
Download
Original Article
The inhibitory effects of polyphenolic compounds on the damage caused by safflower fly (Acanthiophilus helianthi) in Carthamus spp.
Karami Soraya1*, Basaki Tayebeh1, Amin Mousavi Khaneghah2
1Department of Agricultural Science, Payame Noor University, Tehran, Iran;
2Department of Food Science and Nutrition, Faculty of Food Engineering, University of Campinas (UNICAMP), Campinas, São Paulo, Brazil
Abstract
Seed coat color is probably a determinant factor in the antibiosis mechanism of developing resistance to safflower fly (Acanthiophilus helianthi). The purpose of this study was to determine the relationship between the phytochemical content of safflower (Carthamus spp.) seed coat extract with the seed coat color and the damage caused by safflower fly. To this end, germplasm consisting of the cultivated species (C111, with a white seed coat), the wild species (Glaucus and lanatus with a black seed coat; Azar with a brown seed coat), and a breeding line (A82 with a black seed coat) was formed. After cultivating the genotypes, the seed loss (%) and its relationship with the polyphenolic compounds and Cyanidin-3-glucoside of the seed coat extract were examined. Agricultural and phytochemical trait data were analyzed through a completely random block design. With a significant difference from other samples, a minimum damage percentage was observed in Lanatus, Glaucus, and A82 genotypes with a black coat (P < 0.05). The concentration of phenolic compounds, that is, chlorogenic acid, caffeic acid, and p-coumaric acid, except for ferulic acid, was almost equal in all genotypes. However, there was an inverse and direct relationship between the concentration of four polyphenolic compounds (rutin, apigenin, quercetin, and ferulic acid) and Cyd-3-glu content with resistance safflower fly, respectively. In general, flavonoid compounds, that is, rutin, quercetin, and apigenin, affect the resistance probably through antibiosis mechanism so that there was a negative relationship between the concentration of these compounds and resistance to safflower fly.
Key words: antibiosis, flavonoid, safflower, safflower fly
*Corresponding author: Karami Soraya, Department of Agricultural Science, Payame Noor University, Tehran, Iran. Email: [email protected]
Submitted: 19 December 2020; Accepted: 24 March 2021; Published: 3 June 2021
© 2021 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Practical applications
This research shows the inhibitory effects of polyphenolic compounds on the damage caused by safflower fly (Acanthiophilus helianthi) in Carthamus spp. A minimum damage percentage was observed in Lanatus, Glaucus, and A82 genotypes with a black coat. Besides, it seems that the chemical compounds of safflower seed color, particularly Cyd-3-glu and flavonoid compounds such as rutin, quercetin, and apigenin, affect the resistance against safflower fly probably through the antibiosis mechanism.
Introduction
Safflower (Carthamus tinctorius L.) is one of the plants used as cooking oil and poultry feed (Chakradhari et al., 2020). The safflower’s oil content ranges between 29 and 34% and is considered a quality oil given its bright color, high iodine index, and pleasant taste (Ergönül and Özbek, 2020; Rezig et al., 2019). Also, safflower oil is rich in linoleic and oleic acids, which are ingredients for eicosanoids generation. They have anti-blood-pressure and vasodilation effect and adjust the immune system (Mani et al., 2020). Despite all of the plant’s nutrient and medical values, it has received less attention recently from an economic viewpoint as a strategic plant. One reason for this is the decline in safflower seed productivity due to a specific pest known as safflower fly (Acanthiophilus helianthin). The larvae of the insect feed on the flower’s leaflet and can cause further damage to the seed. In addition to a decrease in oil content in the contaminated seeds (about 37.8%), it ultimately damages the seed (Ashri, 1971). Chemical pesticides are not efficient ways to fight the pest given its multi-host and polyphagia nature and its spawning method (Ashti and Efron, 1964). It appears, therefore, that using resistant genotypes is the best way to control the pest. The wild genotypes like C. glaucus, C. palaestinus, C. tenuis, and C. oxyacanthus are resistant to the fly (Ashri, 1971). Recently, a novel, advanced, and uniform line of safflower with a black seed coat and resistance to safflower fly (A82) was obtained via interspecific hybridization of C. tinctorius * C. oxyacanthus following backcrossing and selfing programs (Sabzalian M.R., pers. comm.) (Sabzalian et al., 2010).
Resistance mechanism in plants toward insects is categorized as antixenosis and antibiosis (Painter, 1951). Antixenosis mechanism is one of the morphological specifications that keep insects away from the host plant (Shim et al., 2005). Specific structures of the seed-like wool, wax, surface wax, fiber coats, rigidness, and color are the morphological specifications of the antixenosis mechanism (War et al., 2012). Using an antibiosis mechanism, the plant produces biochemical compounds that negatively affect the insect’s growth, reproduction, and survival (Powell et al., 2006). Chemical compounds are the most common compounds involved in developing resistance through antibiosis mechanisms in plants. There are five groups of these chemical compounds: terpenoids, quinones, alkaloids, glucosinolates, and flavonoids.
