
Download




Original Article
Kirenol inhibits TNF-α-induced proliferation and migration of HaCaT cells by regulating NF-κB pathway
Jin Li, Fang Ren, Wenliang Yan, Hong Sang*
Department of Dermatology, The Affiliated Jinling Hospital of Nanjing Medical University, Nanjing, Jiangsu, China
Abstract
Psoriasis is a common chronic, inflammatory skin disease possessing properties of inflammatory cell infiltration and excessive proliferation of keratinocytes, the occurrence and development of which remain fully elucidated. Therefore, the study was designed to determine the effects of kirenol (50, 100 and 200 μg/mL) on Cultured Human Keratinocytes (cells) (HaCaT) in vitro and reveal its molecular mechanism. The in vitro psoriasis model was established utilizing tumor necrosis factor-α (TNF-α)-stimulated HaCaT cells. Kirenol, a diterpenoid compound, was applied at different concentrations (50, 100 and 200 μg/mL) to HaCaT cells for 24 h. The Cell Counting Kit-8 (CCK-8) and thymidine monobromodeoxyuridine (BrdU) assays were used to assess cell viability and proliferation, followed by assessment of cell migration by Transwell assay. Subsequently, inflammatory cytokines were measured by enzyme-linked immunosorbent assay (ELISA), and Western blot assay was used to evaluate expressions of p65, p-p65, IκBα and p-IκBα. Activities of superoxide dismutase (SOD), catalase (CAT), glutathione (GSH) and malondialdehyde (MDA) contents were measured spectrophotometrically. The results demonstrated that TNF-α induced a significant increase in cell viability and inflammatory cytokines, including expressions of Interleukin (IL)-6, IL-8, IL-22 and IL-1β in HaCaT cells, which was dose-dependently inhibited by kirenol. Similarly, TNF-α-induced cell migration was also suppressed by kirenol treatment. Furthermore, TNF-α stimuli induced the upregulation of phosphorylation levels of p65 and IκBα as well as p-p65–p65 and p-IκBα–IκBα ratios, whereas kirenol significantly suppressed the activation of cellular nuclear factor-kappa B (NF-κB) signaling pathway. In addition, kirenol significantly decreased the level of MDA but increased the levels of SOD, CAT and GSH in a dose-dependent manner. These results proposed that kirenol could inhibit the proliferation, migration, expression of inflammatory factors, and oxidative stress in HaCaT cells via suppressing NF-κB signaling pathway.
Key words: kirenol, psoriasis, proliferation, migration, inflammation
*Corresponding Author: Hong Sang, Department of Dermatology, The Affiliated Jinling Hospital of Nanjing Medical University, No. 305 Zhongshan East Road, Xuanwu District, Nanjing, Jiangsu 210029, China. Email: [email protected]
Received: 15 September 2021; Accepted: 27 October 2021; Published: 27 November 2021
© 2021 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Psoriasis is a recurrent, noninfectious erythema scaly skin disease characterized by epidermal proliferation and inflammation. Its etiology is related to heredity, infection, allergy, metabolic disorders, and autoimmunity (Allergologia et immunopathologia, 2020; Aquino et al., 2021; Evans et al., 2020; Srivastava et al., 2021; Zhou et al, 2020). It is a typical chronic inflammatory disease with an increasing incidence ranging from 1 % to 3 % worldwide (Griffiths & Barker, 2007; Hoegler et al., 2018; Langley et al. 2005). At present, psoriasis is an extremely complicated pathophysiologic process, the occurrence and development of which remain unclear (Boehncke & Schön, 2015). Currently, psoriasis is believed to mainly involve genetic, psychological, immunologic, and environmental factors (Yang & Zheng, 2020). Moreover, the principle obsessive progressions from psoriasis hold numerous different features in keratinocyte dysplasia, keratosis, neovascularization as well as inflammatory cell infiltration (Kim et al., 2014a). At the same time, excessive proliferation and abnormal migration of keratinocytes leading to the microenvironment of psoriasis lesions are vital features of psoriasis (Winchell & Watts, 1988). As psoriasis is prone to relapse and challenging to cure, it is conducive to exploring new therapeutic targets related to keratinocyte proliferation as well as migration for the treatment of psoriasis.
As the primary pathogenic cells in psoriasis, keratinocytes is considered as a promoter for amplification of inflammatory response with the additional production of tumor necrosis factor-α (TNF-α) in the initiation of disease (Nickoloff, 2006). Following stimulation with TNF-α, keratinocytes may provoke the abbrent expression of pro-inflammatory cytokines and chemokines such as IL-6, IL-8, IL-22 and IL-1β, which contribute to the subsequent recruitment of immune cells into inflammatory lesions in the skin (Cho, Lee, & Kim, 2007; Lee, Chung, Kang, Park, & Hyun, 2019; Wolk et al., 2009). It is believed that there are multiple signaling pathways involved in the inflammatory response, among which the nuclear factor-kappa B (NF-κB) pathway plays a decisive role (Nair et al., 2009; A. Wang et al., 2019; Xu, Xu, Xiong, & Deng, 2019). During the signaling pathway, inflammatory stimuli activate IKK, promoting phosphorylation and degradation of IκB and subsequent activation of NF-κB signaling pathway, which results in a series of inflammatory responses (Lawrence, 2009; Zhang, Lenardo, & Baltimore, 2017). Recognizing the certainty that NF-κB signaling pathway is involved in the progression of psoriasis, inhibition of the pathway is beneficial to reduce hyperproliferation and hyper inflammation in keratinocytes. Therefore, inflammatory factor (IL-6/IL-8/IL-22/IL-1β) and NF-κB were chosen to investigate the effect of kirenol on psoriasis in this study.
