
Download
Research article
Aloin promotes oral squamous cell carcinoma cell apoptosis and autophagy through Akt/mTOR pathway
Yongxin Hu1, Xiaosong Xiang2, Yuhui Zhang3, Zhongqi Tian4, Lijian Wang5*
1Department of Oral and Maxillofacial Surgery, Affiliated Stomatology Hospital of Guangzhou Medical University, Guangdong Engineering Research Center of Oral Restoration and Reconstruction, Guangzhou Key Laboratory of Basic and Applied Research of Oral Regenerative Medicine, Guangdong Province, China;
2Department of Orthodontic, Affiliated Stomatology Hospital of Guangzhou Medical University, Guangdong Engineering Research Center of Oral Restoration and Reconstruction, Guangzhou Key Laboratory of Basic and Applied Research of Oral Regenerative Medicine, Guangzhou, Guangdong Province, China;
3Department of Pediatric Dentistry, Affiliated Stomatology Hospital of Guangzhou Medical University, Guangdong Engineering Research Center of Oral Restoration and Reconstruction, Guangzhou Key Laboratory of Basic and Applied Research of Oral Regenerative Medicine, Guangzhou, Guangdong Province, China;
4Department of Oral and Maxillofacial Surgery, Mai Ya Oral Clinic of Shenzhen, Shenzhen, Guangdong Province, China;
5Department of Stomatology, Lishui People’s Hospital, Lishui, Zhejiang Province, China
Abstract
Aloin, a natural product, has demonstrated anticancer activity in a variety of cancers, but its role in oral squamous cell carcinoma (OSCC) has not been reported. In this study, we aimed to investigate the effects of Aloin on the biological functions of OSCC cells. Additionally, we evaluated the classical Akt/mTOR signal transduction pathway. CAL-27 OSCC cells were treated with Aloin for in vitro experiments. 5-bromo-2’-deoxyuridine (BrdU) and colony formation assays were applied to evaluate cell proliferation potential. Wound scratch and Transwell assays detected cell migration and invasion, respectively. Flow cytometry detected apoptosis. Autophagy was assessed by p62, Beclin-1, light chain 3 (LC3)-I and LC3-II protein expressions through Western blot test. Akt/mTOR signal pathway-related protein levels were also detected by Western blot test. Here, compared with the control group, Aloin treatment memorably reduced the number of BrdU-positive cells and colonies, and undermined the ability of cell migration and invasion. Moreover, Aloin promoted apoptosis and autophagy. Western blot test proved that the protein expression of p62 was down-regulated, while Beclin-1 and LC3-I/LC3-II protein expressions were up-regulated. Further investigations revealed that Aloin inhibited p-Akt and p-mTOR, suggesting that Aloin could block the activation of AKT/mTOR pathway. Our findings demonstrated that Aloin could inhibit the activation of the AKT/mTOR pathway to induce autophagic cell death, which could provide a novel vision for OSCC treatment.
Key words: oral squamous cell carcinoma, Aloin, proliferation, migration and invasion, apoptosis and autophagy, Akt/mTOR signal transduction pathway
*Corresponding Author: Lijian Wang, Department of Stomatology, Lishui People’s Hospital, No. 15 Dazhong Street, Liandu District, Lishui, Zhejiang Province 323000, China. Email: [email protected]
Received: 11 October 2021; Accepted: 29 October 2021; Published: 1 April 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Oral squamous cell carcinoma (OSCC), which primarily originates from the mucosal epithelium of the oral cavity, is the most common type of head and neck cancer, accounting for more than 90% cases (Johnson et al., 2020; Tandon, Dadhich, Saluja, Bawane, & Sachdeva, 2017). It is estimated that more than 3 million new cases of OSCC occur worldwide each year, and the number of related deaths is as high as 145,400 (Torre et al., 2015). Surgery with auxiliary radiation or chemoradiation therapy is the primary treatments for OSCC (Yang, Chen, Liu, & Yang, 2019). Owing to late diagnosis, drug resistance, adverse effects, and postoperative recurrences, the 5-year survival rate of patients with OSCC is still low (Fridman et al., 2018; Quadri, Tadakamadla, & John, 2019); hence, there is an urgent need to find a suitable treatment.
