (0.1%) (mobile phase A)–acetonitrile (99.8%) (mobile phase B) | 10–26% B for 40 min, 65% B for 70 min, and finally 100% B for 75 min | HPLC-MS | Between 200 nm and 400 nm | Motalebnejad et al.,2023 |
| Pakistan | Peptides | 0.1% Formic acid in H2O (mobile phase A)–0.1% formic acid (in acetonitrile) with 2% (mobile phase B) | 2–30% B in 35 min | LC–MS/MS | – | Khaliq et al., 2021 |
| Hungary | Glucosinolates | Water with 0.1% (v/v) formic acid, (mobile phase A)–acetonitrile with 0.1% (v/v) formic acid (mobile phase B) | – | UPLC-ESI-IMS-QTOF-MS | – | Dernovics et al., 2023 |
| Italy | Phenolic compounds | Water (mobile phase A) and acetonitrile (mobile phase B), both acidified with 0.1% (v/v) formic acid | 95% (v/v) of A and 5% (v/v) of B; varied linearly to 25% A and 75% B in 25 min | LC–PDA–MS | – | Pane et al., 2020 |
| Norway | Nucleosides and amino acids | Water, 0.1% formic acid (mobile phase A)–acetonitrile, 0.1% formic acid (mobile phase B) | 0 min, 5% B; 15 min, 5% B; 20 min, 95% B; 30 min, 95% B; 31 min, 5% B; 34 min, 5% B | LC-MS/MS | 227 nm, 254 nm, 280 nm, and 330 nm | Fagerlund et al., 2021 |
| Republic of Korea | Flavonols | 100-mM formic acid in deionized water (mobile phase A)–acetonitrile (mobile phase B) | 0 (15% B) to 45 min (40% B), from 75 to 84 min (98% B), and from 85 to 90 min (15% B) | HPLC-UV-MS | 371 nm | Park et al., 2024 |
| India | Phenolic contents | Aqueous formic acid 0.1% (mobile phase A)–formic acid 0.1% in acetonitrile (mobile phase B) | 0 min: A 95%, B 5%; 2 min: A 95%, B 5%; t = 32 min: A 40%, B 60%; t = 47 min: A 0%, B 100%; t = 55 min: A 0%, B 100%; t = 60 min: A 95%, B 5% | UPLC-ESI-MS/MS | – | Khoobchandani et al., 2011 |
HPLC-DAD–ESI-MS: High-performance liquid chromatography coupled to photodiode array and electrospray ionization mass spectrometric; LC-MS: Liquid Chromatography–mass spectrometry; LC-ESI-MS: Liquid chromatography–electrospray ionization–tandem mass spectrometry; HPLC: high-performance liquid chromatography; LC-ESI-MS: HPLC-Electrospray Ionization-Mass Spectrometry; UPLC-DAD-MS: Ultra-high-performance liquid chromatography–diode array detector–tandem mass spectrometry; UPLC/ESI-Q-TOF-MS: Ultra Performance Liquid Chromatography/Electrospray Ionization-Quadrupole-Time Of Flight-Mass Spectrometry; UHPLC-Q-Exactive-Orbitrap MS/MS: Ultra-high-performance liquid chromatography coupled with hybrid quadrupole-orbitrap tandem mass spectrometry; HS/SPME/GC–MS: Headspace Solid-Phase Microextraction (HS-SPME) and Gas Chromatography/Mass Spectrometry Analysis; HR-LC/MS: High Resolution-Liquid Chromatography Mass Spectrometry; HPTLC: High-performance thin layer chromatography; LC/ESI-MSn: liquid chromatography-electrospray ionization/multi-stage mass spectrometry; UHPLC-ESI-MS/MS: Ultrahigh-performance liquid chromatography tandem mass spectrometry with electrospray ionization; UPLC-ESI-IMS-QTOF-MS: ultra-high-performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight mass spectrometry; LC-PDA-MS: Liquid chromatography-photodiode-array-mass spectrometry; HPLC-UV-MS: High-performance liquid chromatography (HPLC) with UV detection coupled with electrospray ionization tandem mass spectrometry; UPLC-ESI-MS/MS: Ultra-performance liquid chromatography-electrospray ionization tandem mass spectrometry.
Figure 4. Methods employed for qualitative and quantitative analysis of various phenolic compounds in Eruca species.
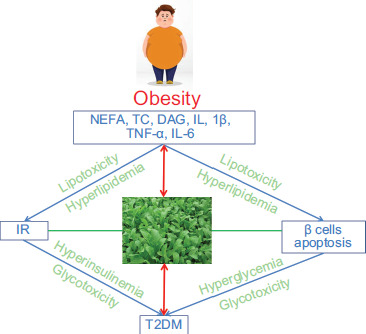
Figure 5. Mechanism of E. sativa extracts as antidiabetics. E. sativa extracts prevent the pathological progress of obesity and insulin-resistance β cells apoptosis in diabetes. IR: insulin resistance; T2DM: type 2 diabetes mellitus.
To determine the effectiveness of polyphenolic contents as health-promoting molecules, it is significant to have extraction methods and exact bioactive molecules from Eruca species. Organic solvents used for the extraction of major molecules with different activities are the most acceptable solvents for extraction from Eruca species. Owing to the chemical multiplicity of phenolic compounds and the complication of composition in Eruca species, it is expensive and difficult to separate each phenolic compound and investigate it independently. Moreover, a combined total antioxidant power of a multifaceted sample is frequently more expressive to assess the health benefits because of the cooperative action of bioactive compounds.
Health-Promoting Potential of Eruca Species
Nutrition assumes a crucial part in human health and impacts the evolution and development of chronic diseases. Therefore, utilization of plant extracts and nutritional supplements could decrease the risk of fisease. Thanks to the presence of different E. sativa varieties, its consumption has demonstrated different health benefits. In this section, the potential use of E. sativa as a therapy in some human health condition is discussed.
Antiulcer activity
Helicobacter pylori is the commonest trigger of chronic gastritis and variably results in serious gastroduodenal disorders in some patients (Kim, 2024). This latter could comprise duodenal peptic ulcers, gastric cancer, and gastric mucosa-associated lymphoid tissue lymphoma. In H. pylori colonization, numerous mechanisms, counting motility, urease production, and adhesion are discussed (Li et al., 2024). In traditional medicine, E. sativa has an important role to kill bacteria. Urease enzyme is essential for the growth, metabolism, and colonization in gastric mucosa. Khan and Khan (2014) investigated the impact of urease inhibition of crude extract and fractions of E. sativa in an in vitro experimental model. Extract of E. sativa was determined to provide protection to gastric mucosa against ulceration triggered by numerous necrotizing agents considering ethanol (EtOH) and strong alkalis. EtOH-induced gastric ulcers have been employed in the evaluation of gastroprotective activity. In this regard, it has been concluded that O2-derived free radicals are concerned in the mechanism of acute and chronic stomach ulceration (Zhang et al., 2021). The EtOH genesis can provoke gastric lesions of multifactorial origin with a decrease in gastric mucus, and is related to the production of free radicals, resulting in an increase of lipid peroxidation (LPO), which causes damage to cells and cell membranes (Zhang et al., 2021). The cytoprotective effect of E. sativa extract may be associated to its ability to prevent gastric acid secretion and/or enhance mucosal defensive factors, such as prostaglandins, and a decrease in LPO (Alqasoumi et al., 2009). Some investigations confirmed that flavonoids may be related to the antiulcer activity (Madjo et al., 2023) and have a main role in the mechanism of gastroprotection (Madjo et al., 2023). In addition to flavonoids, other compounds in EER, such as sterol and/or triterpenes, are recognized for their antioxidant activities, which may contribute to some of the antiulcer mechanisms (Singh et al., 2022).
With half maximal inhibitory concentration (IC50) = 7.77 mg/mL, the crude extract displayed a patent inhibition against urease. On the other hand, the dominant fraction was ethyl acetate was followed by the aqueous extract at IC50 = 4.17 and 5.83 mg/mL. In a study, Alqasoumi et al. (2009) studied the anti-ulcerogenic property of EER in diverse ulcer mice models. In pylorus-ligated Shay mice, the EtOH extract of EER meaningfully decreased the basal gastric acid secretion, titratable acidity and ruminal ulceration. EER could reduce gastric ulceration and indomethacin- and hypothermic-restraint stress, thus histologically confirming its antiulcer effect. Similarly, pretreatment with EER hindered EtOH-induced necrosis in the superficial layers of gastric mucosa with congestion. On the other hand, the extract significantly refilled the levels of gastric wall mucus, non-protein sulfhydryl, and malondialdehyde.
Saleh et al. (2016) reported in animal models that by maintaining the acid–base balance of gastric contents, the leaf extracts of E. sativa were found to display antiulcer activity against ethanol-induced gastric mucosa injury.