Kirenol is a diterpenoid characteristic product compound extracted from herba siegesbeckiae (Ibrahim et al., 2021; Wang et al., 2021a). In addition, kirenol attenuates the inflammatory response to collagen-induced arthritis by inhibiting the activation of signaling pathways and reducing the expression of pro-inflammatory cytokines such as IL-1β (Wang et al., 2011). Moreover, kirenol inhibits inflammatory factors and improves ulcerative colitis in mice (Xiuhong et al., 2019). Besides, kirenol inhibits rheumatoid arthritis (RA)-associated synovial fibroblasts proliferation, migration, invasion, and inflammation (Wu et al., 2019). Although kirenol has also been reported to have immunomodulatory and anti-inflammatory properties, the efficacy of kirenol on psoriasis has not been further investigated. In the present study, we investigated the effects of kirenol on psoriasis development and its mechanism of function. The results revealed that kirenol significantly inhibited the proliferation capacity and migration ability of Cultured Human Keratinocytes (cells) (HaCaT) through regulating the NF-κB pathway.
Materials and Methods
Cell culture
Cultured human immortalized keratinocytes (HaCaT; Procell, Wuhan, China) were cultured in Dulbecco’s Modified Eagle medium (DMEM), supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin in a humidified incubator at 37°C and 5% CO2. HaCaT cells in a logarithmic growth phase were induced with 10 ng/mL TNF-α for 48 h to establish an in vitro cell model for psoriasis at 37°C and 5% CO2. The kirenol (Yuan-ye Bio-Technology Co. Ltd, Shanghai, China) concentration was increased to 50, 100 and 200 μg/mL for 24 h at 37°C and 5% CO2. Furthermore, the cells were separately divided in the following five groups: the control group; the TNF-α group; the TNF-α + kirenol (50 μg/mL) group, the TNF-α + kirenol (100 μg/mL) group and the TNF-α + kirenol (200 μg/mL) group. Dimethyl sulfoxide (DMSO; Merck, Germany) was used in equal concentrations as a negative control to dilute kirenol.
Cell viability assay
The antiproliferative impact of kirenol concerning HaCaT was determined utilizing Cell Counting Kit-8 reagent (CCK-8; Bestbio, Shanghai, China). For a short period of time, HaCaT cells were inoculated in 96-well plates at a density of 5 × 103 cells/well overnight, treated with different concentrations of kirenol (50, 100, 200 and 400 μg/mL) for 24 h, and incubated with CCK-8 reagent in dark for 1 h at 37°C. After TNF-α treatment for 48 h at 37°C and 5% CO2, kirenol at concentrations of 50, 100, 200 μg/mL was applied to HaCaT cells. The absorbance of the tests was recorded at a wavelength of 450 nm utilizing a Microplate Reader (Infinite M200, Tecan, Austria).
Measurement of proliferation by incorporation of thymidine monobromodeoxyuridine (BrdU)
Cell proliferation was investigated by thymidine monobromodeoxyuridine (BrdU) under de novo-synthesized DNA conditions utilizing the BrdU Cell Proliferation assay kit (Santa Cruz Biotechnology). Briefly, HaCaT cells were seeded into 96-well plates at 2 × 103 cells/well, and incubated at 37°C for 48 h. Subsequently, 20-mL BrdU was added, and incubated at 37°C for 4 h. After incubation with anti-BrdU antibody and peroxidase-conjugated goat anti-mouse immunoglobulin G (IgG), successively, 100-mL 3,3',5,5'-Tetramethylbenzidine (TMB) peroxidase substrate was added and the mixture was kept in dark at room temperature for 30 min. The absorbance at 450/550 nm was obtained using a microplate reader.
Measurement of superoxide dismutase (SOD) activity
The mixture containing 0.033-mM etheylenediaminetetraacetic acid (EDTA), 3.3-mM methionine, 0.33-µg/mL riboflavin and 0.01-mM KCN (Sigma, St. Louis, MO, USA) was applied to supernatant. Cuvette containing the reaction mixture was placed in a box illuminated with 20-W neon lamp for 10 min. One unit of SOD enzyme activity is defined as the amount of enzyme required for inhibiting the chromogen production (optical density at 560 nm) by 50% in 1 min under assay conditions and expressed as specific activity in units/min/mg protein.
Measurement of glutathione (GSH) activity
The GSH level was measured by reacting with 5,5’-dithiobis(2-nitrobenzoic) acid (DTNB; Sigma, St. Louis, MO, USA), producing a yellow-colored compound, 5’-thio-(2-nitrobenzoic) acid (TNBA). Concentration of TNBA was measured at 412-nm wavelength, and the amount of GSH was expressed in terms of nmol GSH/mg protein.
Measurement of catalase (CAT) activity
The CAT activity was determined by splitting H2O2 for 1 min. The reaction was stopped by dichromate/acetic acid reagent, and the remaining H2O2 was measured at 570 nm for chromic acetate. The CAT activity was expressed as μmol/L H2O2 decomposed/min/mg protein.
