
Download
ORIGINAL ARTICLE
Impact of thermal processing methods on the composition and content of 4′-O-methylpyridoxine analogues in Ginkgo biloba seeds
Gong Hao1, 2, Wu Cai-e1*, Kou Xiao-Hong3, Fan Gong-Jian1, 4, Li Ting-Ting1, 5, Wang Jia-Hong1, 4, Wang Tao1, 5
1College of Light Industry and Food Engineering, Nanjing Forestry University, Nanjing, China;
2College of Food Engineering, Xuzhou University of Technology, Xuzhou, China;
3School of Chemical Engineering and Technology, Tianjin University, Tianjin, China;
4Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing, China;
5Department of Chemistry Engineering, Xuzhou College of Industrial Technology, Xuzhou, China
Abstract
This study investigated the effect of thermal processing methods, namely, boiling, microwaving, baking, and frying, on the 4′-O-methylpyridoxine (MPN) analogues of Gingko biloba seeds. All thermal processing methods decreased MPN, pyridoxine, and pyridoxal-5′-phosphate; total MPN; and total vitamin B6 contents but increased MPN -5′-glucoside and pyridoxamine contents. Baking and frying reduced total MPN content by 36.38–54.56% and 46.54–54.67%, respectively. Frying was identified as the optimal thermal processing method that maintains the total vitamin B6 compound content of G. biloba seeds at high levels (72.92–84.62%). Principal component analysis revealed the different effects of thermal processing methods on MPN analogues in G. biloba seeds. The results of this study demonstrate that compared with other thermal processing methods, frying can better reduce the toxic compound content (total MPN) of G. biloba seeds and promote vitamin B6 retention.
Key words: Ginkgo biloba seeds, thermal processing methods, thermal processing parameter, 4′-O-methylpyridoxine, vitamin B6
*Corresponding Author: Cai-e Wu, College of Light Industry and Food Engineering, Nanjing Forestry University, Nanjing, China. Tel/Fax: +86-25-85427844. Email: [email protected]
Received: 20-04-2020| Accepted: 08-07-2020| Published: 22-08-2020
© 2020 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Ginkgo biloba L. is a living fossil that has existed on earth for 200,000,000 years. Ginkgo biloba (G. biloba) seeds have been used as traditional food in China, Japan, and Korea (Huang et al. 2010). The overconsumption of G. biloba seeds can result in poisoning (Leistner and Drewke 2010). The symptoms of G. biloba poisoning include abdominal pain, diarrhea, clonic convulsions, and unconsciousness (Wada et al. 1988). The poisonous effects of G. biloba are ascribed to 4′-O-methylpyridoxine (MPN) and its glycoside forms (Leistner and Drewke 2010). MPN, a neurotoxin with actions against vitamin B6 (VB6) compounds, can reduce the pyridoxal 5′-phosphorylated (PLP) contents of the human body because its affinity for human pyridoxal kinase is higher than that for pyridoxal (PL) (Buss et al. 2001; Kästner et al. 2007). VB6 is a group of six interconvertible water-soluble vitamins that are based on pyridine (Table 1). These vitamins include pyridoxine (PN), pyridoxamine (PM), PL, and their pyridoxal 5′-phosphorylated forms, namely, PNP, PMP, and PLP (Hellmann and Mooney 2010). VB6 comprises essential organic micronutrients in the human diet. They participate in the metabolism of amino acids, carbohydrates, lipids, and neurotransmitters (Eliot and Kirsch 2004; Fudge et al. 2017). They also demonstrate important biological activities, such as immunomodulatory, antitumor, and anti-oxidative activities (Eliot and Kirsch 2004; Galluzzi et al. 2012; Mesripour et al. 2017).
Table 1. Chemical structure of the investigated 4’-O-methylpyridoxine analogues.
| Number | R1 | R2 | Analytes | |
|---|---|---|---|---|
| 1 | CH2OH | H | Pyridoxine (PN) | |
| 2 | CHO | H | Pyridoxal (PL) | |
| 3 | CH2NH2 | H | Pyridoxine (PM) | |
| 4 | CHO | PO3H2 | Pyridoxal-5´-phosphate (PLP) | |
| 5 | CH2NH2 | PO3H2 | Pyridoxine-5´-phosphate (PMP) | |
| 6 | CH2OCH3 | H | 4’-O-methylpyridoxine (MPN) | |
| 7 | CH2OCH3 | C6H11O6 | 4’-O-methylpyridoxine-5´-glucoside (MPNG) |
Physical and chemical factors, including heat, light exposure, and pH, can influence the VB6 content of foodstuffs (Park et al. 2016). G. biloba seeds are consumed after processing through boiling, microwaving, baking, or frying (Goh and Barlow 2002). MPN is converted to MPNG (4’-O-methylpyridoxine-5´-glucoside) in boiled and microwaved G. biloba seeds (Kobayashi et al. 2011). Heating results in the loss of PL and PN from G. biloba seeds (Yoshimura et al. 2006). The other forms of MPN analogues, such as PLP, PMP, and PM, were not investigated during the thermal processing. Baking and frying are some of the most commonly used methods for the preparation of G. biloba seeds. Information on the effects of baking and frying on MPN analogues in G. biloba seeds, however, remain limited. Thermal processing parameters can also affect the VB6 contents of various foodstuffs (Leskova et al. 2006). Thermal processing duration and temperature, oxygen exposure, and water are the major factors involved in thermal processing methods and may exert potentially harmful or beneficial effects on food. Thus far, however, no studies have focused on the effect of thermal processing parameters on MPN analogues in G. biloba seeds. Plants are an important source of vitamins that cannot be synthesized de novo by humans (Salvo et al. 2011). G. biloba seeds are potential sources of VB6 (Gong et al. 2018) and MPN, a VB6 derivative. Thermal processing methods may positively or negatively influence the composition and contents of MPN analogues in G. biloba seeds. Current studies have failed to fully evaluate the changes in the contents of toxic substances, such as MPN and MPNG, and nutrients in G. biloba seeds during thermal processing.