Antibacterial activity
Malik (2015) investigated the activity of E. sativa seeds against S. aureus and B. cereus. Interestingly, methanolic extracts showed an inhibition zone of 3–8 mm. Regarding minimum inhibitory concentration (MIC) values, the concentrations were 80 µg/mL and 20 µg/mL against S. aureus and B. cereus, respectively. Koubaa et al. (2015) studied the potential of EER flowers against 11 pathogenic strains. By using 14 mg of extract, the diameter of inhibition zones had a maximum of 16.7 mm vs. Salmonella typhimurium. Furthermore, Khoobchandani et al. (2010) described the antimicrobial activity of crude extracts of several parts of E. sativa against two Gram-positive and three Gram-negative bacteria. Particularly, a larger activity was stated for the seed oil against Gram-positive bacteria compared to Gram-negative bacteria. Moreover, Qaddoumi and El-banna (2019) outlined the antagonistic activity of E. sativa aqueous extract for E. coli and S. aureus. The corresponding φ values of the inhibition zones for E. coli and S. aureus were 19 mm and 12 mm, respectively. In the same study, antimicrobial activity of crude extract of ethyl acetate had no antimicrobial activity toward the tested pathogens.
In another study, Rizwana et al. (2016) investigated the antimicrobial potential of some organic extracts, viz. ethanol (EtOH), MeOH, ethyl acetate (EtOAc), acetate (AC), and CHCl3 of E. sativa, against Gram-positive and Gram-negative bacteria. Higher inhibition activity was determined in EtOAc and CHCl3 extracts against S. aureus (φ = 25 mm and 23 mm, respectively), followed by MeOH and EtOH (φ = 16 mm and 14 mm, respectively). The antimicrobial activity of solvent extracts of E. sativa (aerial parts and roots) and seed oil against antibiotic-resistant Gram-negative (E. coli, P. aeruginosa, and Shigella flexneri) and Gram-positive (S. aureus and B. subtilis) bacteria was investigated by Gulfraz et al. (2011). Among the various preparations, seed oil was the most active extract, displaying a zone of inhibition of 97% against Gram-positive bacteria and that of 74–97% against Gram-negative bacteria. The MIC of the seed oil was found as 65–75 μg/mL and 60–70 μg/mL for Gram-negative and Gram-positive bacteria, respectively (Khoobchandani et al., 2010). These authors assumed that antimicrobial activity of Eruca oil was mainly due to higher concentration of erucic acid, which was present in both free and triglyceride form. Awadelkareem et al. (2022b) reported that crude extract of E. sativa was active against food-borne pathogens, unveiling a rapid kinetics of killing bacteria in a time-dependent manner. The MIC and minimum bactericidal concentration (MBC) values of E. sativa crude extract ranged between 125 µg/mL and 500 µg/mL and between 250 µg/mL and 1,000 µg/mL, respectively. The inhibition of developed biofilm of E sativa was extended from 59% to 73% for all tested strains, and the E. sativa crude extract decreased the bacterial cells viability in biofilms.
Antioxidant activity
Ghazwani et al. (2020) established the aptitude of E. sativa decoction to inhibit hepatic LPO at a level of 150–400 μg/mL. Hence, a concentration of 400 μg/mL ensued 68.46% inhibition of oxidation of hepatic lipids, and the total antioxidant capacity of leaves was equal to 217 μg/mL (IC50). In addition, the in vitro design proposed the antioxidant mechanism of E. sativa by transfer of H atom and reduction of metal ions.
Barillari et al. (2005) reported that glucoerucin and its metabolite erucin are hydroperoxide scavenging antioxidants and assert direct antioxidant activity. The isolated fraction served as an active inducer of phase II enzymes, and it is highly operative to decrease oxidative stress and cell damage in various pathological conditions (Barillari et al., 2005).
With the aim of exploring variation in antioxidant potential of wild EER in connection to agricultural practices, Durazzo et al. (2013) reported that the ferric reducing antioxidant power (FRAP) ranged from 4.44 mmol/kg fresh weight (FW) to 9.92 mmol/kg FW for conventional EER and from 4.13 mmol/kg FW to 11.02 ± 0.45 mmol/kg FW for integrated EER. A study conducted by Heimler et al. (2007) observed a correlation between 2,2-Diphenyl-1-picrylhydrazyl (DPPH) activity and polyphenol content (R2 = 0.92 with GAE) in the case of five E. sativa varieties. Alam et al. (2007) studied the antioxidant impact of ethanolic extract of E. sativa seeds and its protecting effect on mercuric chloride (HgCl2)-induced renal toxicity. The corresponding extract was determined to comprise 4.5 μg/mg DW of glucoerucin and 6.5 μg/mg DW of flavonoids, thus attributing a significant antioxidant activity (IC50 ~60–65 μg/mL). The alcoholic extract of E. sativa seeds showed a powerful free radical scavenging and nephroprotective properties, since the HgCl2-inducing nephrotoxicity could induce oxidative stress. Alam et al. (2007) concluded that E. sativa seed extract inhibited HgCl2-induced LPO and nephrotoxicity. Interestingly, restoration of glutathione (GSH) levels by E. sativa seeds extracts also maintained glutathione reductase (GR) activity. In a mice model, a decrease in the activities of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), GR, and GSH and recovery to normal levels because of pretreatment with E. sativa seeds extract demonstrated that the oxidative stress caused by HgCl2 intoxication was neutralized due to the antioxidant activity of seeds extract.
Keyata et al. (2021) calculated the levels of bioactive compounds and antioxidant activities of Girgir leaves in Ethiopia. Results showed that the methanolic extract of leaves had a TFC of 14.03 mg of CE/g, TAC of 0.25 mg of cyanidin-3-glucoside/g, β-carotene = 0.36 mg/g, and L-ascorbic acid = 1.49 mg/g. Using DPPH and FRAP methods, the antioxidant activity of methanolic extract of Girgir leaves at 0.56 mg/mL was 80%. In addition, the extract had relatively low DPPH scavenging ability (71.70%) with IC50 = 0.15 mg/mL and a low FRAP (123.16 mM of Fe2+/g).
Awadelkareem et al. (2022a) reported the scavenging of radicals by E. sativa crude extract. The scavenging efficacy of crude extract with DPPH (IC50 = 66 µg/mL) radicals was more complete, compared to hydrogen peroxide (H2O2) (IC50 = 76 µg/mL) radicals. By applying total antioxidant capacity, reducing power, and H2O2, nitric oxide (NO), and SOD scavenging activity, Kishore et al. (2016) explored the antioxidant potential of alcohol and hydro-alcohol E. sativa extract. The total antioxidant capacity was 111.00 µM/g and 230.60 µM/g of ascorbic acid equivalent. Alcohol and hydro-alcohol extracts were found to scavenge DPPH and H2O2 radicals, NO, and SOD radicals. The IC50 values were found to be 3.28 and 3.53 µg/mL for DPPH, 188.11 and 181.56 µg/mL for H2O2, 73.05 and 64.33 µg/mL for NO, and 87.91 and 41.12 mg/mL for SOD radicals.
Findings of Koubaa et al. (2015) for antioxidant activities showed >90% DPPH free radical inhibition, 315 μg AAE/mL for 71 mg/mL extract, and >70% inhibition using β-carotene bleaching assay. The cited authors concluded that the main compounds of E. sativa leaf extract, viz. kaempferol 3,4-di-O-glucoside, kaempferol 3-glucosyl, quercetin 3-glucosyl, and isorhamnetin 3-glucosyl were linked to antioxidant potential. Owing to its high antioxidant activities, E. sativa extracts could be used as a food preservative or a nutraceutical in the food manufacturing or processing industry. In addition, E. sativa extracts could produce novel natural products for the food industry with safer and enhanced antioxidants with a potential for protection against oxidative damage in body as well as food.
Anticancer activity
By using 3-(4,5-dimethylthiazolyl-2)-2,5 diphenyltetrazolium bromide (MTT) assay, Awadelkareem et al. (2022a) studied the anticancer activity of E. sativa crude extract in two human colorectal cancer cells (HCT-116 and Caco-2). The extract could prevent the cell viability of both cells in a dose-dependent manner. The survival of HCT-116 cell line was found to be higher, and IC50 = 64 μg/mL, compared to CaCO2 with IC50 = 83 μg/mL.
Nazif et al. (2010) assessed the cytotoxic potential of the alcoholic extract of DSM, and isolated compounds to counter different tumor cell lines using sulforhodamine B (SRB) test. The activity was assessed against the following cell lines: colon carcinoma (HCT116), cervix carcinoma (Hela), liver carcinoma (HepG2), breast carcinoma (MCF7), and brain carcinoma (U251). Crude extract, glucoerucin, and glucoiberin showed cytotoxic activity for HCT116 at IC50 = 0.74, 2.42, and 0.94 μg/mL, respectively, while IC50 for Hela, HEPG2, MCF7, and U251 was >10 μg/mL.