Measurement of malondialdehyde (MDA) activity
The MDA activity was determined by a Lipid Peroxidation MDA assay kit (Beyotime Institute of Biotechnology, Haimen, China) as recommended by the manufacturer. After treatment with kirenol (50, 100, 200 μg/mL) for 24 h, HaCaT cells were harvested, lysed by cell lysis buffer and centrifuged at 12,000 g for 15 min at 4°C. The obtained supernatant was reacted with thiobarbituric acid, and the reaction products were measured with a microplate reader at 532 nm. Total protein level was determined by BCA protein detection kit (Beyotime Biotechnology Institute, Haimen, China), and MDA level was normalized.
Enzyme-linked immunosorbent assay (ELISA)
The TNF-α-induced HaCaT cells were treated with kirenol (50, 100, 200 μg/mL) for 24 h. Then, inflammatory factors (IL-6, IL-8, IL-22 and IL-1β) in the supernatant were detected by ELISA kit (eBioscience, CA, USA) following the manufacturer’s instructions. Briefly, 50 mL of standard, samples, and blank control were added to a 96-well plate coated with primary antibodies against cytokines, including IL-6 (Cat No. ab178013; Abcam, Cambridge, MA, USA), IL-8 (Cat No. ab214030; Abcam, Cambridge, MA, USA), IL-22 (Cat No. ab216170; Abcam, Cambridge, MA, USA) and IL-1β (Cat No. ab214025; Abcam, Cambridge, MA, USA), followed by incubation with HRP-conjugated detection antibodies (100 mL) for 1 h at 37°C. After washing for five times, substrates A and B were added for further 15-min incubation. Finally, the absorbance was measured at 450 nm in a microplate reader. A standard curve was produced to quantify the concentration of cytokines from different samples.
Transwell assay
To induced cell migration, CoStar Transwell chambers (8-μm pore size; Corning, Costar, NY, USA) were used. TNF-α-induced HaCaT cells (1 × 105/well) were inoculated in the wells of upper chambers filled with 200-μL serum-free DMEM medium, and lower chambers were filled with 500-μL medium, which contained 20% fetal bovine serum. After incubation at 37°C in 5% CO2 incubator for 24 h, cells on the upper surface of polycarbonate membrane were gently removed with a wet cotton swab. Cells that migrated to the lower surface of the membrane were fixed with 4% paraformaldehyde and stained with 1% crystal violet. The number of cells was counted with a microscope (CKX53, Olympus, Japan), and the images were captured.
Western blotting assay
For Western blot analysis, the TNF-α-stimulated HaCaT cells were treated with kirenol (50, 100, 200 μg/mL) for 24 h. Proteins were extracted from the cells grown on 35-mm culture dishes using the Trizol protein isolation method. Bradford’s assay quantified the isolated proteins. The membrane was blocked at room temperature for 2 h and incubated overnight at 4°C with the following primary antibodies: anti-p65 (ab19870, 1:500; Abcam, Cambridge, MA, USA), anti-p-p65 (ab194726, 1:500; Abcam, Cambridge, MA, USA), anti-IκBα (ab247825, 1:1000; Abcam, Cambridge, MA, USA), anti-p-IκBα (9246, 1:500; Cell Signaling, Boston, Massachusetts, USA) and anti-GAPDH (ab8245, 1:1000; Abcam, Cambridge, MA, USA), following brooding for IgG horseradish peroxidase (HRP)-conjugated secondary antibody at room temperature for 1 h. At this point, the particular proteins were distinguished by improved chemiluminescence (Invitrogen, Carlsbad, CA, USA) according to manufacturer’s instructions.
Statistical analysis
The mean ± standard deviation (SD) was calculated for each experimental group. Statistical comparisons of the data were analyzed using the SPSS software (SPSS 26, Chicago, IL, USA). Student’s t-test was used to compare the two groups. Differences between the groups were statistically significant (P < 0.05).
Results
Kirenol restrained TNF-α-induced proliferation of HaCaT cells in a dose-dependent manner
Cultured human keratinocytes were first treated with 10-ng/mL TNF-α for 48 h to simulate the epithelial state of psoriasis. The proliferation of HaCaT cells was detected by CCK-8 and BrdU assays to verify the effects of kirenol on TNF-α-induced proliferation of HaCaT cells. Then HaCaT were incubated with the following gradient concentrations of kirenol: 50, 100 and 200 μg/mL for 24 h. The following five groups were used in the experiment, including the control group: TNF-α group, TNF-α + kirenol (50 μg/mL) group, TNF-α + kirenol (100 μg/mL) group and TNF-α + kirenol (200 μg/mL) group. Figure 1A shows that kirenol concentrations of 50, 100 and 200 μg/mL were nontoxic to HaCaT cells. The CCK-8 assay results indicated that kirenol markedly inhibited the viability of HaCaT cells in a dose-dependent manner in comparison to the control group (Figure 1B). In addition, the BrdU assay illustrated that TNF-α-induced proliferation of HaCaT cells was significantly suppressed by kirenol in a dose-dependent manner compared to the control group (Figure 1C).
Figure 1. Kirenol inhibited TNF-α-induced proliferation of HaCaT cells in a dose-dependent manner. (A) Kirenol at concentrations of 50, 100 and 200 µg/mL was nontoxic to HaCaT cells. (B) Kirenol reduced cell viability in a dose-dependent manner. (C) Kirenol increased BrdU positive cells in a dose-dependent manner. (D) The percentage of cells with BrdU-positive (BrdU+) staining was determined. **P < 0.01 vs. control; ***P < 0.001 vs. control; #P < 0.05, ##P < 0.01 vs. TNF-α alone. TNF-α: tumor necrosis factor-α.