Natural products (such as traditional Chinese medicine) are established to be an essential source of medicines against cancer and infectious diseases (Sanchez, Gonzalez-Burgos, Iglesias, & Gomez-Serranillos, 2020). Complementary and alternative medicines, such as natural products, have gained more and more attention for treatment of malignant tumors (Kitamura et al., 2020). Aloin is a natural bioactive anthracycline extracted from the leaves of the perennial herb Aloe vera (Zhong et al., 2015). Aloin has a wide range of medicinal values, such as neuroprotection (Chang et al., 2016), cardiopulmonary protection (Birari et al., 2020; Lei et al., 2020; McMullen & McMullen, 1989), regulation of immunity (W. Lee et al., 2019) and the regulation of microbial groups (Boudreau et al., 2017; I. C. Lee & Bae, 2021). In addition, Aloin has been reported to have anticancer effects in many types of cancers. Wang et al. (2020b) found that Alion could regulate the proliferation and migration of gastric cancer cells through NOX2-reactive oxygen species (ROS)-mediated Akt/mTOR, the signal transducer and activator of transcription-3 (Stat3), and Nuclear Factor kappa B (NF-κB) signals. Sun et al. (2020) reported that Aloin reduced the phosphorylation levels of PI3K, AKT and mTOR in liver cancer cells, and promoted liver cancer cell apoptosis and autophagy to enhance the anticancer effect of metformin. However, the role of Aloin in OSCC is still unknown.
Based on the multiple effects of Aloin, we speculated that it also exerts anticancer activity in OSCC. Clarifying the mechanisms related to the development and progression of OSCC would help to improve the level of treatment. In this study, we aimed to explore the effects of Aloin on the functional level of OSCC cells CAL-27, including cell proliferation, cell migration and invasion, cell apoptosis and autophagy. In addition, we also evaluated the classical Akt/mTOR signal transduction pathway, the primary pathway for autophagy regulation (Kim & Guan, 2015). These findings of Aloin might provide a potential strategy for further prognosis for OSCC treatment.
Methods
OSCC cell line
The study adopted CAL-27 cells (Catalog No. CRL-2095; American Type Culture Collection [ATCC], USA) as the research object in vitro. This cell line is epithelial, polygonal with a highly granular cytoplasm. CAL-27 cells were kept in Dulbecco’s Modified Eagle’s Medium (Catalog No. 30-2002, ATCC, USA) supplemented with 10% fetal bovine serum (FBS) (Catalog No. 30-2021; ATCC) in a humid environment at 37°C.
Chemicals
Natural antitumor anthraquinone glycosides Aloin, having purity of 98.32%, was purchased from MedChem Express, USA (Catalog No. No. 123). It was dissolved in phosphate buffered saline (PBS) (Catalog No. D917703; MACKLIN, Shanghai, China) to prepare a 100-mg/mL storage solution.
Aloin treatment
CAL-27 cells were treated with Aloin for 24 h. The cells were divided into four groups according to the treatment dose, namely control group (equal amount of PBS), 100-μg/mL group, 200-μg/mL group, and 400-μg/mL group. Subsequently, the cells were collected to evaluate respective functions (Z. Wang et al., 2020).
5-bromo-2’-deoxyuridine (BrdU) assay
CAL-27 cells were resuspended and around 1 × 106 cells were seeded onto 35-mm petri dishes with built-in glass slides (Catalog No. 150460; Thermo Fisher Scientific, USA) and fixed with 4% paraformaldehyde (Catalog No. P885233; MACKLIN) for 15 min. Then cells were incubated in DNA labeling solution (TUNEL Assay Kit, Catalog No. ab66110; Abcam, UK) for 60 min at 37°C. Cells were incubated with anti-BrdU-Red antibody in dark at room temperature for 30 min. DAPI (4’,6-Diamidino-2-phenylindole [double stranded DNA staining], Catalog No. C1002; Beyotime, Shanghai, China) was utilized as a counterstain (blue) for nucleus. Cells were analyzed within 3 h of staining and the five fields of view were randomly selected to acquire pictures by fluorescence microscopy (Leica, TCS SP5II, Germany).