Anthocyanins are a group of water-soluble flavonoids with antioxidant and anti-inflammatory activities without side effects (Myung and Hwang, 2008). Anthocyanins in some plants may also be defensive molecular and other flavonoids in plants’ resistance to herbivores and pathogens (Harborne and Williams, 1995). For instance, Johnson et al. reported that insects feeding on floral tissues of Petunia hybrida could synthesize many different anthocyanins (Johnson et al., 2008). In addition, this researcher proposed that delphinidin-3-glucoside could reduce fall armyworms and cabbage loopers while adding cyanidin-3- glucoside to fall armyworms diet could significantly inhibit growth of these insects (Johnson et al., 2008). On the other hand, Costa-Arbulu et al. suggested that anthocyanins produced by Sorghum halepense after aphid of Sipha flava infestation could reduce the aphid fecundity (Costa-Arbulu et al., 2001). All these compounds belong to many secondary metabolites (Way and Van Emden, 2000). Some non-nutrient compounds in the seed, like lectins proteinase inhibitors and alpha-amylases inhibitors, affect this mechanism (Beneke, 2010). In the case of endurance mechanism, while the plant is inflicted, it keeps growing without a considerable loss of strength and yield, although it is damaged to some extent (Weber et al., 2020). Some authors believe that some plants use only one defense mechanism, while two or three mechanisms are engaged (Nalam et al., 2019).
There has been no report for antixenosis defense mechanism in safflower against safflower fly. However, Imrie and Knowles (1970) claimed that the seed color could create resistance to seed bugs. Our previous work showed that colored seed coats, compared to the white coat, tend to be thicker with higher density. Based on this finding, the authors proposed that seed coat color could be involved in the resistance against safflower fly by affecting the morphological structure and rigidity of the seed coat (Karami et al., 2017). Also, there are reports that chemical compounds of seed color via antibiosis mechanisms play a role in developing resistance against insects’ herbivory (Winkel-Shirley, 2001).
Despite the significant relationship between the seed coat color and resistance to safflower fly, limited studies have been carried out on phytochemical content differences in white and colored seeds and damage rate. Thus, the present study is an attempt to examine the relationship between the phytochemical content of safflower seed extract and the seed coat color and the damage level caused by safflower fly.
Materials and Methods
Plant samples
Plant material included seeds of five genotypes of safflower. Three genotypes belonged to three species of wild safflower (one genotypes from C. oxyacanthus, “Azar”, one from C. lanatus, “Lanatus”, one from C. glaucus, “Glaucus”), and two genotypes consisted of cultivated safflower “C111 and A82 “. (Table 1). The breeding line A82 is an advanced generation from C. tinctorius * C. oxyacanthus with a black seed coat (Karami et al., 2017). The female parent of breeding line A82 of C111 genotype has a white seed coat, and the male parent of wild genotype Isfahan II has a black seed coat (Karami et al., 2017).
Table 1. Genotype information and mean score of the traits under study.
| Species | No. | Genotype | Country | Seed coat color | Seeds/uninfested head | Seeds/infested head | Seed loss/head (%) |
|---|---|---|---|---|---|---|---|
| C. tinctorius | 1 | A82 | Iran | Glossy black | 38.8 ± 0.1 | 37.5 ± 0.0 | 3.35 ± 0.4c |
| C. tinctorius | 2 | C111 | Iran | Pure white | 51.8 ± 0.0 | 43.2 ± 0.0 | 16.7 ± 0.5a |
| C. oxyacanthus | 3 | Azar | Iran | Dark brown | 5.5 ± 0.0 | 4.8 ± 0.0 | 11.9 ± 0.3b |
| C. lanatus | 4 | Lanatus | Iran | Black | 11.6 ± 0.7 | 11.4 ± 0.1 | 1.72 ± 0.4d |
| C. glaucus | 5 | Glaucus | – | Black | 8.9 ± 0.1 | 8.8 ± 1.0 | 1.12 ± 0.0d |
Resistance in a choice test
To study the resistance in the introduced genotypes, a choice test was carried out as described in Davis (1985) with a few modifications in the randomized complete block design with three replications in the research farm, Isfahan University of Technology, and Lavark-Najafabad (22°32’E latitude, 51°32’N longitude, 1630 m above the sea level). Each block contained four rows with a 50 cm gap and 2 m length. To synchronize the plant’s encounter with maximum safflower fly attack, the plantation was done on June 5th. The plants were irrigated twice a week until the development of seedlings followed by an irrigation plan based on the plants’ need. In the stemming stage, weeds were removed manually. To examine the damage caused by safflower fly, the host was selected arbitrarily. To determine intra/inter-species resistance in the unit of area, the number of seeds in contaminated and noncontaminated heads and seed generation decline percentage in heads was measured.