This study focused on the effects of thermal processing on MPN analogues, including MPN, MPNG, PMP, PLP, PM, PL, and PN, in G. biloba seeds. The specific aims of this study were (i) to evaluate the effects of different types of thermal processing methods, namely, boiling, microwaving, baking, and frying, on the composition and contents of MPN analogues and (ii) to study the effects of boiling time, microwave power, and baking and frying temperatures on MPN analogues in G. biloba seeds. The present study will describe the changes in MPN analogues during thermal processing and recommend a thermal processing method that will decrease toxic MPN contents while promoting VB6 retention in G. biloba seeds.
Materials and Methods
Chemicals and standards
MPN and MPNG standards with 98% purity were synthesized by Kangbei Biochemical Co. (Ningbo, China) through high-performance liquid chromatography (HPLC). Standards, including PLP, PMP, pyridoxal hydrochloride, pyridoxamine dihydrochloride, and pyridoxine hydrochloride (≥98%), were purchased from Yuanye Biological Technology Co. (Shanghai, China). HPLC-grade acetonitrile and methanol were acquired from Tedia Company Inc. (Ohio, USA). HPLC-grade phosphoric acid (85%) was obtained from Kemiou Chemical Reagent Co. (Tianjin, China). HPLC-grade sodium pentanesulfonate (≥99.5%) was procured from Yuwang Industrial Co. (Shandong, China). Other reagents were of analytical grade. Distilled water was prepared using a Milli-Q system (Millipore A10; Billerica, MA, USA).
Plant materials and thermal processing methods
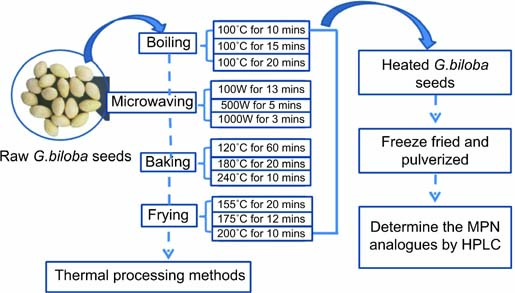
The G. biloba seeds used in this study were harvested in 2017 in Pizhou, Jiangsu Province, China. The episperm, mesosperm, and endopleura were removed from each collected seed. The processed seeds (approximately 5000 g) were then divided into five groups. One group was analyzed raw (control). The other four groups were subjected to boiling for different durations (10, 15, and 20 min), microwave treatment at different powers (100, 500, and 1000 W), baking treatment at different temperatures (120°, 180°, and 240°), and frying treatment at different temperatures (150°, 175°, and 200°). Raw and heated seeds were freeze-dried, pulverized, and stored at 0°C for further analysis. Each heated sample was dehydrated in an oven at 105°C for the determination of water content. Oil content after frying was determined through petroleum ether extraction. Thermal processing details, including heating conditions and times, are summarized in Figure 1. The minimum and maximum duration of each thermal processing method met the requirements set by professional judges.
Figure 1. Diagram of the thermal processing of Ginkgo biloba seeds. MPN, methylpyridoxine; HPLC, high-performance liquid chromatography.
MPN analogue determination
HPLC methods were applied to determine the composition and contents of MPN analogues in raw and heated G. biloba seeds. The HPLC conditions were set in accordance with a previously reported method with slight modifications (Teruki et al. 2006). MPN analogues were chromatographically analyzed with a Waters HPLC 2695 system (Waters, Milford, USA) equipped with a fluorescence measurement apparatus (Emission wavelength: 395 nm; excitation wavelength: 295 nm). Separation was performed with a Waters XBridge RP18 column (250 mm × 4.6 mm, 5 μm; Waters Corp., USA). Mobile phase A comprised 5 mmol/L potassium phosphate containing 5 mmol/L sodium pentanesulfonate. The pH of the mobile phase was adjusted to 2.5 using phosphoric acid. Acetonitrile was used as mobile phase B. Column temperature was maintained at 30°C, flow rate was 1.0 mL/min, and injection volume was 10 μL. The HPLC gradient program was as follows: from 4% mobile phase B to 8.5% mobile phase B at 15 min, to 15% mobile phase B at 30 min, and to 4% mobile phase B in the final 20 min. The linearity, correlation coefficient, linear range, limit of detection, limit of quantification, precious test, and accuracy of the HPLC methods used in this study had been validated in previous works (Gong et al. 2018).