Colony formation assay
Approximately 500 cells were evenly inoculated onto 60-mm petri dishes (Catalog No. 353002; Corning, USA) and cultured for 14 days; the medium was replenished with fresh medium every 24 h. Subsequently, cells were fixed with 4% paraformaldehyde for 30 min and stained with 0.2% crystal violet staining solution (Catalog No. V5265; Sigma-Aldrich, Germany) for 15 min. The images were acquired and the number of colonies formed (≥50 cells) were counted using GX53 microscope (Olympus Corporation, Japan).
Wound scratch assay
Wound scratch assay was performed to evaluate cell migration. CAL-27 cells were inoculated onto 6-well plates, https://www.sigmaaldrich.cn/CN/zh/product/sigma/cls3516?context=product, (Catalog No. CLS3516; Corning) and cultured to 90–100% monolayer confluency; a sterile pipette was used to make a gap of about 1 mm. Subsequently, the cells were treated with different concentrations of Aloin for 24 h. The images were captured by GX53 microscope.
Cell invasion assay
Transwell assay was performed to evaluate cell invasion. Approximately 5 × 104 CAL-27 cells were inoculated onto upper transwell chambers (Catalog No. 356234; BD Bioscience, USA) with 8-µm pore size coated with Matrigel. The upper chambers were added with serum-free medium and the lower chambers were filled with medium supplemented with 10% FBS. In addition, PBS or Aloin with different concentrations was added into the upper chamber medium. After incubation for 24 h, cells in the lower chambers were fixed for 15 min and stained for 30 min with 0.1% crystal violet staining solution. The invasive number of cells was calculated and imaged by GX53 microscope.
Apoptosis
Approximately 1 × 105 cells were resuspended with binding solution using Annexin V-FITC/7-AAD Fluorescence Double Stain Cell Apoptosis Detection Kit according to manufacturers’ instructions (Procell Life Science & Technology Co. Ltd., Hubei, China). Then these were added with 5-μL annexin V-FITC and 5-μL 7-ADD staining solution followed by incubation at room temperature for 15 min. The apoptosis of cells was analyzed on a flow cytometer (CytoFLEX S Flow Cytometer; Beckman Coulter, Indianapolis, USA).
Western Blot test
Western blot test was performed to detect the protein levels of p62, Beclin-1, LC3-I, LC3-II, AKT, phospho (p)-AKT, mTOR, and p-mTOR. Briefly, the experiment was divided into following three parts: total protein extraction, membrane transfer, and antibody incubation.
CAL-27 cells were lysed with RIPA lysis buffer (Catalog No. 20-188; Sigma-Aldrich). Then the concentrations of protein were quantified by a bicinchoninic acid (BCA) kit (Catalog No. P0011; Beyotime). Equal amount of total proteins in each sample was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride (PVDF) Western blotting membranes (Catalog No. FFP28; Beyotime). After being blocked with 5% skimmed milk at room temperature for 1 h, the membranes were incubated with primary antibodies comprised those against p62 (rabbit, 1:1,000, #23214; Cell Signaling Technology [CST], USA), against Beclin-1 (rabbit, 1:2,000, ab207612; Abcam, UK), against LC3-I/LC3-II (rabbit, 1:1,000, #12741; CST), against AKT (rabbit, 1:1,000, ab8933; Abcam, UK), against p-AKT (rabbit, 1:1,000, ab38449; Abcam, USA), against mTOR (rabbit, 1:1,000, #2983; CST, USA), and against p-mTOR (rabbit, 1:1,000, #5536; CST). GAPDH (Glyceraldehyde 3-phosphate dehydrogenase) (mouse, 1:1,000, ab8245; Abcam, UK) was used as an internal reference. Afterwards, the membranes were incubated with the goat anti-rabbit immunoglobulin G (IgG; 1:3000, ab6721; Abcam, USA) or goat anti-mouse IgG (1:3000, ab6789; Abcam, USA) secondary antibodies. The signals of proteins were tested and collected by enhanced chemiluminescence (ECL) kit (Catalog No. P0018S; Beyotime). The autophagy level was analyzed by p62, Beclin-1, LC3-I, and LC3-II, and Akt/mTOR signal transduction pathway was assessed by the phosphorylation levels of Akt and mTOR.