Cyd-3-Glu content
The seed coat extract was acidified using a methanolic solution following Siegelman and Hendricks’ instruction (Siegelman and Hendricks, 1958). Then, Cyd-3-Glu content in the seed coat extract was measured based on dry weight (mg/g) and pH difference method using two buffer systems including chloride potassium (pH = 1) and sodium carbonate (pH = 4.5). The procedure was based on Gusty and Rolestad’s method using a spectrophotometer (530–700 nm).
Identification and quantification of polyphenolic compounds of seed coat extract using the HPLC method
To identify and quantify polyphenolic compounds, the seed coat’s methanolic extract was extracted first, and components using High-Performance Liquid Chromatography (HPLC) analysis (Shimadzu, Tokyo, Japan) were used. To prepare the methanolic extract, 10 g of dry powder of seed coat and methanol-water solution (80%) were kept at ambient temperature for 24 h. Afterward, the extracted essence was dried entirely using a rotary set in a vacuum condition and kept in methanol solution (80%) for further analysis. Then, following Lin and Harnly method, polyphenolic compounds were quantified using HPLC (Shimadzu, Tokyo, Japan) [21]. The injection volume was 50 µL, and a chromatograph was prepared (260 & 330 nm). Identification of phenolic and flavonoid compounds of seed coat extracts was performed depending on the retention time (RT) of each one compared with those of pure standards. The results were expressed as mg per 100 g of sample dry weight. The standards followed in this study included flavonoid compounds, namely, rutin, apigenin, and quercetin, and phenolic compounds, namely, p-Coumaric acid, ferulic acid, chlorogenic acid, and caffeic acid. According to Lin and Harnly, after performing sloping chromatography at a speed of 1 mm per min, the chromatogram was obtained by HPLC manager software, and then the curves were calculated and interpreted.
Statistical analysis
Data analysis of agricultural and phytochemical traits in a randomized complete block designwas performed by using GLM procedure of SAS statistical program (version 9.1; SAS Institute Inc., Cary, NC, USA). The mean scores of the traits were compared using the lowest significant difference (LSD). To normalize the data, agricultural traits data were converted. Principal component analysis was done to identify the internal relationship between the genotypes and the measured traits. In addition, traits and genotypes biplots were developed using Stat Graphics.
Results and Discussion
Resistance assessment
The results obtained from genotypes under study in the choice tests are listed in Table 1. As shown, the mean number of seed count in the contaminated and non-contaminated heads was significantly higher in the breeding genotypes (C111 and A82) compared to the wild genotypes (viz., Glaucus, Lantus, and Azar). A comparison among the genotypes in terms of damage to seeds showed that the damage was significantly lower in the wild genotypes. Among them, Lanatus and Glaucus genotypes, with dark seed coat color, had the lowest damage level. A comparison between the breeding genotypes showed that A82 with a black seed coat had a significantly lower level of damage to seeds than C111 with a white seed coat.
It appears that the seed coat color of safflower affects the damage caused by safflower fly. Consistent with our findings, Sabzalian et al. reported that C. oxyacanthus mixed with white and brown-black seed coats had significantly lower damage (0.08%) compared to white coat seeds (17.7%) (Sabzalian et al., 2010). It appears that seeds with colored coats, compared to those with a white coat, are less desirable for the safflower fly. Therefore, the genotypes with colored seed coats receive minor damage than the genotypes with a white seed coat. Probably, the color compounds in the seed coat affect the nutrient behavior of the insect. Imrie and Knowles reported that seeds with a colorful coat in the wild genotypes were more resistant to seed bugs than seeds with a white coat (Imrie and Knowles, 1970). Lale and Kartay highlighted the importance of corn seed color (Zea mays) and its role in resistance to Sitophilus zeamais and S. oryzae contamination after harvest (Lale and Kartay, 2006).
The seeds with colored coats decreased the damage caused by the safflower fly. Therefore, the coat color of the seed can be considered as an index for identifying resistant genotypes. The coat color is a physical and antibiosis factor to resist safflower fly. However, studies have shown that except for color, the rigidness, flavonoid and phenol compounds of seed coat can improve pests’ resistance and damage. For instance, C. palaestinus (white seed coat) and C. oxyacanthus (black seed coat) are the same in terms of rigidity and thickness of the coat; however, the damage sustained by the former is significantly lower (Karami et al., 2017). Therefore, the seed coat’s chemical compounds, such as polyphenolic compounds, decrease the desirability of the seed for seed bugs, which decreases the damage.