For the analysis of the MPN analogue, contents of raw and heated G. biloba seeds, 50 mg of freeze-dried powders, were mixed with 1.5 mL of distilled water (adjusted to pH 2.5 by using phosphoric acid). The mixtures were incubated on a shaking table (220 r/min) (Jinghong, THZ320, Shanghai, China) at 25°C for 40 min and then centrifuged at 9167 × g for 30 min at 4°C (Sigma, model 2-16K, Germany). The supernatant was filtered with a 0.45 μm syringe filter (Jinlong, Tianjin, China) and injected into the HPLC system. All of the samples were prepared in triplicate.
Statistical analysis
Results were expressed as the mean and standard deviation of the results for three replicates. The effect of thermal processing methods on MPN analogues in G. biloba seeds was evaluated through one-way ANOVA. Statistically significant differences between mean values at the 5% (P < 0.05) level were determined with Tukey’s test. Principal component analysis (PCA) was used to determine the different changes in MPN analogues during thermal processing. Analyses were performed using SPSS version 20.0 (USA).
Results
Composition and contents of MPN analogues in raw and heated G. biloba seeds
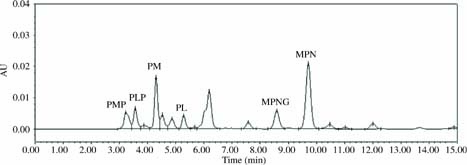
The representative HPLC chromatography of MPN analogs obtained from raw G. biloba seeds are shown in Figure 2. The traditional thermal processing methods of boiling, microwaving, baking, and frying significantly affected the composition and contents of MPN analogues in G. biloba seeds (P < 0.05). MPN analogues showed different changes when processed through the four different thermal processing methods under different thermal processing parameters (Table 2). The MPN contents of seeds subjected to the four thermal processing treatments were lower than those of raw seeds. Seeds that had been baked at 120°C and those that had been fried at 150°C had the lowest MPN contents (P < 0.05). The MPN contents of seeds that had been microwaved changed negligibly. The reduction in MPN contents decreased as baking temperature increased. The MPN contents of seeds that had been boiled for different durations (10 and 20 min) and fried at different temperatures (150 and 200°C) varied marginally. The MPNG contents of heated seeds were higher than those of raw samples, irrespective of the thermal processing method. In addition, samples that have been boiled for 10 min had the highest MPNG contents among all cooked samples (P < 0.05). MPNG contents decreased considerably with prolonged boiling time and frying temperatures but did not change with the change in microwaving and baking processing parameters.
Figure 2. The representative HPLC chromatography of MPN analogs obtained from raw G. biloba seeds (CK). HPLC, high-performance liquid chromatography; MPN, 4’-O-methylpyridoxine.
Table 2. The composition and contents of 4’-O-methylpyridoxine analogues in raw and heated Ginkgo biloba seeds (μg/g).
| MPN | MPNG | PMP | PLP | PM | PL | PN | |
|---|---|---|---|---|---|---|---|
| Raw | 175.98 ± 0.87a | 108.71 ± 4.44d | 24.68 ± 0.23a | 64.05 ± 3.12a | 52.11 ± 12.11d | 49.35 ± 0.42a | 142.72 ± 0.20a |
| Boiling—10 min | 29.29 ± 2.75b | 259.85 ± 10.23a | 7.34 ± 0.13b | 52.90 ± 1.67b | 71.98 ± 0.15a | 17.98 ± 0.77b | 10.25 ± 1.18d |