Statistical methods
All experiments were performed for at least three times, and all values were presented as mean + standard deviation (SD). The GraphPad Prism 8 software (GraphPad, CA, USA) was utilized for data analyzing. One-way analysis of variance (ANOVA) was used with one categorical independent variable in mutigroups with paired t-test to analyze data from two groups. For measurements, P < 0.05 was considered significant.
Results
Aloin inhibited the proliferation of OSCC cells
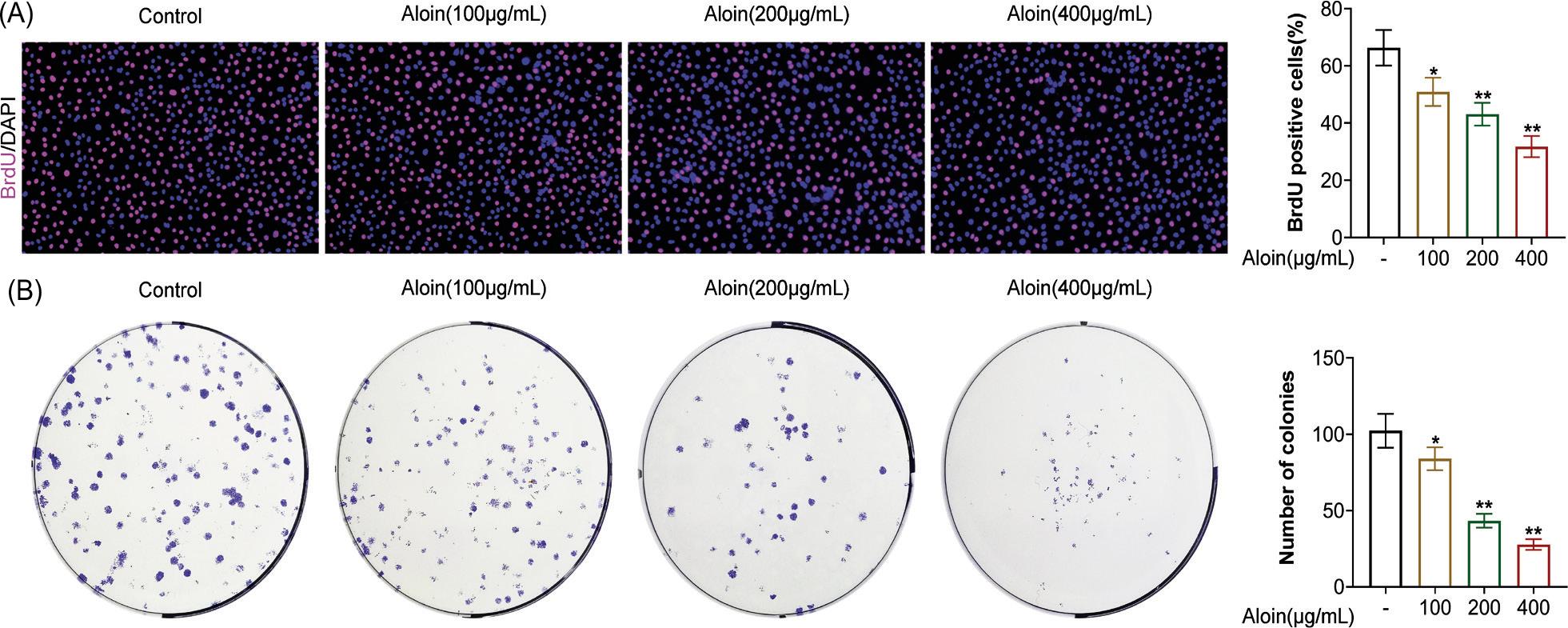
First, we evaluated the effect of Aloin on the proliferation of CAL-27 cells. BrdU and colony formation assays are recognized methods for judging cell proliferation. As shown in Figure 1A (P < 0.05), the merger graph of BrdU and DAPI illustrated that Aloin could reduce the number of BrdU-positive cells (red) in a dose-dependent manner, which was consistent with statistical results. Moreover, as shown in Figure 1B (P < 0.05), Aloin also significantly decreased the number of single colonies in a dose-dependent manner. The evidence indicated that Aloin could effectively inhibit the proliferation of OSCC cancer cells.