Similarly, Michael et al. (2011) reported a powerful anticancer potential of 70% EtOH extract of E. sativa against the following cell lines: liver carcinoma (HepG2), breast carcinoma (MCF7), colon carcinoma (HCT116), and larynx carcinoma (Hep2). Hereto, several investigations established the effectiveness of erucin (1-isothiocyanato-4-(methylthio)butane) against various human cancer cell lines. AtHalf-maximal inhibitory concentration (IC50) equal to 28 µM, Azarenko et al. (2014) found that erucin stops proliferation of a breast cancer cell line (MCF7). In addition, cell cycle arrests mitosis at IC50 = 13 mM and apoptosis by a mechanism consistent with the impairment of microtubule dynamics. In the same direction, a study conducted by Singh et al. (2023) focused on the development of cubosomes loaded with erucin or ER-cubosomes (ER-CUB) isolated from seeds of E. sativa and evaluated their effectiveness for anti-proliferative activity against a colon cancer cell line Ehrlich-Ascites Carcinoma (EAC). The anticancer impact of erucin was described for dissimilar human cancer cell lines, such as the lung, liver, colon, and prostate, with the help of different mechanisms, such as cell cycle regulation, apoptosis, and inhibition of proliferation as well as mitochondrial depolarization. Generally, the mechanism of action showed the modulation of phase I, II, and III detoxifications, the regulation of cell growth by the induction of apoptosis, cell cycle arrest, the induction of reactive oxygen species (ROS) mechanism, and regulation of androgen receptor pathways (Dinkova-Kostova et al., 2017). Azarenko et al. (2014) found that erucin prevented the proliferation of MCF7 (breast cancer cells; IC50 = 28 mM) in parallel with cell cycle arrest at mitosis (IC50 = 13 mM) and apoptosis by a mechanism consistent with the impairment of microtubule dynamics.
Pure erucin and ER-CUB presented an efficient growth inhibition with the IC30 values of 0.0021 and 0.006 µL/mL, 0.016 and 0.062 µL/mL, and 0.023 and 0.084 µL/mL. According to Singh et al. (2023), ER-CUB cytotoxicity is principally due to the effect of erucin present in cubosomes, and the anticancer activity of erucin was reported on various cancerous cell lines; however, the effect of ER-CUB on EAC is not proved yet. ER-CUB revealed a substantial growth inhibitory outcome on tested cell line with IC50 = 0.0230 μL/mL. Erucin- and ER-CUB-treated cells showed morphological shifts, counting viability loss, chromatin aberrations, and loss of membrane integrity (Peng et al., 2010).
In a study conducted by Awadelkareem et al. (2022b), the anticancer effect of silver nanoparticles (Ag NPs) synthesized from E. sativa was assessed. In human lung cancer cells (A549), the anticancer potential of Ag NPs was examined by MTT, scratch, and invasion assays. The results indicated that Ag NPs inhibited the migration of A549 cells at 25.15 µg/mL. Singh et al. (2021) explored the anti-cancer effect of 4-(methylthio)butylisothiocyanate against 7,12-dimethylbenz[a]anthracene (DMBA)-induced breast cancer. Using reverse transcription-polymerase chain reaction (RT-PCR), hypoxia pathway was estimated and it was found that 40 mg/kg of 4-MTBITC depressed the expression of HIF-1α. Akt/mTOR signing pathway was one of the major pathways engaged in 4-MTBITC cell growth. Amino acid profiling of serum-free plasma discovered the down-regulation of specific amino acids required for vital mechanisms of fast-growing cancer cells. 4-MTBITC reduced the levels of Ser, Arg, Ala, Asn, and Glu. Isothiocyanate sulforaphane (SFN) is shown to prevent, directly or through a competitive mechanism, expression and activity of various phase I cytochrome P450 (CYP450) enzyme isoforms in mice and human tissues (Melchini and Traka, 2010), although phase I enzyme inhibition by erucin has not been demonstrated to date. Erucin (5–20 μM) did not inhibit CYP1A1 protein expression in HepG2 cells after coming in contact to human carcinogen benzo[a]pyrene (BaP, 50 μM), although erucin reduced the BaP-induced CYP1A1 activity in a dose-dependent manner, with a realization of 25% inhibition at the maximum studied level. Moreover, exposure of HepG2 cells to a concentration of 1-μM erucin decreased the BaP-induced DNA migration by 50%. Interestingly, one of the ITC constituents of E. sativa, identified as erysolin, showed stronger activity, compared to erucin, by inhibiting CYP1A1 activity in BaP-treated HepG2 cells by 50% at a lower concentration of 5 μM (Lamy et al., 2008).
Anti-obesity and hypoglycemic activities
Diabetes and obesity are the most dominant global health issues and their occurrence is cumulative at a high proportions, leading to huge economic burden. Obesity, a multifaceted illness, is accompanied by insulin resistance (IR) and increase in oxidative stress and inflammatory marker expression, leading to increased body fat mass. A number of investigations have confirmed the potential health benefits of E. sativa extracts in treating obesity and hypoglycemia. For instance, anti-obesity and hypoglycemic effects of dietary supplementation with E. sativa seeds extract on high fat (HF)-induced obesity was evaluated in mice by Piragine et al. (2021). Remarkably, in an experimental obesity model, these authors reported that seed extracts were able to reduce body weight, and therefore enhance glucose homeostasis. These findings were in agreement with the studies conducted with other Brassicaceae species. For example, a 4-week supplementation with Brassica rapa L. juice decreased cholesterol concentration in middle-aged men and enhanced the metabolism of cholesterol (Aiso et al., 2014). Lee et al. (2018) stated that Brassica juncea leaves extract supplementation had a positive effect on lipid profile and body fat in mice fed with HF diet. Certainly, a decrease in body fat aggregation and an amelioration of lipid profile were observed by modulating lipogenesis and cholesterol metabolism (Lee et al., 2018). It should be noted that main bioactive components in Brassicaceae vegetables are glucosinolates and their ITC derivatives produced by myrosinase enzyme (Palliyaguru et al., 2018).
Lately, glucosinolate glucoraphanin elevated inflammation and decreased insulin resistance linked with obesity. Nuclear factor erythroid 2-related factor 2 (Nrf2) was acknowledged as a main player in these beneficial effects, because the activation of Nrf2 endorsed the stimulation of both AMPK and uncoupling protein UCP1, which were altered in energy consumption (Xu et al., 2018).
Recently, hydrogen sulfide (H2S) has been considered a biological mediator in obesity. H2S-releasing goods of erucin have been established and abridged levels of H2S-synthesizing enzymes have been reported in fat tissues of obese mice and in genetically diabetic-obese (db/db) animals (Katsouda et al., 2018). According to Fuentes et al. (2014), E. sativa extracts displayed platelet activation inhibition, accumulation and release of inflammatory mediators, and a restriction of pro-inflammatory transcription factor NF-Κb. Lucarini et al. (2019) evaluated the anti-hyperalgesic impact of E. sativa DSM, along with its main glucosinolate, glucoerucin on diabetic neuropathic pain prompted in mice by streptozotocin (STZ). E. sativa DSM at 1 g/kg and glucoerucin at 100 µmol/kg presented a dose-dependent pain-relieving effect in STZ-diabetic mice. Co-administration with H2S scavengers abrogated the pain relief mediated by both E. sativa meal and glucoerucin. Their effect was also prevented by selectively blocking Kv7 K channels. Lucarini et al. (2019) concluded that erucin reduced both pain and neuro-inflammation linked with diabetes-induced neuropathic pain in mice.
In order to assess the antidiabetic potential of E. sativa fresh leaves extract, Hetta et al. (2017) investigated their in vitro potential for stimulation of glucose uptake and inhibition of G6Pase and adipogenic activities. Ethanolic extract (EC50 = 8.0 µg/mL) and its unsaponifiable fraction (EC50 = 5.8 µg/mL) could stimulate glucose uptake and inhibit G6Pase activity (ethanolic extract and its unsaponifiable fraction IC50 = 4.8 µg/mL and 9.3 µg/mL, respectively) and substantial adipogenic activities (ethanolic extract and its unsaponifiable fraction EC50 = ≈4.3 µg/mL and 6.1 µg/mL, respectively).
E. sativa extracts regulate lipid metabolism and improve insulin resistance to reduce lipotoxicity. Next, E. sativa extracts enhance insulin signaling and reinstate equilibrium between glucose production and utilization. Lastly, E. sativa extracts renovate imbalance in autophagy-apoptosis to protect β cells.
Cardiovascular benefits
Testai et al. (2022) studied in vitro hypertensive mice for E. sativa DSM extract and it stimulating hypotensive effect. In addition, these authors suggested dose-dependent cardio-protection in vivo model of acute myocardial infarct, which attained a reversible coronary occlusion. This latter effect was sensitive to blockers of mitochondrial ATP-sensitive potassium (KATP) and Kv7.4K channels, signifying a probable role of these mitochondrial channels in the protective effects of DSM extract. Accordingly, DSM extracts condensed calcium (Ca) uptake and apoptotic cell death in isolated cardiac mitochondria.