Kirenol improved TNF-α-induced inflammation of HaCaT cells
In order to determine the impact of kirenol (50, 100, and 200 μg/mL) on TNF-α-induced inflammation of HaCaT cells, ELISA was performed to measure inflammatory cytokines associated with the development of inflammation, such as IL-6, IL-8, IL-22 and IL-1β. The ELISA data revealed that compared with the control group, the expressions of inflammation factors were significantly increasing in the TNF-α alone group. In contrast, they were reduced considerably by kirenol in a concentration-dependent manner. The results revealed that kirenol improved TNF-α-induced inflammation of HaCaT cells in a dose-dependent manner (Figure 2).
Figure 2. Kirenol alleviated TNF-α-induced inflammation of HaCaT cells. Kirenol decreased the levels of Interleukin-8 (IL-8), IL-1β, IL-6, and IL-22. ***P < 0.001 vs. control; #P < 0.05 vs. TNF-α alone, ##P < 0.01 vs. TNF-α alone, and ###P < 0.001 vs. TNF-αα alone.
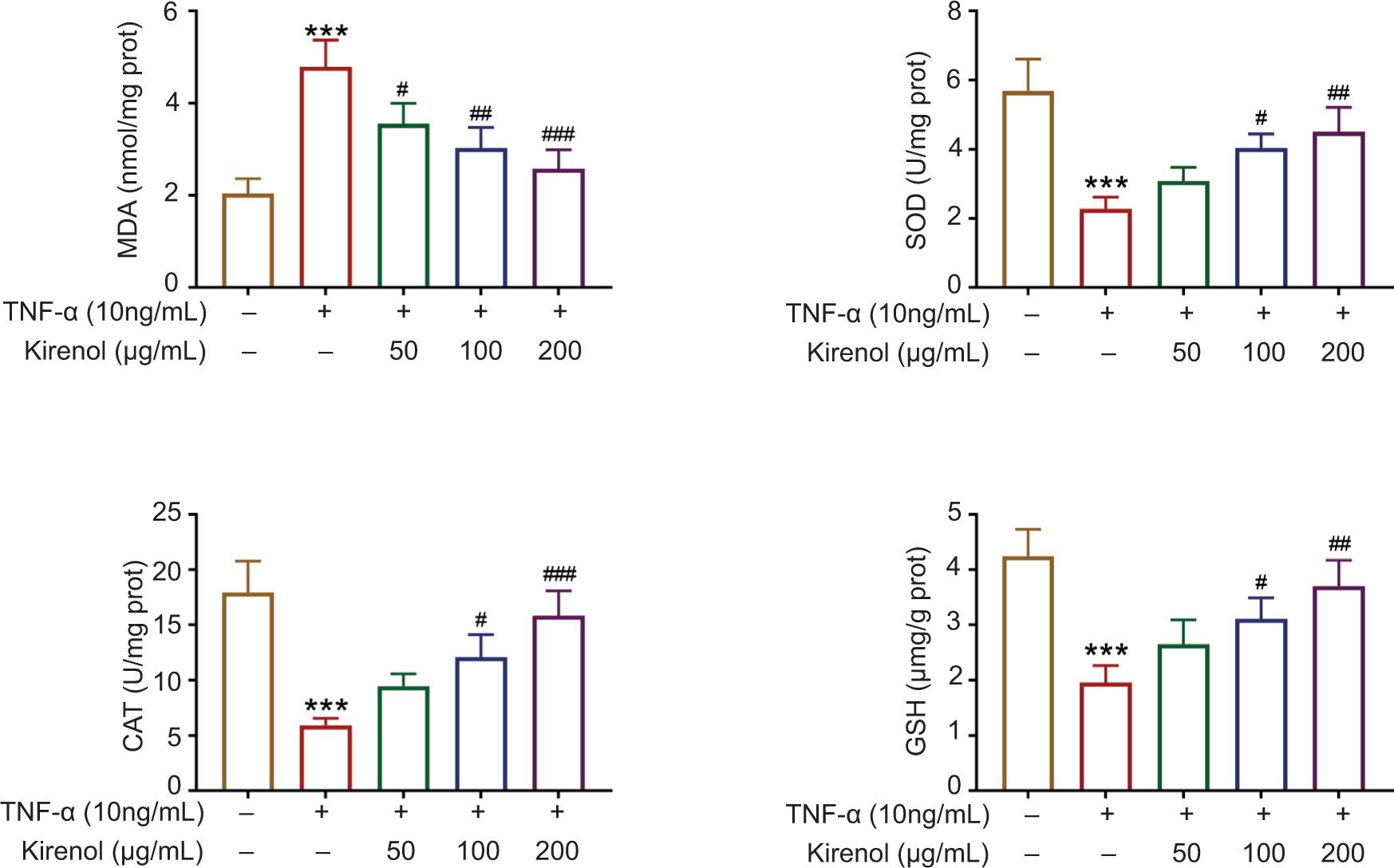
Kirenol inhibited oxidative stress in TNF-α-stimulated HaCaT cells
Oxidative stress is the main factor for psoriasis. Therefore, oxidative stress assay was performed and the results displayed that TNF-α increased MDA but decreased SOD, CAT and GSH, indicating that TNF-α stimuli promoted oxidative stress. However, application of kirenol significantly decreased the level of MDA but increased the levels of SOD, CAT and GSH in a dose-dependent manner (Figure 3). Collectively, kirenol inhibited oxidative stress of HaCaT cells.