| Boiling—15 min | 24.71 ± 2.03c | 236.20 ± 1.70b | 7.20 ±0.03b | 43.46 ± 1.11c | 64.90 ± 0.45c | 15.84 ± 0.27c | 17.68 ± 1.28c |
| Boiled—20 min | 32.16 ± 0.74b | 220.19 ± 2.11c | 7.14 ± 0.07b | 40.26 ± 0.44c | 68.82 ± 0.73b | 14.77 ± 0.42d | 21.59 ± 0.25b |
| Raw | 175.98 ± 0.87a | 108.71 ± 4.44c | 24.68 ± 0.23a | 64.05 ± 3.12a | 52.11 ± 12.11c | 49.35 ± 0.42a | 142.72 ± 0.20a |
| Microwaving—100W | 30.19 ± 0.42b | 250.80 ± 0.37a | 7.77 ± 0.01b | 49.20 ± 0.34c | 83.02 ± 0.74b | 16.66 ± 0.03d | 25.06 ± 6.63c |
| Microwaving—500 W | 30.60 ± 0.11b | 239.65 ± 0.10b | 7.96 ± 0.06b | 53.47 ± 2.36b | 84.58 ± 4.86b | 20.79 ± 4.61c | 35.42 ± 10.70c |
| Microwaving—1000 W | 30.52 ± 0.43b | 246.36 ± 3.18a | 7.93 ± 0.04b | 49.45 ± 1.79bc | 102.36 ± 2.08a | 26.70 ± 0.63b | 65.37 ± 1.55b |
| Raw | 175.98 ± 0.87a | 108.71 ± 4.44d | 24.68 ± 0.23a | 64.05 ± 3.12a | 52.11 ± 12.11d | 49.35 ± 0.42a | 142.72 ± 0.20a |
| Baking—120°C | 17.48 ± 0.13c | 203.24 ± 2.71b | 5.89 ± 0.11c | 36.41 ± 1.98c | 70.18 ± 0.67c | 35.05 ± 4.13b | 15.93 ± 0.45d |
| Baking—180°C | 15.70 ± 0.20d | 170.52 ± 0.84c | 4.84 ± 0.04d | 32.07 ± 0.56d | 97.98 ± 1.85a | 32.00 ± 0.96c | 34.20 ± 0.01c |
| Baking—240°C | 26.53 ± 0.38b | 230.16 ± 0.11a | 6.62 ± 0.12b | 43.57 ± 1.40b | 76.86 ± 1.48b | 31.26 ± 0.38d | 38.49 ± 0.58b |
| Raw | 175.98 ± 0.87a | 108.71 ± 4.44d | 24.68 ± 0.23a | 64.05 ± 3.12a | 52.11 ± 2.11d | 49.35 ± 0.42d | 142.72 ± 0.20a |
| Frying—150°C | 17.91 ± 1.04c | 201.55 ± 0.19a | 8.28 ± 0.33b | 44.12 ± 7.02d | 104.81 ± 2.64a | 92.64 ± 1.54a | 19.05 ± 3.16c |
| Frying—175°C | 24.03 ± 3.54b | 186.30 ± 7.39b | 7.56 ± 0.45c | 47.68 ± 3.68c | 82.23 ± 3.58b | 83.2 ± 0.65b | 32.95 ± 0.69b |
| Frying—200°C | 19.88 ± 0.20c | 162.14 ± 0.26c | 6.39 ± 0.04d | 59.67 ± 0.60b | 71.22 ± 0.30c | 65.67 ± 0.70c | 32.50 ± 0.01b |
Values are the mean ± standard deviation (SD) from three replicates.
Different letters (a–d) in each column means significant differences under the different thermal processing methods conditions (P < 0.05).
MPN, 4’-O-methylpyridoxine; MPNG, 4’-O-methylpyridoxine-5´-glucoside; PMP, phosphate; PLP, phosphorylated; PL, pyridoxal; PN, pyridoxine; PM, pyridoxamine.
Most thermal processing methods decreased the PMP, PLP, PN, and PL contents of seeds. The reduction in the content of PN, the major VB6 compound, was more intense than that in the contents of other VB6 forms. The reduction in PN contents decreased as boiling time and microwave power increased. Seeds microwaved at 100 W and fried at 150°C had the lowest PN contents. The reduction in PL content intensified as boiling time and baking temperature increased but weakened as microwaving power increased. Only frying increased the PL content of G. biloba seeds. Increasing the frying temperature from 150 to 200° led to an increase in PL content. Boiling time and microwave power did not exert marked effects on PMP content. Meanwhile, the PLP contents of cooked G. biloba seeds changed slightly. PM contents increased after thermal processing. The increase in the PM contents of fried seeds was related to increases in frying temperature. Samples fried at 150°C had the highest PM content. These results indicate that thermal processing methods and parameters have important effects on the composition and content of MPN compounds in G. biloba seeds.