Alotaibi et al. (2020) examined the therapeutic role of rocket seeds (RS) against hydroxyapatite nanoparticles (HAP NPs) injection in mice that caused cardiac injury by diminishing oxidative stress and apoptosis. Regarding the HAP NPs+RS sample, a significant reduction of cardiac markers, viz. creatine phosphokinase (CPK), creatine phosphokinase-myocardial band (CPK-MB), lactate dehydrogenase (LDH), and myoglobin, was obtained. In addition, treated rats with RS extract after HAP NPs injection displayed a rise in cardiac SOD and CAT levels, compared to HAP NPs injection only. More recently, Flori et al. (2023) showed that erucin exerts cardioprotective effects against ischemia-reperfusion (I/R) damage through the involvement of mitochondrial KATP (mitoKATP) channels. Salma et al. (2018) also stated that E. sativa is an antihypertensive remedy mainly because of its vasodilatory and partly cardiac effects. Although the supposed mechanisms of action through which H2S can have cardioprotective impact are abundant and involve antioxidant transcription factors (such as Nrf2), anti-inflammatory cytokines, and other agents, such as nitric oxide (NO), mitoK channels are documented as a stimulating target. Noteworthy is the fact that the cardioprotection promoted by sodium hydrosulfide (NaHS) in a model of myocardial I/R injury was meaningfully upturned by pharmacological blockage of mitoKATP channels (Testai et al., 2022). The involvement of mitoKATP channels in the cardioprotective effects of H2S was demonstrated by using a synthetic H2S-donor isothiocyanate (). Noticeably, sulforaphane at micromolar concentrations preserved both cultured cardiomyoblasts and adult cardiomyocytes from H2O2-induced oxidative stress through the modulation of Nrf2 signaling pathway (Testai et al., 2022).
Neuroprotective effect
In an in vivo design following addition of gentamicin, Abdelkader et al. (2022) examined the nephroprotective effect of E. sativa seeds aqueous extract. The authors reported that 150 mg/kg of the studied extract could preserve the kidneys from gentamicin-induced nephrotoxicity, and reduce oxidative damage induced by the antibiotic. Additionally, double dose of E. sativa seeds extract (300 mg/kg) provoked an increase in nitric oxide at kidney level in gentamicin-nephrotoxic animals (Abdelkader et al., 2022). Remarkably, E. sativa extract at both low and high doses, could reduce the inflammatory cascade activated by gentamicin after nephrotoxicity induction, thereby activating reduction of tumor necrosis factor-α (TNF-α) and interleukin 1β (IL-1β) (Abdelkader et al., 2022). Moreover, an in vitro study described dampening of lipopolysaccharide (LPS)-induced neuro-inflammation, with a decrease in pro-inflammatory cytokines, elimination of cyclooxygenase-2 (COX-2), and amended some expressions in cells pretreated with erucin extract (Gugliandolo et al., 2018). In this study, the NSC-34 motor neurons liable to the culture medium of LPS-stimulated macrophages, E. sativa (E. sativa, Mill.) extract could prevent the LPS-induced cell death and disintegration by neutralizing apoptosis. Interestingly, inhibition of Fas ligand (FasL) expression represses pro-inflammatory mediators (via COX-2 and toll-like receptor 4 [TLR4] and TNF-α) and excites the release of cytokine IL-10 (Gugliandolo et al., 2018). Recent studies have shown the multiple neuroprotective mechanisms of ITCs—major compounds found in E. sativa extracts. In this way, the electrophilic interaction of ITCs with the cysteine residues of the cytoplasmatic Kelchlike ECH-associated protein 1 (Keap1)−Nrf2 complex is a central event to endorse the binding of Nrf2 with antioxidant responsive element (ARE) at nuclear level (Morroni et al., 2018). On the other hand, SFN, derived from precursor glucosinolate present in E. sativa, and, after uptake in organism, is conjugated with GSH and metabolized through the mercapturic acid path to its corresponding mercapturic acid derivate SFN-cysteinylglycine, SFN-cysteine, and SFN-Nacetylcysteine. One interesting aspect is the reduction of sulfoxide SFN to its thioether analogue erucin (i.e. 4-methylthiobutyl ITC erucin), which is metabolized through mercapturic acid pathway and excreted in urine or bile (Morroni et al., 2018).
Clinical trials and the main outcomes of E. sativa bioactive compounds
In a preclinical study conducted by Alqasoumi et al. (2009), E. sativa extract could expressively decrease urease activity; also, gastric ulcers prompted by necrotizing agents, such as indomethacin and hypothermic agent, were also reduced, as established histologically.
In a preclinical study, the aphrodisiac potential of E. sativa extract was verified. Ethanolic extract of E. sativa extract displayed androgenic action and induced testicular steroids production that stimulated the pituitary gland and improved spermatogenesis in male mice testes (Grami et al., 2024). The cited authors reported that adapted sperm parameters existing in diabetic mouse model, which was induced by streptozotocin exposure, were significantly improved by the administration of 250- and 500-mg/kg ethanol extract for 8 consecutive weeks. Compared to untreated diabetic animals, weight of the testes, epididymis, seminal vesicles, and prostate was suggestively augmented in treated mice at the end of the treatment (Grami et al., 2024).
Extract of E. sativa leaves is considered for the rise of testosterone level and increased sperm activity as well as it reduced sperm death and its abnormalities (Hadi, 2017). E. sativa extract contained saponins and alkaloids and could develop sperm activity. Its histological examination showed a significant increase in the diameter of its tubules, spermatids, and Leydig cells as well as reduction in interstitial space. EER extract increases the growth of testis and augments proliferation and maturation of spermatozoa. E. sativa seeds oil also reduces nicotine-induced testicular damage by morphometric and histological modification (Grami et al., 2024). This impact was attributable to desulfo-glucosinolates, erucic acid, and ITCs. The study conducted by Abd El-Aziz et al. (2016) exposed that minor doses of seeds oil stimulate spermatogenesis.
Regarding nephroprotective effect, a preclinical study conducted by Elgazar and Abo Raya (2013) showed that alcoholic extract of E. sativa seeds had an impact on HgCl2-induced nephrotoxicity. This effect was due to antioxidant molecules, enzymes, flavonoids, and glucoerucin constituents that had an important role in nephroprotection. Hussein et al. (2018) evidenced that aqueous E. sativa extract significantly reduced the occurrence of nephrocalcinosis by halting calcium oxalate crystal formation and its deposition in renal tissues by diuresis and alkalization. E. sativa extract also reduced the incidence of calcium oxalate kidney stones because of the presence of large amounts of magnesium, which reduced the binding of oxalate to calcium ions in renal tubules.
A preclinical study demonstrated that oral and intravenous administration of methanolic E. sativa extract had hypotensive effects on hypertensive and normotensive subjects. This action was due to vasodilatory and cardiotonic effects. Vasodilation action of E. sativa extract across vasodilatory mediators (nitric oxide and certain muscarinic receptors), which are presents on vascular endothelial cells, has a direct effect on vascular smooth muscle (Alqasoumi et al., 2009).
Potential Commercial Applications of E. sativa
Traditionally, the plant was used to treat hypertension and diabetes and as a rubefacient, tonic, and diuretic, digestive, astringent, laxative, emollient, stimulant, stomachic and scurvy agent (Jaafar and Jaafar, 2019; Salma et al., 2018). The airy tender fresh parts of E. sativa were used in salad and occasionally cooked as a potherb (Khoobchandani et al., 2010;). In addition, an antibacterial cream was also commercialized (Sanad et al., 2016).
Of all the research papers concerning E. sativa and its phytochemistry, none has discussed how details could be employed within a population. A few plants breeding programmes were established by virtue of the number of environmental factors affecting E. sativa growth, development, and reproduction. For instance, selection of E. sativa plants through conventional/molecular breeding could be a valuable tool for research community and provide an excellent incentive for breeding companies to fund research on pharmaceuticals and supplements. The authentic checking and quantification of characteristics of E. sativa compounds would not only validate the heritability of such traits in EER but also provide a “roadmap” for how other minor crops could be developed for commercial use. Attentiveness should be rendered to the phytochemical content of E. sativa. Utilizing genetic resources, the falling costs of sequencing and bioinformatics can soon produce nutritive superior varieties of E. sativa. Plant breeding typically takes long time than the average research project, even with the use of advanced genomic selection methods. This situation could be remedied by long-term industrial collaboration and sponsorship by plant-breeding firms.
There are still unanswered questions related to standardizing of E. sativa extracts formulations. In view of variations involved in harvesting, extraction, and product formulation methods, variabilities in quality, efficacy, and composition of E. sativa compounds are maintained. In addition, in spite of potential of compounds extracted from E. sativa, commercialization of new pharmaceuticals and supplements is generally trailing and a large volume of scientific findings remain unutilized. Standardization of active E. sativa compounds therefore seems to be one of the missing links to ensure consistency in efficacy. Crude plant products or semi-refined mixtures seem to provide consistent efficacy. However, application of crude plant materials may only offer a short-term solution for small-scale farming, as production for use on industrial scale may not be economically feasible in terms of production costs and space. Therefore, improved novel formulation methods that can preserve the chemical composition of analogues and compartmentalization of E. sativa active compounds are required to minimize environmental concerns, and improve their performance and persistence in soil.