Figure 3. Kirenol inhibited the oxidative stress in TNF-α-stimulated HaCaT cells. Kirenol decreased the level of MDA but increased the levels of SOD, CAT, and GSH in a dose-dependent manner. ***P < 0.001 vs. control; #P < 0.05 vs. TNF-α alone, ##P < 0.01 vs. TNF-α alone, and ###P < 0.001 vs. TNF-α alone. TNF-α: tumor necrosis factor-α; MDA: malondialdehyde; SOD: superoxide dismutase; CAT: catalase; GSH: glutathione.
Kirenol suppressed TNF-α-induced migration of HaCaT cells
In order to investigate the role of kirenol (50, 100, and 200 μg/mL) in the process of TNF-α-induced migration of HaCaT cells, we further assessed its migration capacity utilizing Transwell assay. Correspondingly, the Transwell migration assay demonstrated that TNF-α enhanced the migratory ability of HaCaT cells compared to the control group. However, it was markedly suppressed by kirenol in a dose-dependent manner. The results established that kirenol suppressed the TNF-α-induced migration of HaCaT cells in a dose-dependent manner (Figure 4).
Figure 4. Kirenol suppressed TNF-α-induced migration of HaCaT cells. Kirenol inhibited TNF-α-induced HaCaT cell migration, and representative images of the HaCaT cell migration assay. ***P < 0.001 vs. control; ###P < 0.001 vs. TNF-α alone. TNF-α: tumor necrosis factor-α.
Kirenol decreased the activation of NF-κB pathway in TNF-α-stimulated HaCaT cells
NF-κB pathway was related to various pathological and physiological changes such as cell proliferation, immunity as well as inflammation. For one thing, accumulating evidence has been reported that NF-κB pathway exhibits a vital role in TNF-α-induced proliferation and migration of HaCaT cells. For another, the inflammatory factors such as IL-1β and IL-6 critically rely on the activation and nuclear translocation of NF-κB. Therefore, we speculated that kirenol performs its function by regulating the NF-κB pathway. To further validate the molecular mechanism by which kirenol (50, 100, and 200 μg/mL) projects in the cell, we measure the levels of NF-κB pathway-related proteins such as p65, p-p65, IκBα and p-IκBα. The TNF-α-induced HaCaT cells were lysed in RIPA buffer, which contained phosphatase inhibitors as well as protease, and then analyzed by Western blotting assay. The Western blotting assay established that the phosphorylation levels of p65 and IκBα, and p-p65–p65 and p-IκBα–IκBα ratios increased markedly, and kirenol reduced their phosphorylation levels in a dose-dependent manner (Figure 5). This finding demonstrated that kirenol exerted its effects of both inhibiting cell activity and migratory capacity by decreasing the activation of NF-κB pathway.
Figure 5. Kirenol inhibited the activation of NF-κB pathway in TNF-α-induced HaCaT cells. Kirenol prevented TNF-α-induced activation of HaCaT cell in NF-κB signaling pathway. Western blotting assay was performed to detect the expression of p-p65, p65, p-IκBα, and IκBα in HACaT cells. Expression of p-p-65 relative to p65, p-p-IκBα relative to IκBα, and p65 or IκBα relative to GAPDH. ***P < 0.001 vs. control; #P < 0.05 vs. TNF-α alone, ##P < 0.01 vs. TNF-α alone, and ###P < 0.001 vs. TNF-α alone. TNF-α: tumor necrosis factor-α; I-κBα: inhibitor of NF-κB; p-IκBα: phosphorylated (p)-inhibitor of NF-κBα.
Discussion
Psoriasis is an extremely complicated pathophysiological process, and the primary pathological characteristics incorporate the inordinate proliferation of keratinocytes and local inflammatory response (Callis Duffin et al., 2017; Cooper, 1990). Currently, the mechanism of etiology and pathogenesis of psoriasis remain unclear. As reported, kirenol has various medicinal properties, including antibacterial, antiviral, anti-inflammatory, antiarthritic as well as anti-adipogenesis effects ( Kim et al., 2014b; Lu et al., 2012; Ren et al., 2020; Wang et al., 2012). Accumulating evidence has proved the biological effects of kirenol on a wide range of cells. For instance, its ability to inhibit cell proliferation and differentiation induces cell apoptosis and alleviates inflammatory response (Wang et al., 2021b; Wu et al., 2019). However, whether kirenol affects psoriasis remains fully understood. The present study found that kirenol inhibited TNF-α-induced phenotypes of HaCaT cells by modulating the NF-κB pathway.
TNF-α-induced inflammation had been previously proved (Choi & Hwang, 2019). It has been reported by a previous study that TNF-α stimulation considerably improved the viability of HaCaT cells and upregulated inflammatory factors such as IL-8, IL-1β, IL-6 and IL-22 (Na Takuathung et al., 2017). A recent study has found that the levels of cytokines, including IL-1α/β, IL-6 and IL-8, were remarkably upregulated after TNF-α stimuli, indicating that TNF-α induced significant inflammatory response in HaCaT cells. Mevastatin was also demonstrated to alleviate inflammation ( Kim et al., 2021). Meanwhile, the expression of inflammatory factors in the supernatant of TNF-α-stimulated cells was significantly down-regulated by kirenol. The results of ELISA assay were consistent with the findings of previous studies, which indicated an excellent anti-inflammatory effect on in vitro model of psoriasis (Lin et al., 2018). To further investigate the role of kirenol, stimulation of HaCaT cells was performed with TNF-α to establish an in vitro model of psoriasis (Liu et al., 2020). In the present study, results of CCK-8 and BrdU assays demonstrated that kirenol restrained cellular phenotypes in a dose-dependent manner, which revealed that the increased cell viability of HaCaT cells induced by TNF-α was considerably reduced by kirenol treatment. Moreover, the finding of Transwell migration assay has pointed out that kirenol could suppress the migration of TNF-α stimulated HaCaT cells.