Total MPN and VB6 contents of raw and cooked G. biloba seeds
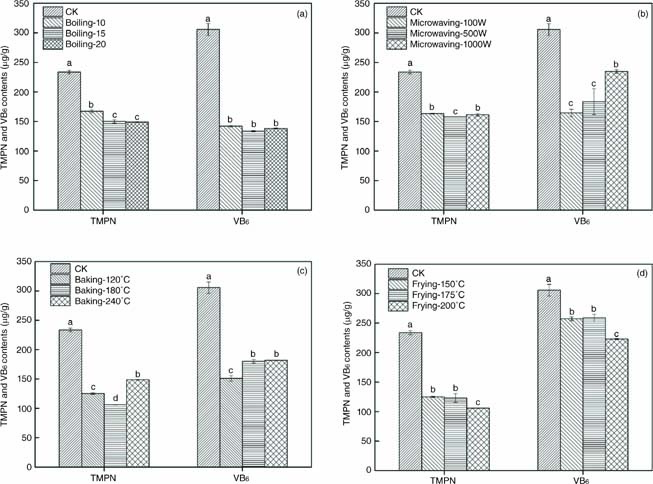
The total MPN (Total 4’-O-methylpyridoxine) and total VB6 contents of the seeds were calculated on the basis of the molecular mass of MPN (183.09 g/mol) and PN (168.19 g/mol) to further analyze the changes in MPN analogues during thermal processing. The TMPN and VB6 contents of raw and cooked G. biloba seeds are presented in Figure 3A–D. The results show that boiling, microwaving, baking, and frying significantly decreased the TMPN and VB6 contents of cooked seeds. Baking and frying resulted in greater losses in TMPN contents than other methods. Baking and frying decreased TMPN contents by 36.38–54.56% and 46.54–54.67%, respectively. TMPN content drastically changed during baking with different process parameters, and seeds baked at 180° had the lowest TMPN content. The TMPN contents of seeds fried at 150° and 175° showed minimal changes. Meanwhile, G. biloba seeds fried at 200° and baked at 180° had the lowest TMPN contents. On the other hand, boiling and baking resulted in severe reductions in VB6 contents (53.53–56.28% and 40.54–50.63%, respectively), whereas frying resulted in slight losses in VB6 contents (15.88–27.08%). The change in TMPN contents of seeds boiled for different durations was negligible. Baking temperature had a marked effect on VB6 retention by the cooked seeds. The reduction in VB6 contents did not change when microwaving power and baking temperature increased from 100 W to 500 W and from 180° to 240°, respectively. These results show that different thermal processing methods and parameters have different effects on the TMPN and VB6 contents of G. biloba seeds.
Figure 3. Total 4’-O-methylpyridoxine and vitamin B6 in raw and heated Ginkgo biloba seeds (A: boiling; B: microwaving; C: baking; D: frying). Each value represents the mean of three replicates, and error bars indicate standard deviation (± SD). Different letters (a–d) mean the significant difference between different thermal processing parameters (P < 0.05). CK, raw Ginkgo biloba seeds; TMPN, total 4’-O-methylpyridoxine; VB6: total vitamin B6.
Principal component analysis
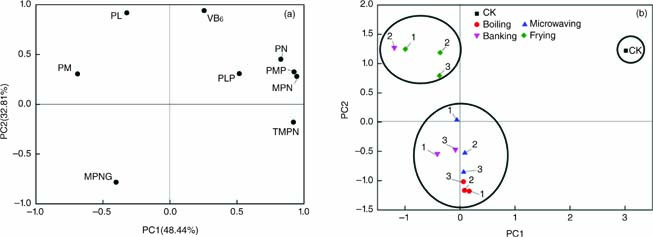
The normalized data for the contents of MPN analogues, including MPN, MPNG, TMPN, PMP, PLP, PM, PL, PN, and VB6, were subjected to PCA to clarify the similarities and differences between raw seeds and seeds processed through four different thermal processing methods. Two principal components (PCs) with eigenvalues >1 were obtained in accordance with Kaiser’s rules. These PCs accounted for 81.25% of the total observed variance. The PCA results revealed that the MPN analogue content of heated seeds were distinct from those of raw seeds. The loading plots between PC1 and PC2 are presented in Figure 3A. PC1 accounted for 48.44% of the total variation and was positively correlated with PMP (0.93), MPN (0.95), TMPN (0.92), PN (0.83), and PLP (0.52). PM and MPNG were negative contributors to PC2 and had values of −0.69 and −0.40, respectively. PC2 explained 32.81% of the total variation and was positively correlated with VB6 (0.94) and PL (0.92), and negatively correlated with MPNG (−0.0.78).
The score plots of PC1 and PC2 obtained for raw samples and the seeds processed through four different thermal processing methods under different parameters are shown in Figure 4B. Raw seeds (CK) formed a separate cluster in the PC2 score plot presented in Figure 4B because the MPN analogue contents of raw seeds were different from those of heated seeds. Fried samples and samples that were microwaved at 100 W localized at high areas on the plot because they retained high VB6 contents. In addition, G. biloba seeds processed through frying have higher PL contents and lower MPNG contents than G. biloba seeds processed through other methods. The positions of seeds processed through three other thermal processing methods were determined by their high VB6 loss and MPNG content. Boiled seeds localized at the bottom of Figure 3B because their PL contents drastically decreased and their MPNG contents increased. Therefore, PCA results can be used to identify the effects of different thermal processing methods on the basis of the MPN analogue contents of raw and heated G. biloba seeds.
Figure 4. Loading plots and score plots obtained from the principal component analysis of selected variables (A and B). PC1, principal component 1; PC2, principal component 1; CK, raw Ginkgo biloba seeds; MPNG, 4’-O-methylpyridoxine-5´-glucoside; PM, pyridoxamine; PL, pyridoxal; VB6, vitamin B6; PLP, phosphorylated; PN, pyridoxine; PMP, phosphate;
Discussion
G. biloba seeds are consumed after processing through different thermal processing methods, including boiling, microwaving, baking, and frying. Boiled and fried G. biloba seeds are also used in the food processing industry. This work investigated the changes exhibited by MPN analogues in heated G. biloba seeds. The influence of thermal processing parameters on MPN analogues was first investigated. Thermal processing induced the loss and gain of MPN analogues in G. biloba seeds. These effects are dependent on the thermal processing technique and its parameters, including time, power, and temperature.