Figure 1. Aloin inhibited the proliferation of CAL-27 oral squamous cell carcinoma (OSCC) cells. CAL-27 cells were treated with Aloin (0, 100, 200 or 400-μg/mL group) for 24 h. (A) BrdU labels newly synthesized DNA in living cells (Red) to assess proliferation. (B) 14-day colony formation experiment were performed, and were captured images and counted the number of colonies formed (≥50 cells) to assess proliferation. *P < 0.05, **P < 0.01 vs. -.
Aloin inhibited the migration and invasion of OSCC cells
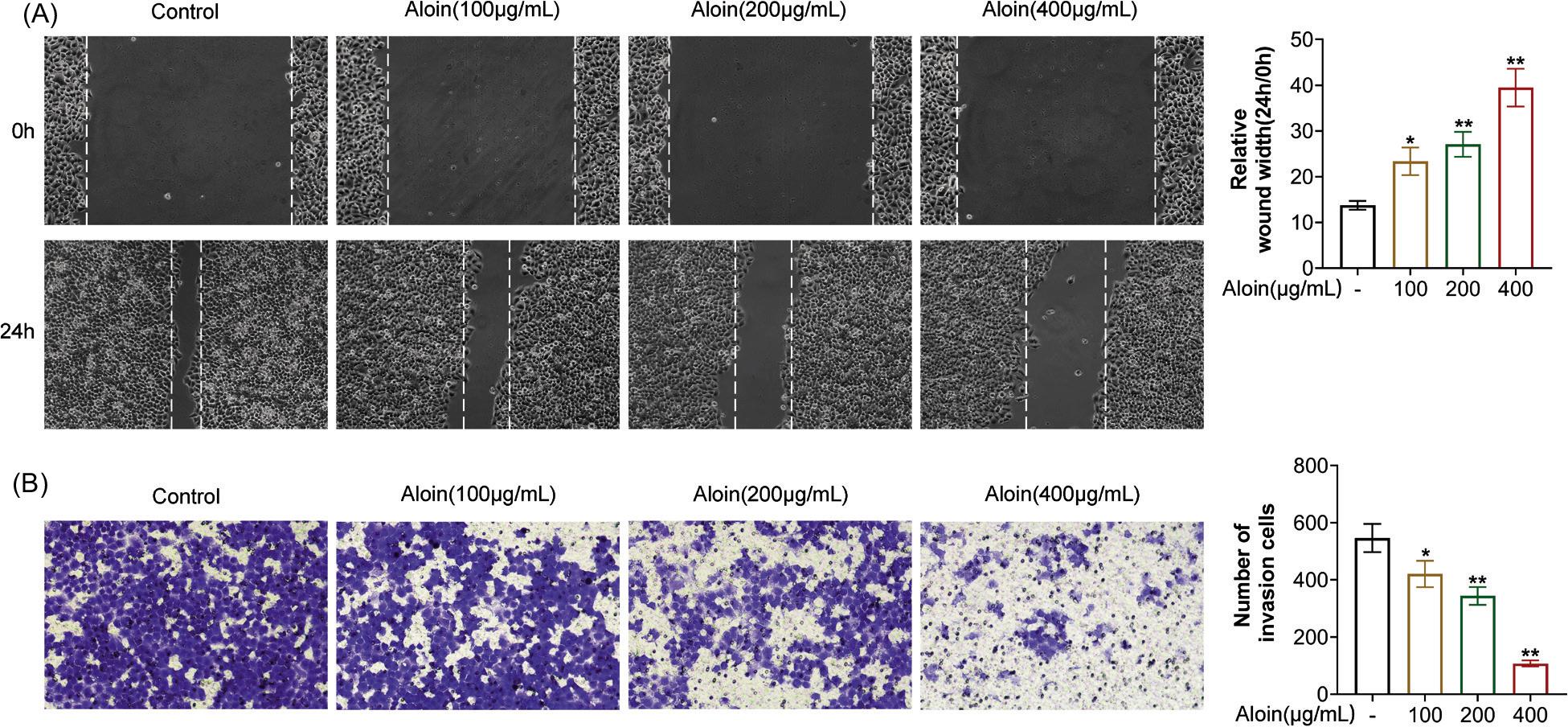
Migration and invasion are the driving force for the spread of cancer cells (Zhang, Tang, Chen, Du, & Qu, 2020). The results of the Wound Scratch assay demonstrated that the width of wound 24 h after Aloin treatment was greater than that of the control group, suggesting that Aloin weakened the migration ability of CAL-27 cells (Figure 2A, P < 0.05). Transwell assay results evidenced that there were markedly fewer invasive cells in Aloin-treated group (Figure 2B, P < 0.05), suggesting that Aloin inhibits the invasive ability