More importantly, the activated keratinocytes could generate plenty of inflammatory factors which induced cell proliferation and viability of HaCaT cells and, in turn, elevated keratinocyte proliferation and epidermal thickening to form psoriasis (Benhadou et al., 2019). It included positive feedback that excessive proliferation and epidermal thickening appeared alternately to perpetuate psoriatic lesions. NF-κB, a protein transcription factor has salient regulatory roles in immune and inflammatory pathways as well as cellular events, such as apoptosis and proliferation (Hayden & Ghosh, 2011; Patel et al., 2018; Yi et al., 2019). Furthermore, the NF-κB signaling pathway has been reported to assume an essential part in the pathogenesis of psoriasis (Wang et al., 2011). Previous study has revealed that TNF-α stimuli resulted in significant increase of inflammatory cytokines and NF-κB in keratinocytes, whereas application of genistein remarkably repressed TNF-α which induced the nuclear translocation of NF-κB and the phosphorylation of IκBα (p-IκBα), suggesting that inhibition of NF-κB expression and phosphorylation of IκBα could attenuate psoriasis (Wang et al., 2019). In addition, Liu et al. (2020) demonstrated that cimifugin exerted protective effects on psoriasis through inhibiting oxidative stress and inflammatory responses by inactivating NF-κB–MAPK pathway. Meanwhile, the fish scale collagen peptides suppressed tinflammatory cytokine expression, reactive oxygen species as well as MAPK–NF-κB pathway (Subhan et al., 2017). The results of Western blot assay demonstrated that increased concentrations of kirenol (50, 100 and 200 μg/mL) significantly down-regulated the expression of p-p65 and p-IκBα, and increased the levels of p65 and IκBα, indicating the increased levels of active phosphorylated NF-κB in psoriasis. However, the impact of kirenol on NF-κB pathway and TNF-α-stimulated HaCaT cells has not been reported. Results of the present investigation revealed that the NF-κB pathway might have been included in the impact of kirenol on TNF-α-induced HaCaT cells. TNF-α stimulation intensified the activation of the NF-κB pathway. At the same time, kirenol considerably repressed NF-κB pathway activation on TNF-α stimulated HaCaT cells.
Generally speaking, the present study discovered that kirenol activity for human keratinocytes played a protective role by inhibiting cell viability and migration as well as inflammatory response through NF-κB signaling pathway in vitro. However, some limitations associated with the study must be mentioned. For one thing, the main limitation is that the study focused more on the results of in vitro examination without in vivo and clinical analysis, which must be attained in the future investigation. For other thing, whether the effects of kirenol on NF-κB signaling pathway were direct or indirect must be identified.
Conclusion
To sum up, the study revealed for the first time the effects of specific concentrations of kirenol on the proliferation, migration and inflammation of TNF-α-stimulated HaCaT cells. Even more profound is the revelation of possible mechanisms of signaling pathways in the efficacy of kirenol. The findings of the study suggest that kirenol is conducive to improving the clinical symptoms of psoriasis, and could be a new therapeutic strategy to treat the scaly skin disease.
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Contribution of Authors
Jin Li and Fang Ren designed the study, supervised the data collection, Wenliang Yan analyzed the data, interpreted the data, Hong Sang prepare the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
Allergologia et Immunopathologia., 2020. Allergologia et immunopathologia: A new journey. Editorial. Allergologia et Immunopathologia (Madr) 48: 521–522. 10.1016/j.aller.2020.10.001
Aquino, T.M., Calvarido, M.G. and North, J.P., 2021. Interleukin 36 expression in psoriasis variants and other dermatologic diseases with psoriasis-like histopathologic features. Journal of Cutaneous Pathology. Online ahead of print. 10.1111/cup.14115
Benhadou, F., Mintoff, D. and Del Marmol, V., 2019. Psoriasis: keratinocytes or immune cells—which is the trigger? Dermatology (Basel, Switzerland) 235(2): 91–100. 10.1159/000495291
Boehncke, W.-H. and Schön, M.P., 2015. Psoriasis. Lancet (London, England) 386: 983–994. 10.1016/S0140-6736(14)61909-7
Callis Duffin, K., Gottlieb, A.B., Merola, J.F., Latella, J., Garg, A. and Armstrong, A.W., 2017. Defining outcome measures for psoriasis: the IDEOM report from the GRAPPA 2016 annual meeting. Journal of Rheumatology 44: 701–702. 10.3899/jrheum.170151