All thermal processing methods decreased the contents of the toxic compounds, MPN and TMPN, and increased those of MPNG in G. biloba seeds. The result is consistent with the results of previous studies showing that MPNG is the predominant compound in processed G. biloba seeds (Lawrence and Scott 2005; Yoshimura et al. 2006). Small amounts of MPN are also present in boiled and microwaved G. biloba seeds (Kobayashi et al. 2011). The reduction in TMPN contents indicates that the structure of MPN was destroyed during thermal processing. Baking and frying promoted the removal of toxic substances, such as TMPN, from G. biloba seeds. Frying at 180° for 20 min and baking at 175° for 12 min reduced TMPN contents by 54.56 and 54.67%, respectively.
The changes in VB6 contents during boiling, microwaving, frying, or baking have previously have been investigated (Sierra and Vidalvalverde 2001). In support of the results of the present study, the results of past studies have shown that thermal processing resulted in the loss of VB6 (Leskova et al. 2006). For example, Beyza and Akif (2009) reported that baking, microwaving, and frying decreased the VB6 contents of African catfish. The VB6 contents of bread decreased by 15% during baking (Perera et al. 1980). Boiling resulted in the loss of 16 and 61% of the VB6 contents of Brussels sprouts and broccoli, respectively. Microwaving resulted in low losses of the PN contents of Brussels sprouts and broccoli (Leskova et al. 2006). In this work, boiling resulted in drastic reductions in PN content, whereas microwaving resulted in low losses of PN content. Frying and roasting reduced the PN content of beef and lamb (Purchas et al. 2014). In the present study, the PM contents of G. biloba seeds heated through the four tested methods increased. Similar results were found for milk treated through different heating methods (Schmidt and Mayer 2018). The contents of phosphorylated VB6 (PMP and PLP) decreased during thermal processing. The reduction in PMP content was higher than the PLP content. PMP and PLP are present at low levels in various foodstuffs. Limited information is available on the changes in the PMP and PLP contents of various foodstuffs after thermal processing. PMP and PL may undergo dephosphorylation, transamination, and depletion during heating (Gregory et al. 1986).
All four thermal processing methods used in this work are heat-treatment processes. Heating treatment may result in the transformation and degradation of substances in foodstuffs (Santiago et al. 2018). The increase in the MPNG contents of cooked G. biloba seeds may be attributed to the conversion of MPN to MPNG (Kobayashi et al. 2011). MPN in G. biloba seed extracts will be thermally degraded into PN at high temperatures above 120°C (Lim and Kim 2018). The conversion of MPN to MPNG and the thermal degradation of MPN to PN may account for the reduction in the MPN content of cooked G. biloba seeds. Furthermore, all thermal processing methods can increase the PM contents of G. biloba seeds. PN may be partially converted into PM and PL in heated broccoli and milk (Leskova et al. 2006; Sierra and Vidalvalverde 2001). Heating temperature and duration play important roles in the changes of MPN analogues in G. biloba seeds. Lim and Kim (2018) reported that the reduction in the MPN contents of G. biloba extract intensified as heating temperature (120–150°) and duration (0–60 min) increased. Previous results also indicated that prolonging heating time (1.5–6 h) can intensify the loss of VB6 in the form of PN from cauliflower puree. Meanwhile, losses in VB6 increase when heating temperatures are increased from 105.9 to 137.7°C (Navankasattusas and Lund 2010). In this work, thermal processing methods, such as baking and frying, that require high temperatures resulted in drastic reductions in MPN and TMPN contents, and slight increases in MPNG content. These results indicate that VB6 may be more stable than MPN and MPNG or that MPN analogues are converted (MPN × PN × other VB6 forms) during heating.
Thermal processing methods that do not require the addition of water could prevent the leaching of water-soluble compounds (Santiago et al. 2017). Boiling resulted in severe reduction in MPN analogue (PN and VB6) contents. As previously reported, additional losses from boiled G. biloba seeds may be mainly attributed to the leaching of MPN analogues into water. Conversely, the use of oil in frying can prevent the dissolution of water-soluble MPN analogues. Hence, frying resulted in the lowest reduction in VB6 content. The greater lipophilicity of MPN than that of other MPN forms (Kästner et al. 2007) may account for the higher losses of TMPN content than that in VB6 during frying. Microwaving can destroy cell walls and subcellular compartments; these changes, in turn, can promote the dissolution of intracellular material (Santiago et al. 2018). Microwave treatment for short durations can promote the preservation of vitamins in vegetables (Lee et al. 2017). Similarly, MPN, PN, and PM were retained in G. biloba seeds processed through 3 min of microwave treatment at 1000 W. Microwave equipment produces high-frequency vibration (2450 MHz) that introduces friction and promotes collision among polar molecules. MPN analogues were disintegrated by microwave treatment. G. biloba seeds are a starch-rich material and may undergo starch gelation after heating. Starch gelation subsequently hinders the extraction of MPN analogues during HPLC detection (Scott et al. 2000) and may account for the reduction in the MPN analogue content of G. biloba seeds.