Long-term Studies to Evaluate the Efficacy and Safety of E. sativa Metabolites Over Extended Periods
Long-term safety of E. sativa leaf ethanolic extract was approved in male mice. Investigations showed that ethanolic extract is not toxic to the structure and functioning of the liver and kidneys when administered to mice orally at a specific dose. The extract rich in antioxidants protect the cells from damage (El-Gayar et al., 2022). Moreover, it has been reported that low dose of E. sativa seed oil was not associated with teratogenic changes; however, high doses induced some abnormalities (Moustafa and El-Makawy, 2002). Figure 6 outlines the medicinal potential of different parts of E. sativa.
Figure 6. Medicinal potential of E. sativa.
Industrial-scale production of E. sativa seeds oil
The usage of inedible oils from natural plants as feedstock is required. Therefore, E. sativa crude oil is viewed as a substitute to mineral oils in many industries, and it has good potential for biodiesel production because of its good stability and high productivity at room temperature. It emerges that seeds of E. sativa Gars (EGS) contain 35% oil, which is suitable for production of biodiesel. As an illustration, the price of ESG oil in northwest of China is about $0.15 per kg, much cheaper than soybean oil. Therefore, ESG oil is an excellent option for producing biodiesel. Li et al. (2009) produced ESG biodiesel, a type of nontoxic, biodegradable, and renewable alternative fuel. These authors generated a high-quality biodiesel from low-cost ESG oil in laboratory. The transesterification of ESG oil was catalyzed by solid heteropoly acid, Cs2.5H0.5PW12O40. Remarkably, the characteristics of biodiesel from ESG were comparable to conventional diesel and comply to the US Standard for biodiesel (ASTM 6751). In addition, harmful emissions, such as HC and CO, were reduced by 33.33%. However, an increase in energy consumption and CO2 and NOx emissions was observed by about 10.15%, 10.71%, and 13.21%, respectively.
On the other hand, Aghababaie et al. (2019) produced E. sativa oil in a two-phase enzymatic membrane bioreactor (TP-EMR). The yield of biodiesel production from crude oil using free Candida rugosa lipase (CRL) was augmented by TP-EMR. With the application of TP-EMR, the yield of biodiesel from non-edible E. sativa oil using CRL was complete: 100% yield was achieved in TP-EMR with commercial polyacrylonitrile (PAN) and organic phase flow rate of 40 mL/min along with 40% initial water content in organic phase.
Conclusions
Here, some important background summarized the actual status of E. sativa exploration concentrating their bioactive compounds. These compounds matched in their usage in medicine, pharmaceutical industries, and as nutraceuticals because of their antioxidant and antibacterial properties. Glucosinolates and their derivatives, flavonoid fractions, soluble and insoluble phenolics, and fatty acids of E. sativa are the main bioactive compounds that merit care for their biological activities. In this review, the isolated chemical components of Eruca from different parts and identified by MS were outlined. Additional tool sophistication coupled with numerous systems, such as complex chromatography with NMR and MS in series, is imperative. In this line, an augmented importance of microcapillary columns with nanotechnology ESI systems appears certain. Moreover, biopharmacological effects are well reviewed. The link between Eruca natural products structure and their biological activity requires examination. Hence, the elucidation of mechanisms of action of Eruca phytochemicals could guide for its clinical application. On the other hand, although E. spp. extracts are generally recognized as safe, further examination is required to control and regulate their usage. Consequently, toxicological tests should be performed to assess its safe edible usage.
Modern methods, counting metabolomics, in silico insights, and nanotechnology have become essential tools for further progress in knowledge. In this regard, these modern tools aid to recognize therapeutics of E. sativa botanically and understand their metabolism. It is also conceivable to control detrimental E. sativa compounds and to estimate appropriate dosages. Furthermore, the introduction of novel techniques would aid in the recognition of novel compounds as well as in identifying their targets and understanding their mode of action.
Acknowledgments
The researcher thanks the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support (QU-APC-2024-9/1).
Conflict of Interest
The author declared no conflict of interest.
REFERENCES
Abd El-Aziz, G.S., El-Fark, M.O., and Hamdy, R.M., 2016. Protective effect of Eruca sativa seed oil against oral nicotine induced testicular damage in rats. Tissue and Cell 48(4): 340–348. 10.1016/j.tice.2016.05.006
Abdelkader, R.S.E., El-Beih, N.M., Zaahkouk, S.A., and El-Hussieny, E.A., 2022. Ameliorative effect of E. sativa seeds and its rutin on gentamicin-induced nephrotoxicity in male rats via targeting inflammatory status, oxidative stress and kidney injury molecule-1 (KIM-1)/cystatin c expression. Indonesian Biomedical Journal 14(1): 74–83. 10.18585/inabj.v14i1.1766
Abd-Elsalam, R.M., El Badawy, S.A., Ogaly, H.A., Ibrahim, F.M., Farag, O.M., and Ahmed, K.A., 2021. Eruca sativa seed extract modulates oxidative stress and apoptosis and up-regulates the expression of Bcl-2 and Bax genes in acrylamide-induced testicular dysfunction in rats. Environmental Science and Pollution Research 28(38): 53249–53266. 10.1007/s11356-021-14532-y
Aghababaie, M., Beheshti, M., Razmjou, A., and Bordbar, A.K., 2019. Two-phase enzymatic membrane reactor for the production of biodiesel from crude Eruca sativa oil. Renewable Energy 140: 104–110. 10.1016/j.renene.2019.03.069
Ahmed, S.K., Hussein, S., Qurbani, K., Ibrahim, R.H., Fareeq, A., Mahmood, K.A., and Mohamed, M.G. 2024. Antimicrobial resistance: impacts, challenges, and future prospects. Journal of Medicine, Surgery, and Public Health 2: 100081. 10.1016/j.glmedi.2024.100081
Alam, M.S., Kaur, G., Jabbar, Z., Javed, K., and Athar, M., 2007. Eruca sativa seeds possess antioxidant activity and exert a protective effect on mercuric chloride induced renal toxicity. Food and Chemical toxicology 45(6): 910–920. 10.1016/j.fct.2006.11.013
Alqasoumi, S., Al-Sohaibani, M., Al-Howiriny, T., Al-Yahya, M., and Rafatullah, S., 2009. Rocket “Eruca sativa”: a salad herb with potential gastric antiulcer activity. World Journal of Gastroenterology: WJG 15(16): 1958. 10.3748/wjg.15.1958
Alotaibi, B., El-Masry, T.A., Tousson, E., Alarfaj, S.J., and Saleh, A. 2020. Therapeutic effect of rocket seeds (Eruca sativa L.) against hydroxyapatite nanoparticles injection induced cardiac toxicity in rats. Pakistan Journal of Pharmaceutical Science 33(4): 1839–1845.