Cho, J.-W., Lee, K.-S. and Kim, C.-W., 2007. Curcumin attenuates the expression of IL-1β, IL-6, and TNF-α as well as cyclin E in TNF-α-treated HaCaT cells; NF-κB and MAPKs as potential upstream targets, International Journal of Molecular Medicine 19: 469–474. PMid: 17273796
Choi, D.H. and Hwang, H.S., 2019. Anti-inflammation activity of Brazilin in TNF-α-induced human psoriasis dermatitis skin model. Applied Biological Chemistry 62: 1–9. 10.1186/s13765-019-0455-z
Cooper, K.D., 1990. Psoriasis. Leukocytes and cytokines. Dermatologic Clinics 8(4): 737–745. PMid: 2249364
Evans, E.A., Sayers, S.R., Kodji, X., Xia, Y., Shaikh, M., Rizvi, A., et al. 2020. Psoriatic skin inflammation induces a pre-diabetic phenotype via the endocrine actions of skin secretome. Molecular Metabolism 41: 101047. 10.1016/j.molmet.2020.101047
Griffiths, C.E. and Barker, J.N., 2007. Pathogenesis and clinical features of psoriasis. Lancet (London, England) 370: 263–271. 10.1016/S0140-6736(07)61128-3
Hayden, M.S. and Ghosh, S., 2011. NF-κB in immunobiology. Cell Research 21: 223–244. 10.1038/cr.2011.13
Hoegler, K.M., John, A.M., Handler, M.Z. and Schwartz, R.A., 2018. Generalized pustular psoriasis: a review and update on treatment. Journal of the European Academy of Dermatology and Venereology (JEADV) 32: 1645–1651. 10.1111/jdv.14949
Ibrahim, S.R.M., Altyar, A.E., Sindi, I.A., El-Agamy, D.S., Abdallah, H.M., Mohamed, S.G.A. and Mohamed, G.A., 2021. Kirenol: a promising bioactive metabolite from siegesbeckia species: a detailed review. Journal of Ethnopharmacology 5(281): 114552. 10.1016/j.jep.2021.114552
Kim, M.Y., Choi, Y.W. and Hwang, H.S., 2021. Regulatory effect on skin differentiation by mevastatin in psoriasis model using TNF-α and IL-17 induced HaCaT cells. Biotechnology and Bioprocess Engineering 26: 348–358. 10.1007/s12257-020-0368-z
Kim, T.-G., Kim, D.S., Kim, H.-P. and Lee, M.-G., 2014a. The pathophysiological role of dendritic cell subsets in psoriasis. BMB Reports 47: 60–68. 10.5483/bmbrep.2014.47.2.014
Kim, M.-B., Song, Y., Kim, C. and Hwang, J.-K., 2014b. Kirenol inhibits adipogenesis through activation of the Wnt/β-catenin signaling pathway in 3T3-L1 adipocytes. Biochemical and Biophysical Research Communications 445: 433–438. 10.1016/j.bbrc.2014.02.017
Langley, R.G.B., Krueger, G.G. and Griffiths, C.E.M., 2005. Psoriasis: epidemiology, clinical features, and quality of life. Annals of the Rheumatic Diseases 64(Suppl 2): ii18–23; discussion ii24–25. 10.1136/ard.2004.033217
Lawrence, T., 2009. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harbor Perspectives in Biology 1: a001651. 10.1101/cshperspect.a001651
Lee, N., Chung, Y.C., Kang, C.I., Park, S.-M. and Hyun, C.-G., 2019. 7, 8-Dimethoxycoumarin attenuates the expression of IL-6, IL-8, and CCL2/MCP-1 in TNF-α-treated HaCaT cells by potentially targeting the NF-κB and MAPK pathways. Cosmetics 6(3): 41. 10.3390/cosmetics6030041
Lin, Z.-M., Ma, M., Li, H., Qi, Q., Liu, Y.-T., Yan, Y.-X., et al. 2018. Topical administration of reversible SAHH inhibitor ameliorates imiquimod-induced psoriasis-like skin lesions in mice via suppression of TNF-α/IFN-γ-induced inflammatory response in keratinocytes and T cell-derived IL-17. Pharmacological Research 129: 443–452. 10.1016/j.phrs.2017.11.012
Liu, W., Li, Y. and Li, C., 2020a. Kirenol exhibits the protective role against N-methyl-N-nitrosourea-induced gastric cancer in rats via modulating the oxidative stress and inflammatory markers. Journal of Environmental Pathology, Toxicology and Oncology: Official Organ of the International Society for Environmental Toxicology and Cancer 39: 345–355. 10.1615/JEnvironPatholToxicolOncol.2020035475
Liu, A., Zhao, W., Zhang, B., Tu, Y., Wang, Q. and Li, J., 2020b. Cimifugin ameliorates imiquimod-induced psoriasis by inhibiting oxidative stress and inflammation via NF-κB/MAPK pathway. Bioscience Reports 40(6): BSR20200471. 10.1042/BSR20200471
Lu, Y., Xiao, J., Wu, Z.-W., Wang, Z.-M., Hu, J., Fu, H.-Z., et al. 2012. Kirenol exerts a potent anti-arthritic effect in collagen-induced arthritis by modifying the T cells balance. Phytomedicine : International Journal of Phytotherapy and Phytopharmacology 19: 882–889. 10.1016/j.phymed.2012.04.010
Na Takuathung, M., Wongnoppavich, A., Pitchakarn, P., Panthong, A., Khonsung, P., Chiranthanut, N., Soonthornchareonnon, N. and Sireeratawong, S., 2017. Effects of wannachawee recipe with antipsoriatic activity on suppressing inflammatory cytokine production in hacat human keratinocytes. Evidence-Based Complementary and Alternative Medicine 2017: 5906539. 10.1155/2017/5906539.