Cyd -3-glu content
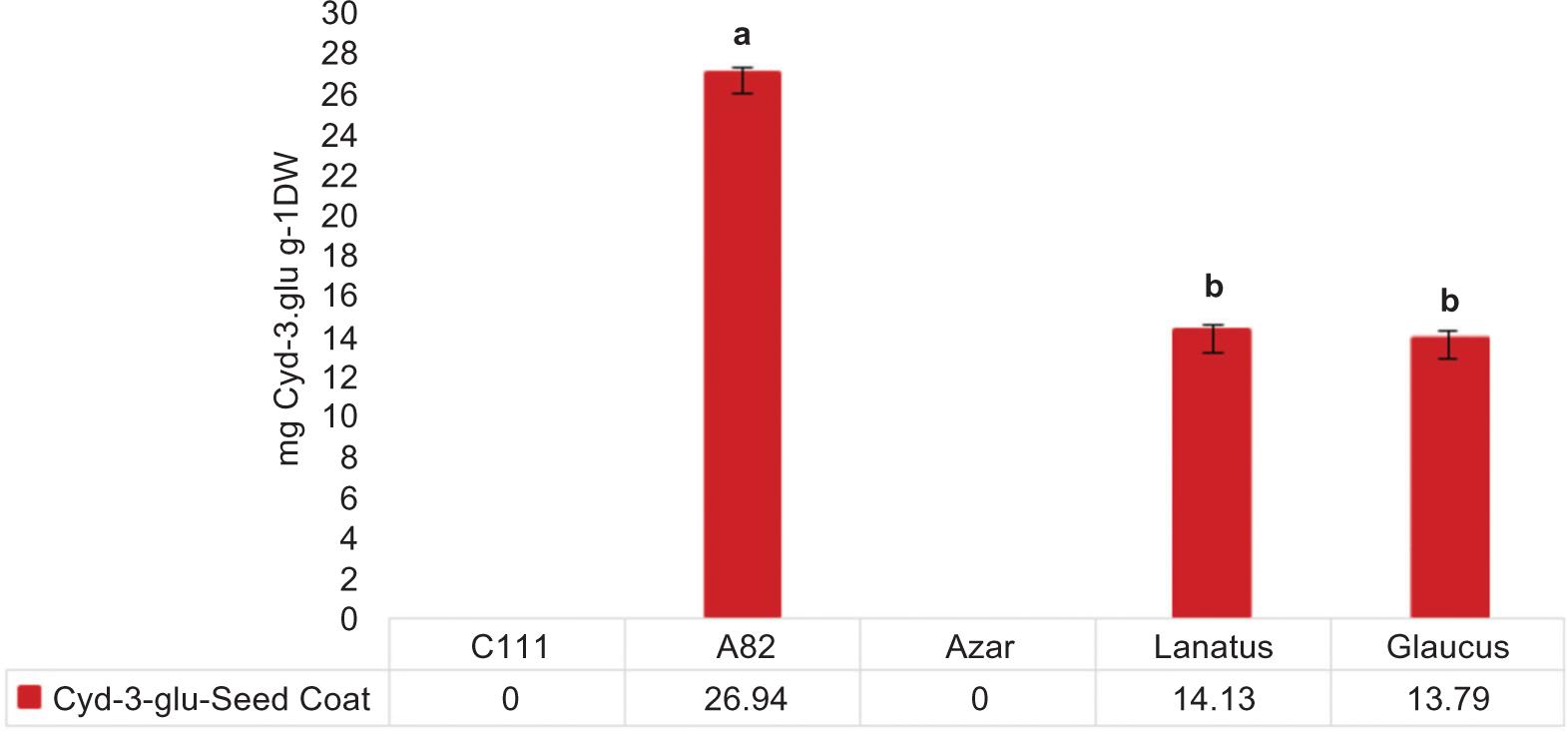
As shown in Figure 1, no Cyd-3-glu was found in extracting the C111 genotype seed coat with white color and the Azar seed coat with brown color. Monomer anthocyanin was found in the seed coat of genotypes with a black color (Glaucus, Lanatus, A82). Among these, A82 had a significantly higher anthocyanin content compared to the other two genotypes. Moreover, there was no significant difference between Glaucus and Lanatus genotypes.
Figure 1. Mean Cyd-3-glu concentration in the seed coat extract of the genotypes under study (g/dry weight).