Aiso, I., Inoue, H., Seiyama, Y., and Kuwano, T., 2014. Compared with the intake of commercial vegetable juice, the intake of fresh fruit and komatsuna (Brassica rapa L. var. perviridis) juice mixture reduces serum cholesterol in middle-aged men: a randomized controlled pilot study. Lipids in Health and Disease 13: 1–8. 10.1186/1476-511X-13-102
Awadelkareem, A.M., Al-Shammari, E., Elkhalifa, A.E.O., Adnan, M., Siddiqui, A.J., Snoussi, M., et al. 2022a. Phytochemical and in silico ADME/Tox analysis of Eruca sativa extract with antioxidant, antibacterial and anticancer potential against CaCO-2 and HCT-116 colorectal carcinoma cell lines. Molecules 27(4): 1409. 10.3390/molecules27041409
Awadelkareem, A.M., Al-Shammari, E., Elkhalifa, A.O., Adnan, M., Siddiqui, A. J., Mahmood, D., et al. 2022b. Anti-adhesion and antibiofilm activity of Eruca sativa miller extract targeting cell adhesion proteins of food-borne bacteria as a potential mechanism: combined in vitro-in silico approach. Plants 11(5): 610. 10.3390/plants11050610
Azarenko, O., Jordan, M.A., and Wilson, L., 2014. Erucin, the major isothiocyanate in arugula (Eruca sativa), inhibits proliferation of MCF7 tumor cells by suppressing microtubule dynamics. PloS One 9(6): e100599. 10.1371/journal.pone.0100599
Badraoui, R., Rebai, T., Elkahoui, S., Alreshidi, M., N. Veettil, V., Noumi, E., A. Al-Motair, K.; Aouadi, K., Kadri, A., De Feo, V., et al., 2020. Allium subhirsutum L. as a potential source of antioxidant and anticancer bioactive molecules: HR-LCMS phytochemical profiling, in vitro and in vivo pharmacological study. Antioxidants 9: 1003. 10.3390/antiox9101003
Barillari, J., Canistro, D., Paolini, M., Ferroni, F., Pedulli, G. F., Iori, R., and Valgimigli, L., 2005. Direct antioxidant activity of purified glucoerucin, the dietary secondary metabolite contained in rocket (Eruca sativa Mill.) seeds and sprouts. Journal of Agricultural and Food Chemistry 53(7): 2475–2482. 10.1021/jf047945a
Bell, L., Chadwick, M., Puranik, M., Tudor, R., Methven, L., Kennedy, S., and Wagstaff, C., 2020. The Eruca sativa genome and transcriptome: a targeted analysis of sulfur metabolism and glucosinolate biosynthesis pre and postharvest. Frontiers in Plant Science 11: 525102. 10.3389/fpls.2020.525102
Bell, L., Oruna-Concha, M.J., and Wagstaff, C., 2015. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC–MS: highlighting the potential for improving nutritional value of rocket crops. Food Chemistry 172: 852–861. 10.1016/j.foodchem.2014.09.116
Bobate, S., Mahalle, S., Dafale, N.A., and Bajaj, A. 2023. Emergence of environmental antibiotic resistance: mechanism, monitoring and management. Environmental Advances 13: 100409. 10.1016/j.envadv.2023.100409
Bouanani, S., Zeggar, M., and Lecouvey, M., 2023. Isolation, characterization, and valorization of hemicelluloses from olive solid residue as biomaterial, partial kaolin hydrolysis, and antiproliferative activity. Biomass Conversion and Biorefinery, 1–17. 10.1007/s13399-023-03962-y
Buitrago-Villanueva, I., Barbosa-Cornelio, R., and Coy-Barrera, E., 2023. Influence of the culture system and harvest time on the specialized metabolite composition of rocket salad (Eruca sativa) leaves. Horticulturae 9(2): 235. 10.3390/horticulturae9020235
Cangui-Panchi, S.P., Ñacato-Toapanta, A.L., Enríquez-Martínez, L.J., Salinas-Delgado, G.A., Reyes, J., Garzon-Chavez, D., and Machado, A., 2023. Battle royale: immune response on biofilms–host-pathogen interactions. Current Research in Immunology 4: 100057. 10.1016/j.crimmu.2023.100057
Cataldi, T.R., Rubino, A., Lelario, F., and Bufo, S.A., 2007. Naturally occurring glucosinolates in plant extracts of rocket salad (Eruca sativa L.) identified by liquid chromatography coupled with negative ion electrospray ionization and quadrupole ion-trap mass spectrometry. Rapid Communications in Mass Spectrometry 21(14): 2374–2388. 10.1002/rcm.3101
Chatterjee, S., and Haque, E., 2024. Pathogenic microbial genetic diversity with reference to significant medical and public health. In: Microbial diversity in the genomic era. Academic Press, pp. 561–587. 10.1016/B978-0-443-13320-6.00038-X
Cheke, R.S., Patil, V.M., Firke, S.D., Ambhore, J.P., Ansari, I.A., Patel, H.M., Shinde, S.D., Pasupuleti, V.R., Hassan, M.I., Adnan, M., et al., 2022. Therapeutic outcomes of isatin and its derivatives against multiple diseases: recent developments in drug discovery. Pharmaceuticals 15: 272. 10.3390/ph15030272
Crescenzi, M.A., Cerulli, A., Montoro, P., and Piacente, S., 2023. Metabolite profiling for typization of “rucola della piana del sele” (PGI), Eruca sativa, through UHPLC-Q-Exactive-Orbitrap-MS/MS analysis. Foods 12(18): 3384. 10.3390/foods12183384
Dernovics, M., Molnár, A., and Szalai, G., 2023. UV-B radiation-induced di-and polysulfide derivatives of 4-mercaptobutyl glucosinolate from Eruca sativa. Journal of Food Composition and Analysis 122: 105485. 10.1016/j.jfca.2023.105485
Dinkova-Kostova, A.T., Fahey, J.W., Kostov, R.V., and Kensler, T.W., 2017. KEAP1 and done? Targeting the NRF2 pathway with sulforaphane. Trends in Food Science & Technology 69(Pt. B): 257–269. 10.1016/j.tifs.2017.02.002
Dong, J., Ma, X., Wei, Q., Peng, S., and Zhang, S., 2011. Effects of growing location on the contents of secondary metabolites in the leaves of four selected superior clones of Eucommia ulmoides. Industrial Crops and Products 34(3): 1607–1614. 10.1016/j.indcrop.2011.06.007
Durazzo, A., Azzini, E., Lazzè, M.C., Raguzzini, A., Pizzala, R., and Maiani, G., 2013. Italian wild rocket [Diplotaxis tenuifolia (L.) DC.]: influence of agricultural practices on antioxidant molecules and on cytotoxicity and antiproliferative effects. Agriculture 3(2): 285–298. 10.3390/agriculture3020285
El-Gayar, H.A., Salem, E.T., and Edrees, G.M., 2022. The protective effect of Eruca sativa against lipid metabolic abnormalities induced by dioxin in male rats. Egyptian Journal of Basic and Applied Sciences, 9(1): 383–392. 10.1080/2314808X.2022.2102772
Elgazar, F.A., and AboRaya, A. O. (2013). Nephroprotective and diuretic effects of three medicinal herbs against gentamicin-induced nephrotoxicity in male rats. Pakistan Journal of Nutrition, 12(8), 715–722. 10.3923/pjn.2013.715.722
El-Nwehy, S.S., Abd-Rabbu, H.S., El-Nasharty, A.B., and Rezk, A.I., 2023. Improvement of (Eruca sativa Mill) yield, oil, chemical constituents and antioxidant activity utilizing a by-product of yeast production (CMS) with zinc and boron under salinity stress conditions. Oil Crop Science 8(4): 218–227. 10.1016/j.ocsci.2023.09.007
Fagerlund, A., Wubshet, S.G., Møretrø, T., Schmidt, G., Borge, G. I., and Langsrud, S., 2021. Anti-listerial properties of chemical constituents of Eruca sativa (rocket salad): from industrial observation to in vitro activity. Plos One 16(4): e0250648. 10.1371/journal.pone.0250648
Fechner, J., Kaufmann, M., Herz, C., Eisenschmidt, D., Lamy, E., Kroh, L.W., and Hanschen, F.S., 2018. The major glucosinolate hydrolysis product in rocket (Eruca sativa L.), sativin, is 1, 3-thiazepane-2-thione: elucidation of structure, bioactivity, and stability compared to other rocket isothiocyanates. Food Chemistry 261: 57–65. 10.1016/j.foodchem.2018.04.023
Flori, L., Montanaro, R., Pagnotta, E., Ugolini, L., Righetti, L., Martelli, A., et al. (2023). Erucin exerts cardioprotective effects on ischemia/reperfusion injury through the modulation of mitoKATP channels. Biomedicines 11(12): 3281. 10.3390/biomedicines11123281
Franco, P., Spinozzi, S., Pagnotta, E., Lazzeri, L., Ugolini, L., Camborata, C., and Roda, A., 2016. Development of a liquid chromatography–electrospray ionization–tandem mass spectrometry method for the simultaneous analysis of intact glucosinolates and isothiocyanates in Brassicaceae seeds and functional foods. Journal of Chromatography A 1428: 154–161. 10.1016/j.chroma.2015.09.001
Fuentes, E., Alarcón, M., Fuentes, M., Carrasco, G., and Palomo, I., 2014. A novel role of Eruca sativa Mill. (rocket) extract: antiplatelet (NF-κB inhibition) and antithrombotic activities. Nutrients 6(12): 5839–5852. 10.3390/nu6125839
Ghazwani, S.M.S., Alhazmi, H.A., Najmi, A., Ashraf, S.E., and Rehman, Z., 2020. Establishing gerger (eruca sativa) leaves as functional food by GC-MS and in-vitro anti-lipid peroxidation assays. Journal of Food and Nutrition Research 8(8): 441–449. 10.12691/jfnr-8-8-8
Ghasemi, K., Ghasemi, Y., Ehteshamnia, A., Nabavi, S.M., Nabavi, S.F., Ebrahimzadeh, M.A., and Pourmorad, F., 2011. Influence of environmental factors on antioxidant activity, phenol and flavonoids contents of walnut (Juglans regia L.) green husks. Journal of Medicinal Plants Research 5(7): 1128–1133.
Grami, D., Selmi, S., Rtibi, K., Sebai, H., and De Toni, L. 2024. Emerging role of Eruca sativa Mill. in male reproductive health. Nutrients 16(2): 253. 10.3390/nu16020253
Gugliandolo, A., Giacoppo, S., Ficicchia, M., Aliquò, A., Bramanti, P., and Mazzon, E., 2018. Eruca sativa seed extract: a novel natural product able to counteract neuroinflammation. Molecular Medicine Reports 17(5): 6235–6244. 10.3892/mmr.2018.8695
Gulfraz, M., Sadiq, A., Tariq, H., Imran, M., Qureshi, R., and Zeenat, A. 2011. Phytochemical analysis and antibacterial activity of Eruca sativa seed. Pakistan Journal of Botany 43(2): 1351–1359.
Hadi, M.A., Almamoori, A.M., Al-Hassnawi, A.T., and Hameedi, E.H. 2017. Oxidative response associated with treatment of male Albino rats with Eruca sativa Mill leaves extract and correlations with complete blood picture. Journal of Pharmaceutical Sciences and Research 9(11): 2278–2285.