Nair, R.P., Duffin, K.C., Helms, C., Ding, J., Stuart, P.E., Goldgar, D., et al. 2009. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways. Nature Genetics 41: 199–204. 10.1038/ng.311
Nickoloff, B.J., 2006. Keratinocytes regain momentum as instigators of cutaneous inflammation. Trends in Molecular Medicine 12(3): 102–106. 10.1016/j.molmed.2006.01.001
Patel, M., Horgan, P.G., McMillan, D.C. and Edwards, J., 2018. NF-κB pathways in the development and progression of colorectal cancer. Translational Research : The Journal of Laboratory and Clinical Medicine 197: 43–56. 10.1016/j.trsl.2018.02.002
Ren, J., Yang, M., Chen, J., Ma, S. and Wang, N., 2020. Anti-inflammatory and wound healing potential of kirenol in diabetic rats through the suppression of inflammatory markers and matrix metalloproteinase expressions. Biomedicine & Pharmacotherapy 129: 110475. 10.1016/j.biopha.2020.110475
Srivastava, A.K., Chand Yadav, T., Khera, H.K., Mishra, P., Raghuwanshi, N., Pruthi, V. and Prasad, R., 2021. Insights into interplay of immunopathophysiological events and molecular mechanistic cascades in psoriasis and its associated comorbidities. Journal of Autoimmunity 118: 102614. 10.1016/j.jaut.2021.102614
Subhan, F., Kang, H.Y., Lim, Y., Ikram, M., Baek, S.-Y., Jin, S., et al. 2017. Fish scale collagen peptides protect against CoCl2/TNF-α-induced cytotoxicity and inflammation via inhibition of ROS, MAPK, and NF-κB pathways in HaCaT cells. Oxidative Medicine and Cellular Longevity 2017: 9703609. 10.1155/2017/9703609.
Wang, Q., Liang, Y.-Y., Li, K.-W., Li, Y., Niu, F.-J., Zhou, S.-J., et al. 2021a. Herba siegesbeckiae: a review on its traditional uses, chemical constituents, pharmacological activities and clinical studies. Journal of Ethnopharmacology 275: 114117. 10.1016/j.jep.2021.114117
Wang, A., Wei, J., Lu, C., Chen, H., Zhong, X., Lu, Y., et al. 2019. Genistein suppresses psoriasis-related inflammation through a STAT3-NF-κB-dependent mechanism in keratinocytes. International Immunopharmacology 69: 270–278. 10.1016/j.intimp.2019.01.054
Wang, Y., Xu, J., Alarifi, S. and Wang, H., 2021b. Kirenol inhibited the cell survival and induced apoptosis in human thyroid cancer cells by altering PI3K/AKT and MAP kinase signaling pathways. Environmental Toxicology 36: 811–820. 10.1002/tox.23083
Wang, J.-P., Zhou, Y.-M. and Zhang, Y.-H., 2012. Kirenol production in hairy root culture of siegesbeckea orientalis and its antimicrobial activity. Pharmacognosy Magazine 8: 149–155. 10.4103/0973-1296.96569
Wang, Z.-M., Zhu, S.-G., Wu, Z.-W., Lu, Y., Fu, H.-Z. and Qian, R.-Q., 2011. Kirenol upregulates nuclear annexin-1 which interacts with NF-κB to attenuate synovial inflammation of collagen-induced arthritis in rats. Journal of Ethnopharmacology 137: 774–782. 10.1016/j.jep.2011.06.037
Winchell, S.A. and Watts, R.A., 1988. Relaxation therapies in the treatment of psoriasis and possible pathophysiologic mechanisms. Journal of the American Academy of Dermatology 18: 101–104.
Wolk, K., Haugen, H.S., Xu, W., Witte, E., Waggie, K., Anderson, M., et al. 2009. IL-22 and IL-20 are key mediators of the epidermal alterations in psoriasis while IL-17 and IFN-γ are not. Journal of Molecular Medicine 87: 523–536.
Wu, J., Li, Q., Jin, L., Qu, Y., Liang, B.-B., Zhu, X.-T., et al. 2019. Kirenol Inhibits the function and inflammation of fibroblast-like synoviocytes in rheumatoid arthritis in vitro and in vivo. Frontiers in Immunology 10: 1304. 10.3389/fimmu.2019.01304
Xiuhong, L., Yajun, D.U., Guoxing, L., Guomei, D., Xin, T. and Juan, X., 2019. Kirenol relieves dextran sulfate sodium-induced ulcerative colitis in mice by inhibiting inflammatory cytokines and inducing CD4 T lymphocyte apoptosis. Nan Fang Yi Ke Da Xue Xue Bao (Journal of Southern Medical University) 39: 1387–1392. 10.12122/j.issn.1673-4254.2019.12.01
Xu, F., Xu, J., Xiong, X. and Deng, Y., 2019. Salidroside inhibits MAPK, NF-κB, and STAT3 pathways in psoriasis-associated oxidative stress via SIRT1 activation. Redox Report: Communications in Free Radical Research 24: 70–74. 10.1080/13510002.2019.1658377
Yang, H. and Zheng, J., 2020. Influence of stress on the development of psoriasis. Clinical and Experimental Dermatology 45: 284–288. 10.1111/ced.14105
Yi, W., Wen, Y., Tan, F., Liu, X., Lan, H., Ye, H. and Liu, B., 2019. Impact of NF-κB pathway on the apoptosis-inflammation-autophagy crosstalk in human degenerative nucleus pulposus cells. Aging 11: 7294–7306. 10.18632/aging.102266
Zhang, Q., Lenardo, M.J. and Baltimore, D., 2017. 30 Years of NF-κB: a blossoming of relevance to human pathobiology. Cell 168: 37–57. 10.1016/j.cell.2016.12.012
Zhou, S., Li, Q., Wu, H. and Lu, Q., 2020. The pathogenic role of innate lymphoid cells in autoimmune-related and inflammatory skin diseases. Cellular & Molecular Immunology 17: 335–346. 10.1038/s41423-020-0399-6