Other studies about other plants have highlighted the correlation between anthocyanin and seed coat color. For instance, Choung et al. compared soybeans with black, green, and yellow coats and found that anthocyanin was only found in soybeans with black coats, and the soybeans with yellow and green colors lacked anthocyanin (Choung et al., 2001). Lack of Cyd-3-glu in the genotype under study can be associated with a lack of biosynthesis path genes. As noted earlier, one of the hypotheses introduced in this study was the potential role of polyphenolic compounds in seed coat and a decrease in the seed damage caused by the safflower fly. The results supported the hypothesis that genotypes A82, Glaucus, and Lanatus contained Cyd-3-glu and had the lowest level of seed damage than the genotypes without Cyd-3-glu. Few other studies on other plants also support the hypothesis, so that accumulation of the secondary metabolites in different parts acts as a defense mechanism against biological and non-biological stresses. For instance, it is found that the accumulation of flavonoids in leaves of wheat is probably related to resistance to drought stress (Ma et al., 2014). Warren et al. found that high accumulation of flavonoids on Populus leaves created defense mechanisms to UV radiation stress (Warren et al., 2003). Accumulation of flavonoids on the seed of Calotropis procera affected pest control in post-harvest contamination. In addition, flavonoid compounds and epicatechin, particularly in Glycine soja, have a positive and significant relationship with seed coat rigidness (Zhou et al., 2010).
Polyphenolic content of seed coat extract
A chromatography result of phenolic and flavonoid compounds is listed in Table 2. As shown, the level of all phenolic compounds (including chlorogenic acid, caffeic acid, p-coumaric acid) except for ferulic acid was the same in all genotypes under study. On the other hand, flavonoid compounds’ level including quercetin and apigenin was different between genotypes with black seed coat (Glaucus, Lanatus, and A82) and the genotypes with white seed coat (C111) and brown seed coat (Azar). In addition, the level of flavonoid compounds, compared to phenolic compounds, was notably higher so that rutin was the most abundant flavonoid compound followed by apigenin and quercetin.
Table 2. Phenolic and flavonoid compounds in the seed coat extract of genotypes under study (mg/100 g dry weight).
| Genotype | Chlorogenic acid | Caffeic acid | p- Coumaric acid | Rutin | Ferulic acid | Quercetin | Apigenin |
|---|---|---|---|---|---|---|---|
| C111 | 3.01 | 7.19 | 8.68 | 111.99 | 29.80 | 14.10 | 64.88 |
| Azar | 3.36 | 6.88 | 9.54 | 117.95 | 31.24 | 11.27 | 53.96 |
| Lanatus | 3.30 | 6.85 | 8.41 | 50.24 | 14.88 | 3.29 | 7.88 |
| Glaucus | 3.34 | 8.35 | 9.53 | 7.23 | 4.49 | 3.09 | 4.37 |
| A82 | 2.62 | 7.16 | 7.74 | 43.67 | 13.30 | 5.96 | 3.63 |
| RT | 13.95 | 14.30 | 26.97 | 29.32 | 29.89 | 51.6 | 56.27 |
Given the hypothesis proposed in this study about the role of flavonoid and phenolic compounds in the resistance to the safflower fly, the concentration of flavonoid compounds in the study was notably lower in the genotypes with a black seed coat compared to those with white and brown seed coats. In other words, there is a relationship between resistance to the safflower fly and flavonoid compounds, and this relationship is negative. That is, more resistive genotypes have a lower level of flavonoid compounds. It appears that the higher concentration of flavonoid compounds in the genotypes with white and brown genotypes increased susceptibility to the safflower fly.