Heimler, D., Isolani, L., Vignolini, P., Tombelli, S., and Romani, A. 2007. Polyphenol content and antioxidative activity in some species of freshly consumed salads. Journal of Agricultural and Food Chemistry 55(5): 1724–1729. 10.1021/jf0628983
Herr, I., and Büchler, M.W., 2010. Dietary constituents of broccoli and other cruciferous vegetables: implications for prevention and therapy of cancer. Cancer Treatment Reviews 36(5): 377–383. 10.1016/j.ctrv.2010.01.002
Hetta, M.H., Owis, A.I., Haddad, P.S., and Eid, H.M., 2017. The fatty acid-rich fraction of Eruca sativa (rocket salad) leaf extract exerts antidiabetic effects in cultured skeletal muscle, adipocytes and liver cells. Pharmaceutical Biology 55(1): 810–818. 10.1080/13880209.2017.1280687
Jaafar, N.S., and Jaafar, I.S., 2019. Eruca sativa Linn.: pharmacognostical and pharmacological properties and pharmaceutical preparations. Asian Journal of Pharmaceutics and Clinical Research 12(3): 39–45. 10.22159/ajpcr.2019.v12i3.30893
Kaczorová, D., Karalija, E., Dahija, S., Bešta-Gajević, R., Parić, A., and Ćavar Zeljković, S. 2021. Influence of extraction solvent on the phenolic profile and bioactivity of two achillea species. Molecules 26: 1601. 10.3390/molecules26061601
Katsouda, A., Szabo, C., and Papapetropoulos, A., 2018. Reduced adipose tissue H2S in obesity. Pharmacological Research 128: 190–199. 10.1016/j.phrs.2017.09.023
Keyata, E.O., Tola, Y.B., Bultosa, G., and Forsido, S.F., 2021. Phytochemical contents, antioxidant activity and functional properties of Raphanus sativus L, Eruca sativa L., and Hibiscus sabdariffa L. growing in Ethiopia. Heliyon 7(1): e05939. 10.1016/j.heliyon.2021.e05939
Khan, H., and Khan, M.A., 2014. Antiulcer effect of extract/fractions of Eruca sativa: attenuation of urease activity. Journal of Evidence-Based Complementary & Alternative Medicine 19(3): 176–180. 10.1177/2156587214527452
Koubaa, M., Driss, D., Bouaziz, F., Ghorbel, R.E., and Chaabouni, S.E., 2015. Antioxidant and antimicrobial activities of solvent extract obtained from rocket (Eruca sativa L.) flowers. Free Radicals and Antioxidants 5(1): 29–34. 10.5530/fra.2015.1.5
Kim, N., 2024. Synopsis of H. pylori-associated diseases. In: Helicobacter pylori. Springer Nature, Singapore, pp. 217–228. 10.1007/978-981-97-0013-4_17
Kim, S.J., and Ishii, G., 2006. Glucosinolate profiles in the seeds, leaves and roots of rocket salad (Eruca sativa Mill.) and anti-oxidative activities of intact plant powder and purified 4-methoxyglucobrassicin. Soil Science and Plant Nutrition 52(3): 394–400. 10.1111/j.1747-0765.2006.00049.x
Kishore, L., Kaur, N., and Singh, R., 2016. Evaluation of antioxidant activity and total phenolic content of Eruca sativa L. Seeds. International Journal of Toxicological and Pharmacological Research 8: 146–51.
Khaliq, B., Falke, S., Saeed, Q., Bilal, M., Munawar, A., Ali, A., et al. 2021. Eruca sativa seed napin structural insights and thorough functional characterization. Scientific Reports 11(1): 24066. 10.1038/s41598-021-02174-6
Khoobchandani, M., Ganesh, N., Gabbanini, S., Valgimigli, L., and Srivastava, M.M. 2011. Phytochemical potential of Eruca sativa for inhibition of melanoma tumor growth. Fitoterapia 82(4): 647–653. 10.1016/j.fitote.2011.02.004
Khoobchandani, M., Ojeswi, B.K., Ganesh, N., Srivastava, M.M., Gabbanini, S., Matera, R., et al. 2010. Antimicrobial properties and analytical profile of traditional Eruca sativa seed oil: comparison with various aerial and root plant extracts. Food Chemistry 120(1): 217–224. 10.1016/j.foodchem.2009.10.011
Kumar, N., Kaur, B., Shukla, S., Patel, M.K., Thakur, M.S., Kumar, R., et al. 2022. Comparative analysis of phytochemical composition and anti-oxidant and anti-inflammatory benefits of Eruca sativa grown at high altitude than at lower altitude. Chemical Papers 76(12): 7759–7782. 10.1007/s11696-022-02418-3
Lamy, E., Schröder, J., Paulus, S., Brenk, P., Stahl, T., and Mersch-Sundermann, V. 2008. Antigenotoxic properties of Eruca sativa (rocket plant), erucin and erysolin in human hepatoma (HepG2) cells towards benzo (a) pyrene and their mode of action. Food and Chemical Toxicology 46(7): 2415–2421. 10.1016/j.fct.2008.03.022
Lee, J.J., Kim, H.A., and Lee, J., 2018. The effects of Brassica juncea L. leaf extract on obesity and lipid profiles of rats fed a high-fat/high-cholesterol diet. Nutrition Research and Practice 12(4): 298. 10.4162/nrp.2018.12.4.298
Li, J., Peng, F., Huang, H., Xu, X., Guan, Q., Xie, M., and Xiong, T., 2024. Characterization, mechanism and in vivo validation of Helicobacter pylori antagonism by probiotics screened from infants’ feces and oral cavity. Food & Function 15(3): 1170–1190. 10.1039/D3FO04592G
Li, S., Wang, Y., Dong, S., Chen, Y., Cao, F., Chai, F., and Wang, X. 2009. Biodiesel production from Eruca Sativa Gars vegetable oil and motor, emissions properties. Renewable Energy 34(7): 1871–1876. 10.1016/j.renene.2008.12.020
Lucarini, E., Pagnotta, E., Micheli, L., Parisio, C., Testai, L., Martelli, A., et al. 2019. Eruca sativa meal against diabetic neuropathic pain: an H2S-mediated effect of glucoerucin. Molecules 24(16): 3006. 10.3390/molecules24163006
Madia, F., Worth, A., Whelan, M., and Corvi, R., 2019. Carcinogenicity assessment: addressing the challenges of cancer and chemicals in the environment. Environment International 128: 417–429. 10.1016/j.envint.2019.04.067
Madjo, K.K.Y., Mbiantcha, M., Fagni, L.Z.N., Matah Marthe, V.M., Ngoufack, E.A., Tsafack, G.E., et al. 2023. Gastroprotective and ulcer healing effects of Nauclea pobeguinii (Rubiaceae) on experimentally induced gastric ulcers in male Wistar rats. Journal of Complementary and Integrative Medicine 20(4): 729–739. 10.1515/jcim-2023-0024
Malik, S.N. 2015. Antibacterial activity of olive (Olea europaea) leaves and arugula (Eruca sativa) seeds extract. Int. J. Pharmacogn. Phytochem. Res 7(2): 307–310.
Martínez-Sánchez, A., Llorach, R., Gil, M. I., and Ferreres, F. 2007. Identification of new flavonoid glycosides and flavonoid profiles to characterize rocket leafy salads (Eruca vesicaria and Diplotaxis tenuifolia). Journal of Agricultural and Food Chemistry 55(4): 1356–1363. 10.1021/jf063474b
Maycotte, P., Illanes, M., and Moreno, D.A. 2024. Glucosinolates, isothiocyanates, and their role in the regulation of autophagy and cellular function. Phytochemistry Reviews 1–35. 10.1007/s11101-024-09944-w
Melchini, A., and Traka, M.H. 2010. Biological profile of erucin: a new promising anticancer agent from cruciferous vegetables. Toxins 2(4): 593–612. 10.3390/toxins2040593
Michael, H.N., Shafik, R.E., and Rasmy, G.E., 2011. Studies on the chemical constituents of fresh leaf of Eruca sativa extract and its biological activity as anticancer agent in vitro. Journal of Medicinal Plants Research (JMPR) 5(7): 1184–1191.
Morroni, F., Sita, G., Djemil, A., D’Amico, M., Pruccoli, L., Cantelli-Forti, G., et al. 2018. Comparison of adaptive neuroprotective mechanisms of sulforaphane and its interconversion product erucin in in vitro and in vivo models of Parkinson’s disease. Journal of Agricultural and Food Chemistry 66(4): 856–865. 10.1021/acs.jafc.7b04641
Motalebnejad, M., Karimmojeni, H., Majidi, M.M., and Mastinu, A., 2023. Allelopathic effect of aqueous extracts of grass genotypes on Eruca Sativa L. Plants 12(19): 3358. 10.3390/plants12193358
Moustafa, N.A., and Aida, E.M. 2002. Effect of Eruca sativa seeds oil on the non pregnant and pregnant female rats. Scientific Journal of Al-Azhar Medical Faculty (Girls) 23(2): 521–543.
Nazif, N.M., Amira, A.E., Wafaa, A.M., and Hassan, R.A., 2010. Chemical composition and cytotoxic activity of Eruca sativa L. seeds cultivated in Egypt. Asian Journal of Chemistry 22(3): 2407.