Conclusion
Thermal processing methods and conditions can drastically influence the composition and contents of MPN analogues in G. biloba seeds. The stabilities of different MPN analogues differed during thermal processing. Thermal processing decreased MPN, PMP, PL, PN, TMPN, and VB6 contents, and increased MPNG and PM contents in G. biloba seeds. High-temperature treatments, including baking and frying, reduced the toxic substance (MPN and MPNG) content of G. biloba seeds. Boiling resulted in the dissolution of PL, PM, and PN in water, and in the severe loss of VB6. Moreover, the physical effects of microwaving caused the degradation of VB6 in G. biloba seeds. Short treatments, such as microwaving at 1000 W, resulted in the retention of some VB6 forms, such as PM and PN. PCA results revealed the similarities and differences among the four different thermal processing methods. Results suggest that many factors contribute to the complex changes experienced by MPN analogues during thermal processing. These factors include high heat-induced conversion and degradation, water- and oil-induced changes, microwave irradiation, and starch gelation.
Acknowledgements
This work was supported by the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX18_0987), Natural Science Foundation of Jiangsu Province (BE2015315), and Natural Youth Foundation of Jiangsu Province (BK20150883). The authors are grateful for the support of the priority academic program development (PAPD) of Jiangsu higher education institutions and co-innovation entre for sustainable forestry in southern China.
REFERENCES
Beyza, E. and Akif, O.Z., 2009. The effect of cooking methods on mineral and vitamin contents of African catfish. Food Chemistry 115: 419–422. 10.1016/j.foodchem.2008.12.018
Buss, K., Drewke, C., Lohmann, S., Piwonska, A. and Leistner, E., 2001. Properties and interaction of heterologously expressed glutamate decarboxylase isoenzymes GAD(65kDa) and GAD(67kDa) from human brain with ginkgotoxin and its 5’-phosphate. Journal of Medicine Chemistry 44: 3166–3174. 10.1021/jm010868f
Eliot, A.C. and Kirsch, J.F., 2004. Pyridoxal phosphate enzymes: Mechanistic, structural, and evolutionary considerations. Annual Review of Biochemistry 73: 383–415. 10.1146/annurev.biochem.73.011303.074021
Fudge, J., Mangel, N., Gruissem, W., Vanderschuren, H. and Fitzpatrick, T.B., 2017. Rationalising vitamin B6 biofortification in crop plants. Current Opinion in Biotechnology 44: 130–137. 10.1016/j.copbio.2016.12.004
Galluzzi, L., Vitale, I., Senovilla, L., Olaussen, K.A., Pinna, G., Eisenberg, T., Goubar, A., Martins, I., Michels, J. and Kratassiouk, G., 2012. Prognostic impact of vitamin B6 metabolism in lung cancer. Cell Reports 2: 257–269.
Goh, L.M. and Barlow, P.J., 2002. Antioxidant capacity in Ginkgo biloba. Food Research International 35: 815–820. 10.1016/S0963-9969(02)00084-4
Gong, H., Wu, C.E., Fan, G.J., Li, T.T., Wang, J.H. and Wang, T., 2018. Determination and comparison of 4’-O-methylpyridoxine analogs in Ginkgo biloba seeds in different growth stages. Journal of Agriculture and Food Chemistry 66: 7916–7922. 10.1021/acs.jafc.8b02522
Gregory, J.F., Ink, S.L. and Sartain, D.B., 1986. Degradation and binding to food proteins of vitamin B6 compounds during thermal processing. Journal of Food Science 51: 1345–1351. 10.1111/j.1365-2621.1986.tb13119.x
Hellmann, H. and Mooney, S., 2010. Vitamin B6: a molecule for human health? Molecules 15: 442–459. 10.3390/molecules15010442
Huang, W., Deng, Q.C., Xie, B.J., Shi, J., Huang, F.H., Tian, B.Q., Huang, Q.D. and Xue, S., 2010. Purification and characterization of an antioxidant protein from Ginkgo biloba seeds. Food Research International 43: 86–94. 10.1016/j.foodres.2009.08.015
Kästner, U., Hallmen, C., Wiese, M., Leistner, E. and Drewke, C., 2007. The human pyridoxal kinase, a plausible target for ginkgotoxin from Ginkgo biloba. FEBS Journal 274: 1036–1045. 10.1111/j.1742-4658.2007.05654.x
Kobayashi, D., Yoshimura, T., Johno, A., Sasaki, K. and Wada, K., 2011. Toxicity of 4’-O-methylpyridoxine-5’-glucoside in Ginkgo biloba seeds. Food Chemistry 126: 1198–1202. 10.1016/j.foodchem.2010.12.001