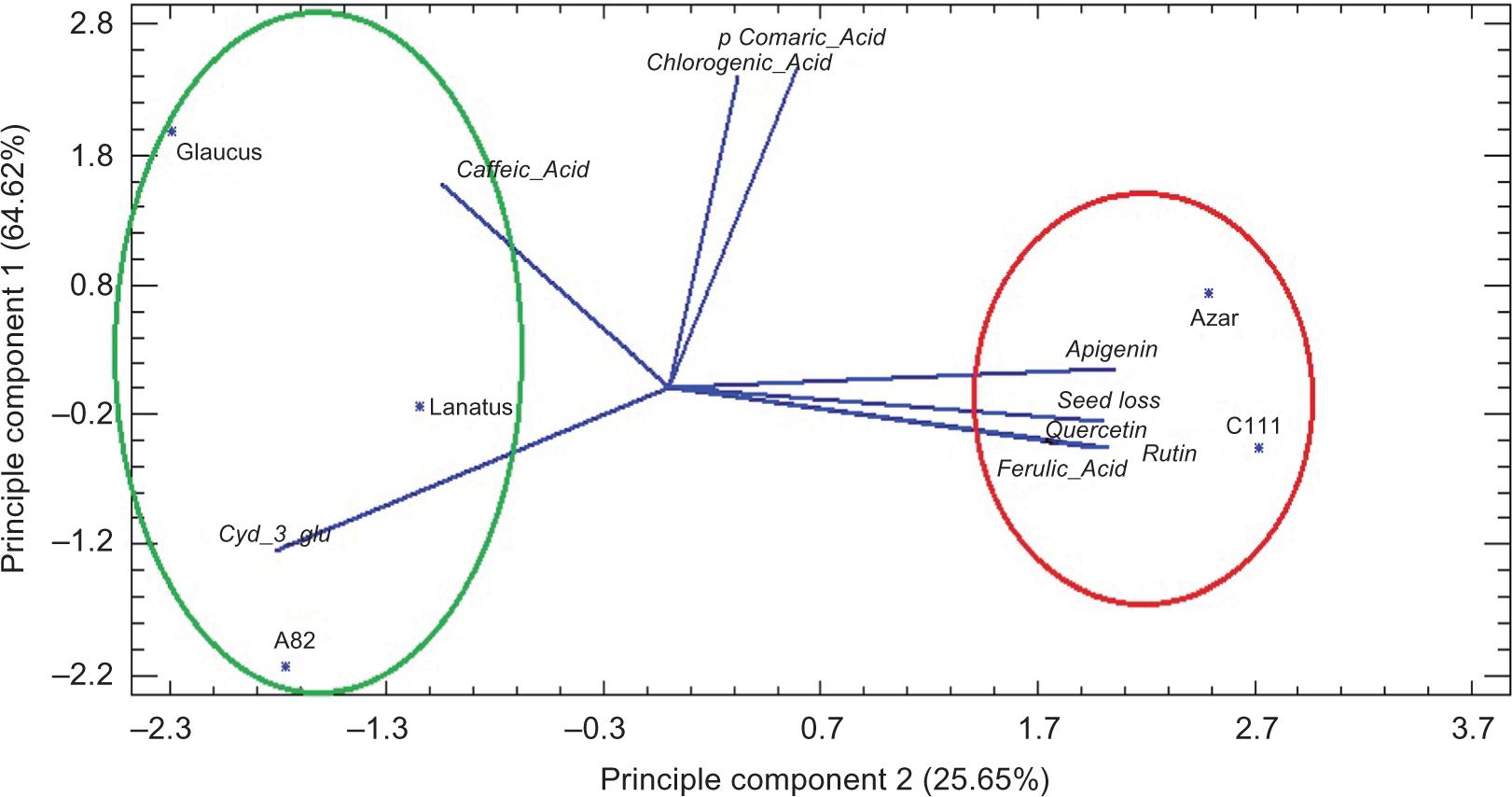
PCA analysis (Principal Component Analysis) was conducted on the selected genotypes and phenolic and flavonoid compounds (Figure 2). The results showed that 64.62% of the total variance was attributed to the first two elements. The first elements had a strong positive relationship with seed yield decrease, rutin, quercetin, and apigenin. Based on genotypes grouping using the biplot of the two first elements, the genotypes with white and brown seed coats (C111 and Azar) formed a separate group of black seed coat genotypes (Glaucus, Lanatus, and A82), characterized by high seed loss, rutin, quercetin, apigenin, and ferulic acid.
Figure 2. The main elements of the genotypes under study based on the measured phenolic and flavonoid compounds.
On the other hand, the genotypes with black seed coats (A82, Lanatus, and Glaucus) were the second independent group. In summary, the PCA results indicated that it is possible to distinguish between resistant and nonresistant genotypes to the safflower fly based on seed yield decrease and flavonoid compounds.
A probable reason for the inverse relationship between flavonoid compounds’ level and resistance to the safflower fly is insects’ skeleton structure and the cuticle formation. At the same time, the light and the rigid skeleton of insects are mostly made of specialized protein, one of the main reasons for the success of insects in the process of evolution. Cuticle resistance depends on the number of reciprocal bonds between protein molecules. That is why insects are interested in polyphenolic compounds, as they constitute 20% of their total body insects (Neville et al., 1975). Among polyphenolic compounds, chlorothe genic acid and gallic acid have more active roles (Brunet, 1980). However, these findings are inconsistent with reports about the toxicity of polyphenolic compounds and their role in plants’ defense mechanisms (Todd et al., 1971). To explain this inconsistency, Anderson (1985) showed that in a diet with a normal concentration of polyphenol, major herbal parts of polyphenols that enter the digestive system of insects are used for cuticle strength. Insects use aromatic compounds of toxic secondary metabolites such as polyphenolic compounds to produce aromatic amino acids. These aromatic compounds have a role in the physiology of insects and the strength of the cuticle structure. However, in a low-protein and low-phenol (plain) diet, flavones (a class of polyphenolic compounds) convert amino acids of the digestive system in insects into cutin, which is toxic and kills the insect. Normal and low-protein (low-phenol) diets can be considered similar to flavonoid compounds in the susceptible (white and brown seed coats) and resistance (black seed coat) genotypes of the safflower.