Nikzad, S., Mirmohammady Maibody, S.A.M., Ehtemam, M.H., Golkar, P., and Mohammadi, S.A., 2023. Response of seed yield and biochemical traits of Eruca sativa Mill. to drought stress in a collection study. Scientific Reports 13(1): 11157. 10.1038/s41598-023-38028-6.
Pagnotta, E., Ugolini, L., Matteo, R., and Righetti, L., 2022. Bioactive compounds from Eruca sativa seeds. Encyclopedia 2(4): 1866–1879. 10.3390/encyclopedia2040129
Palaiogiannis, D., Chatzimitakos, T., Athanasiadis, V., Bozinou, E., Makris, D.P., Lalas, S.I. 2023. Successive solvent extraction of polyphenols and flavonoids from Cistus creticus L. leaves. Oxygen 3: 274–286. 10.3390/oxygen3030018
Palliyaguru, D.L., Yuan, J.M., Kensler, T.W., and Fahey, J.W., 2018. Isothiocyanates: translating the power of plants to people. Molecular Nutrition & Food Research 62(18): 1700965. 10.1002/mnfr.201700965
Pandey, G., Khatoon, S., Pandey, M.M., and Rawat, A.K.S., 2018. Altitudinal variation of berberine, total phenolics and flavonoid content in Thalictrum foliolosum and their correlation with antimicrobial and antioxidant activities. Journal of Ayurveda and Integrative Medicine 9(3): 169–176. 10.1016/j.jaim.2017.02.010
Pane, C., Caputo, M., Francese, G., Manganiello, G., Lo Scalzo, R., Mennella, G., and Zaccardelli, M., 2020. Managing rhizoctonia damping-off of rocket (Eruca sativa) seedlings by drench application of bioactive potato leaf phytochemical extracts. Biology 9(9): 270. 10.3390/biology9090270
Park, J., Choi, W., Kim, J., Kim, H.W., Lee, J.Y., Lee, J., and Kim, B., 2024. Quantitative analysis and molecular docking simulation of flavonols from Eruca sativa Mill., and their effect on skin barrier function. Current Issues in Molecular Biology 46(1): 398–408. 10.3390/cimb46010025
Pasini, F., Verardo, V., Caboni, M.F., and D’Antuono, L.F., 2012. Determination of glucosinolates and phenolic compounds in rocket salad by HPLC-DAD–MS: evaluation of Eruca sativa Mill., and Diplotaxis tenuifolia L. genetic resources. Food Chemistry 133(3): 1025–1033. 10.1016/j.foodchem.2012.01.021
Peng, X., Wen, X., Pan, X., Wang, R., Chen, B., and Wu, C. 2010. Design and in vitro evaluation of capsaicin transdermal controlled release cubic phase gels. AAPS PharmSciTech 11: 1405–1410. 10.1208/s12249-010-9481-1
Piragine, E., Flori, L., Di Cesare Mannelli, L., Ghelardini, C., Pagnotta, E., Matteo, R., et al. 2021. Eruca sativa mill. seed extract promotes anti-obesity and hypoglycemic effects in mice fed with a high-fat diet. Phytotherapy Research 35(4): 1983–1990. 10.1002/ptr.6941
Qaddoumi, S., and El-Banna, N., 2019. Antimicrobial activity of arugula (Eruca sativa) leaves on some pathogenic bacteria. International Journal of Biology 11(3): 10–15. 10.5539/ijb.v11n3p10
Raffo, A., Masci, M., Moneta, E., Nicoli, S., Del Pulgar, J. S., and Paoletti, F., 2018. Characterization of volatiles and identification of odor-active compounds of rocket leaves. Food Chemistry 240: 1161–1170. 10.1016/j.foodchem.2017.08.009
Ramazzina, I., Lolli, V., Lacey, K., Tappi, S., Rocculi, P., and Rinaldi, M., 2022. Fresh-cut Eruca sativa treated with plasma activated water (PAW): evaluation of antioxidant capacity, polyphenolic profile and redox status in CaCo2 cells. Nutrients 14(24): 5337. 10.3390/nu14245337
Rizwana, H., Alwhibi, M.S., Khan, F., and Soliman, D.A., 2016. Chemical composition and antimicrobial activity of Eruca sativa seeds against pathogenic bacteria and fungi. J. Anim. Plant Sci. 26: 1859–1871. Corpus ID: 201921122.
Sadiq, A., Hayat, M.Q., and Mall, S.M., 2014. Qualitative and quantitative determination of secondary metabolites and antioxidant potential of Eruca sativa. Natural Products Chemistry & Research 2(4): 137. 10.4172/2329-6836.1000137
Saleh, M.M., Qader, S.W., and Thaker, A.A., 2016. Gastroprotective activity of Eruca sativa leaf extract on ethanol-induced gastric mucosal injury in Rattus norvegicus Jordan. Journal of Biological Sciences 9: 47–52. 10.12816/0027027
Salma, U., Khan, T., and Shah, A. J. 2018. Antihypertensive effect of the methanolic extract from Eruca sativa Mill, (Brassicaceae) in rats: Muscarinic receptor-linked vasorelaxant and cardiotonic effects. Journal of Ethnopharmacology 224: 409–420. 10.1016/j.jep.2018.06.013
Sanad, R.A.E.B., and Mabrouk, M.I. 2016. Development and assessment of stable formulations containing two herbal antimicrobials: Allium sativum L., and Eruca sativa miller seed oils. Drug Development and Industrial Pharmacy 42(6): 958–968. 10.3109/03639045.2015.1096280
Sharma, A., Sharma, R., Arora, R., Arora, S., Singh, B., and Sharma, U., 2017. Quantitative and qualitative analysis of Eruca sativa and Brassica juncea seeds by UPLC-DAD and UPLC-ESI-QTOF. Natural Product Communications 12(9): 1934578X1701200927. 10.1177/1934578X1701200927
Singh, M.P., Chawla, V., Kaushik, D., Chawla, P.A., and Sisodia, S.S. 2022. Pharmacodynamic stance of phytoconstituents as a gastric ulcer protective mechanism: an overview. Current Molecular Medicine 22(5): 431–441. 10.2174/1566524021666210806161835
Singh, S., Singh, G., Attri, S., Kaur, P., Rashid, F., Bedi, N., et al. 2023. Development and optimization of nanoparticles loaded with erucin, a dietary isothiocyanate isolated from Eruca sativa: antioxidant and antiproliferative activities in ehrlichascites carcinoma cell line. Frontiers in Pharmacology 13: 1080977. 10.3389/fphar.2022.1080977
Singh, D., Thakur, S., Singh, D., Buttar, H.S., Singh, B., and Arora, S., 2021. Modulatory effect of 4-(methylthio) butyl isothiocyanate isolated from Eruca sativa Thell. on DMBA-induced overexpression of hypoxia and glycolytic pathway in Sprague-Dawley female rats. Frontiers in Pharmacology 12: 728296. 10.3389/fphar.2021.728296
Sut, S., Boschiero, I., Solana, M., Malagoli, M., Bertucco, A., and Dall’Acqua, S., 2018. Supercritical CO2 extraction of Eruca sativa using cosolvents: phytochemical composition by LC-MS analysis. Molecules 23(12): 3240. 10.3390/molecules23123240
Testai, L., Pagnotta, E., Piragine, E., Flori, L., Citi, V., Martelli, A., et al. 2022. Cardiovascular benefits of Eruca sativa mill. Defatted seed meal extract: potential role of hydrogen sulfide. Phytotherapy Research 36(6): 2616–2627. 10.1002/ptr.7479
Tian, M., Yu, P., Li, Z., Liu, C., Liang, H., and Yuan, Q., 2024. Effects and mechanism of metal ions on the stability of glucosinolates in aqueous solution. Food Chemistry 448: 139098. 10.1016/j.foodchem.2024.139098
Wells, R. 2024. Origins and diversity of brassica and its relatives. In: Vegetable brassicas and related crucifers. CABI, GB, pp. 1–41. 10.1079/9781789249170.0001
Xu, L., Nagata, N., and Ota, T., 2018. Glucoraphanin: a broccoli sprout extract that ameliorates obesity-induced inflammation and insulin resistance. Adipocyte 7(3): 218–225. 10.1080/21623945.2018.1474669
Zhang, Y., Sun, L., Lai, X., Peng, X., Wen, S., Zhang, Z., et al. 2021. Gastroprotective effects of extract of Jasminum grandiflorum L. flower in HCl/EtOH-induced gastric mucosal ulceration mice. Biomedicine & Pharmacotherapy 144: 112268. 10.1016/j.biopha.2021.112268
Zhu, B., Qian, F., Hou, Y., Yang, W., Cai, M., and Wu, X., 2021. Complete chloroplast genome features and phylogenetic analysis of Eruca sativa (Brassicaceae). PLoS One 16(3): e0248556. 10.1371/journal.pone.0248556
Zidorn, C., and Stuppner, H., 2001. Evaluation of chemosystematic characters in the genus Leontodon (Asteraceae). Taxon 50(1): 115–133. 10.2307/1224515