Lawrence, G.A. and Scott, P.M., 2005. Improved extraction of ginkgotoxin (4’-O-methylpyridoxine) from Ginkgo biloba products. Journal of AOAC International 88: 26–29. 10.1093/jaoac/88.1.26
Lee, S., Choi, Y., Jeong, H.S., Lee, J. and Sung, J., 2017. Effect of different cooking methods on the content of vitamins and true retention in selected vegetables. Food Science Biotechnology 27: 1–10. 10.1007/s10068-017-0281-1
Leistner, E. and Drewke, C., 2010. Ginkgo biloba and ginkgotoxin. Journal of Nature Products 73: 86–93. 10.1021/np9005019
Leskova, E., Kubikova, J., Kovacikova, E., Kosicka, M., Porubska, J. and Holcikova, K., 2006. Vitamin losses: Retention during heat treatment and continual changes expressed by mathematical models. Journal of Food Composition and Analysis 19: 252–276. 10.1016/j.jfca.2005.04.014
Lim, H.B. and Kim, D.H., 2018. Effect of heat treatment on 4’-O-methylpyridoxine (MPN) content in Ginkgo biloba seed extract solution. Journal of the Science of Food and Agriculture 98: 5153–5156. 10.1002/jsfa.9017
Mesripour, A., Hajhashemi, V. and Kuchak, A., 2017. Effect of concomitant administration of three different antidepressants with vitamin B6 on depression and obsessive compulsive disorder in mice models. Research in Pharmaceutical Sciences 12: 46–52. 10.4103/1735-5362.199046
Navankasattusas, S. and Lund, D.B., 2010. Thermal destruction of vitamin B6 vitamers in buffer aolution and cauliflower puree. Journal of food Science 47: 1512–1518. 10.1111/j.1365-2621.1982.tb04972.x
Park, J.E., Kim, K.E., Choi, Y.J., Park, Y.D. and Kwon, H.J., 2016. The stability of water-and fat-soluble vitamin in dentifrices according to pH level and storage type. Biomedical Chromatography 30: 191–199. 10.1002/bmc.3535
Perera, A.D., Leklem, J.E. and Miller, L.T., 1980. Stability of vitamin B6 during bread making and storage of bread and flour. Cereal Chemistry 56: 577–580.
Purchas, R.W., Wilkinson, B.H.P., Carruthers, F., Jackson, F. and Carruthers, F., 2014. A comparison of the nutrient content of uncooked and cooked lean from New Zealand beef and lamb. Journal of Food Composition and Analysis 35: 75–82. 10.1016/j.jfca.2014.04.008
Salvo, M.L.D., Contestabile, R. and Safo, M.K., 2011. Vitamin B6 salvage enzymes: mechanism, structure and regulation*. Biochimica et Biophysica Acta 1814: 1597–1608. 10.1016/j.bbapap.2010.12.006
Santiago, E.D., Caro, G.P., Rojas, J.M.M., Cid, C. and Paz De Peña, M., 2018. Digestibility of (poly)phenols and antioxidant activity in raw and cooked cactus cladodes (Opuntia ficus-indica). Journal of Agriculture and Food Chemistry 66: 5832–5844. 10.1021/acs.jafc.8b01167
Santiago, E.D., Domínguez-Fernández, M., Cid, C. and Peña, M.P.D., 2017. Impact of cooking process on nutritional composition and antioxidants of cactus cladodes (Opuntia ficus-indica). Food Chemistry 240: 1055–1062. 10.1016/j.foodchem.2017.08.039
Schmidt, A. and Mayer, H.K., 2018. Milk process authentication by vitamin B6 as a novel time temperature integrator. Food Control 91: 123–127. 10.1016/j.foodcont.2018.03.024
Scott, P.M., Lau, B.P., Lawrence, G.A. and Lewis, D.A., 2000. Analysis of Ginkgo biloba for the presence of ginkgotoxin and ginkgotoxin 5’-glucoside. Journal of AOAC International 83: 1313–1320. 10.1093/jaoac/83.6.1313
Sierra, I. and Vidalvalverde, C., 2001. Vitamin B1 and B6 retention in milk after continuous-flow microwave and conventional heating at high temperatures. Journal of Food Protection 64: 890–894. 10.4315/0362-028X-64.6.890
Wada, K., Ishigaki, S., Ueda, K., Take, Y., Sasaki, K., Sakata, M. and Haga, M., 1988. Studies on the constitution of edible and medicinal plants. I. Isolation and identification of 4-O-methylpyridoxine, toxic principle from the seed of Ginkgo biloba L. Chemical and Pharmaceutical Bulletin 36: 1779–1782. 10.1248/cpb.36.1779
Yoshimura, T., Udaka, N., Morita, J., Zhang, J.Y., Sasaki, K., Kobayashi, D., Wada, K. and Hori, Y., 2006. High performance liquid chromatographic determination of ginkgotoxin and ginkgotoxin-5’-glucoside in seeds. Journal of Liquid Chromatography & Related Technologies 29: 605–616. 10.1080/10826070500531466