Conclusion
It seems that the safflower seed coat color probably is involved in developing resistance to the safflower fly through antixenosis and antibiosis mechanisms. This trait can be used to identify the genotypes resistant to the safflower fly. Flavonoid compounds, such as rutin, quercetin, and apigenin, particularly affect the plant’s resistance through an antibiosis mechanism so that there is an inverse relationship between the concentration of these compounds and resistance of the plant. The concentration of these compounds in resistant genotypes with a black seed coat was significantly lower than that of the non-resistant genotypes with white and brown seed coats. In addition to the effect of flavonoid compounds on resistance to the safflower fly, it appears that Cyd-3-glu probably had a role in developing resistance in the plant through the antibiosis mechanism. This compound was only found in the resistant genotypes with a black seed coat.
REFERENCES
Anderson, S. O. 1985. Sclerouzation and tanning of the cuticle. In: Comprehensive Insect Physiology, Biochemistry and Pharmacology. Kerkut, G.A. and L.I. Gilbert (eds.). Oxford, Pergamon Press, 59–74 pp.
Ashri, A., 1971. Evaluation of the world collection of safflower, carthamus tinctorius LI reaction to several diseases and associations with morphological characters in Israel 1. Crop Science 11(2): 253–257. 10.2135/cropsci1971.0011183X001100020026x
Ashti, A. and Efron, Y., 1964. Inheritance studies with fertile interspecific hybrids of three carthamus L. species 1. Crop Science 4(5): 510–514. 10.2135/cropsci1964.0011183X000400050023x
Beneke, C.J., 2010. The expression and inheritance of resistance to acanthoscelides obtectus (Bruchidae) in South African dry bean cultivars. University of the Free State, South Africa, Bloemfontein, 1–97.
Brunet, P.C., 1980. The metabolism of the aromatic amino acids concerned in the cross-linking of insect cuticle. Insect Biochemistry 10(5): 467–500. 10.1016/0020-1790(80)90082-7
Chakradhari, S., Perkons, I., Mišina, I., Sipeniece, E., Radziejewska-Kubzdela, E., Grygier, A., Rudzińska, M., Patel, K.S., Radzimirska-Graczyk, M. and Górnaś, P., 2020. Profiling of the bioactive components of safflower seeds and seed oil: cultivated (Carthamus tinctorius L.) vs. wild (Carthamus oxyacantha M. Bieb.). European Food Research and Technology 246(3): 449–459. 10.1007/s00217-019-03414-w
Choung, M.-G., Baek, I.-Y., Kang, S.-T., Han, W.-Y., Shin, D.-C., Moon, H.-P. and Kang, K.-H., 2001. Isolation and determination of anthocyanins in seed coats of black soybean (Glycine max (L.) Merr.). Journal of Agricultural and Food Chemistry 49(12): 5848–5851. 10.1021/jf010550w
Costa-Arbulú C., Gianoli E., Gonzáles M.L. and Niemeyer H.M., 2001. Feeding by the Aphid Sipha flava produces a reddish spot on leaves of Sorghum halepense: an induced defense. Journal of Chemical Ecology 27(2): 273–283. 10.1023/A:1005676321251
Davis, F.M., 1985. Entomological techniques and methodologies used in research programs on plant resistance to insects. Insect Science and Its Application 6: 391e400. 10.1017/S1742758400004689
Ergönül, P.G. and Özbek, Z.A., 2020. Cold pressed safflower (Carthamus tinctorius L.) seed oil. Cold Pressed Oils 323–333. 10.1016/B978-0-12-818188-1.00029-3
Harborne, J.B. and Williams, C.A., 1995. Anthocyanins and other flavonoids. Natural Product Reports 12(6): 639–657. 10.1039/np9951200639
Imrie, B. and Knowles, P., 1970. Inheritance studies in interspecific hybrids between carthamus flavescens and C. tinctorius 1. Crop Science 10(4): 349–352. 10.2135/cropsci1970.0011183X001000040009x
Johnson, E.T., Berhow, M.A. and Dowd, P.F., 2008. Clored and white sectors from star-patterned petunia flowers display differential resistance to corn earworm and cabbage looper larvae. Journal of Chemical Ecology 34(6): 757–765. 10.1007/s10886-008-9444-0
Karami, S., Sabzalian, M.R., Rahimmalek, M., Saeidi, G. and Ghasemi, S., 2017. Interaction of seed coat color and seed hardness: an effective relationship which can be exploited to enhance resistance to the safflower fly (Acanthiophilus helianthi) in Carthamus spp. Crop Protection 98: 267–275. 10.1016/j.cropro.2017.04.006
Lale, N. and Kartay, M., 2006. Role of physical characteristics of the seed in the resistance of local cultivars of maize to Sitophilus zeamais infestation in storage. Tropical Science 46(1): 1–3. 10.1002/ts.5
Ma, D., Sun, D., Wang, C., Li, Y. and Guo, T., 2014. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiology and Biochemistry 80: 60–66. 10.1016/j.plaphy.2014.03.024
Mani, V., Lee, S.-K., Yeo, Y. and Hahn, B.-S., 2020. A metabolic perspective and opportunities in pharmacologically important safflower. Metabolites 10(6): 253. 10.3390/metabo10060253
Myung, J.E. and Hwang, I.K., 2008. Functional components and antioxidative activities of soybean extracts. Korea Soybean Digest 25: 23–29.
Nalam, V., Louis, J. and Shah, J., 2019. Plant defense against aphids, the pest extraordinaire. Plant Science 279: 96–107. 10.1016/j.plantsci.2018.04.027
Neville, A. C. 1975. Biology of insect cuticle. Springer-Verlag, Berlin, 448 pp.
Painter, R.H. 1951. Insect resistance in crop plants. Macmillan, New York.
Powell, G., Tosh, C.R. and Hardie, J., 2006. Host plant selection by aphids: behavioral, evolutionary, and applied perspectives. Annual Review of Entomology 51: 309–330. 10.1146/annurev.ento.51.110104.151107
Rezig, L., Sadaa, M., Trabelsi, N., Tammar, S., Limam, H., Rebey, I.B., Smaoui, A., Sghaier, G., Del Re, G. and Ksouri, R., 2019. Chemical composition, antioxidant and antimicrobial activities of Aloysia Triphylla L. essential oils and methanolic extract. Italian Journal of Food Science 31(3): 591–603. 10.14674/IJFS-1373
Sabzalian, M.R., Saeidi, G., Mirlohi, A. and Hatami, B., 2010. Wild safflower species (Carthamus oxyacanthus): a possible source of resistance to the safflower fly (Acanthiophilus helianthi). Crop Protection 29(6): 550–555. 10.1016/j.cropro.2009.12.013
Shim, K., Lee, Y., Cho, S., Pae, S. and Suh, D., 2005. Inheritance of seed coat color in sesame. Korean Journal of Breeding 37(1): 1–4. 10.1590/S0100-204X2014000400007
Siegelman, H.W. and Hendricks, S.B., 1958. Photocontrol of anthocyanin synthesis in apple skin. Plant Physiology 33(3): 185. 10.1104/pp.33.3.185
Todd, G.W., Getahun, A. and Cress, D.C., 1971. Resistance in barley to the greenbug, Schizaphis graminum. 1. Toxicity of phenolic and flavonoid compounds and related substances. Annals of the Entomological Society of America 64(3): 718–722. 10.1093/aesa/64.3.718
War, A.R., Paulraj, M.G., Ahmad, T., Buhroo, A.A., Hussain, B., Ignacimuthu, S. and Sharma, H.C., (2012). Mechanisms of plant defense against insect herbivores. Plant Signaling & Behavior 7(10): 1306–1320. 10.4161/psb.21663
Warren, J.M., Bassman, J.H., Fellman, J.K., Mattinson, D.S. and Eigenbrode, S., 2003. Ultraviolet-B radiation alters phenolic salicylate and flavonoid composition of Populus trichocarpa leaves. Tree Physiology 23(8): 527–535. 10.1093/treephys/23.8.527
Way, M. and Van Emden, H., 2000. Integrated pest management in practice—pathways towards successful application. Crop Protection 19(2): 81–103. 10.1016/S0261-2194(99)00098-8
Weber, D., Egan, P.A., Muola, A., Ericson, L.E. and Stenberg, J.A., 2020. Plant resistance does not compromise parasitoid-based biocontrol of a strawberry pest. Scientific Reports 10(1): 1–10. 10.1038/s41598-020-62698-1
Winkel-Shirley, B., 2001. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiology 126(2): 485–493. 10.1104/pp.126.2.485
Zhou, S., Sekizaki, H., Yang, Z., Sawa, S. and Pan, J., 2010. Phenolics in the seed coat of wild soybean (Glycine soja) and their significance for seed hardness and seed germination. Journal of Agricultural and Food Chemistry 58(20): 10972–10978. 10.1021/jf102694k