of CAL-27 cells. Similarly, the inhibitory effect of high concentrations of Aloin was more obvious (Figures 2A and 2B). These results suggested that Aloin could attenuate the migration and invasion of OSCC cells.
Figure 2. Aloin inhibited the migration and invasion of CAL-27 cells. (A) Wound scratch assay was performed to evaluate cell migration. The cells were treated with different concentrations of Aloin for 24 h. (B) Transwell assay was performed to evaluate cell invasion. P < 0.05, P < 0.01 vs. -.
Aloin promoted the apoptosis of OSCC cells
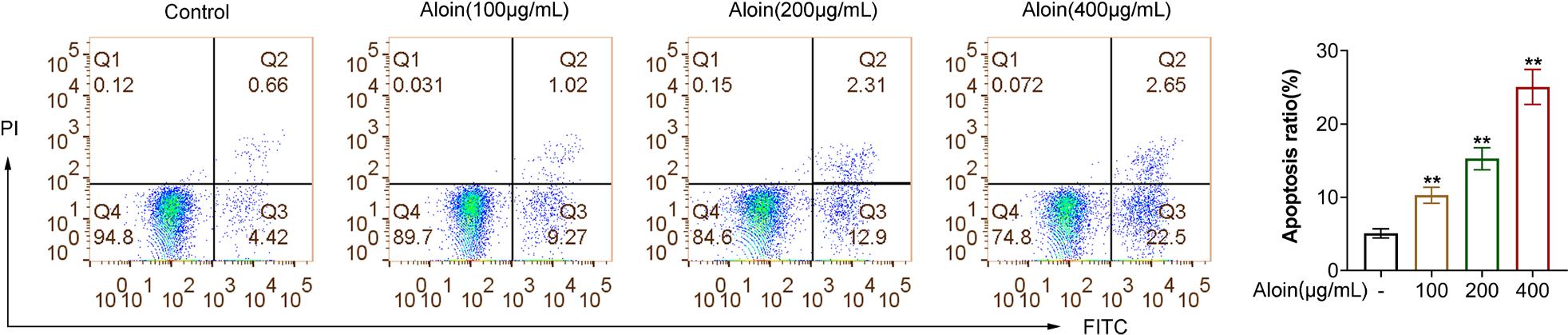
In addition to inhibiting proliferation, promoting apoptosis of cancer cells is also an important strategy for cancer treatment. We utilized flow cytometry to detect the rate of cell apoptosis, and the results demonstrated that Aloin promoted cell apoptosis in a dose-dependent manner (Figure 3, P < 0.01).
Figure 3. Aloin promoted the apoptosis of CAL-27 cells. Flow cytometry was performed to assess apoptosis. **P < 0.01 vs. -.
Aloin promoted autophagy in oral squamous cancer cells
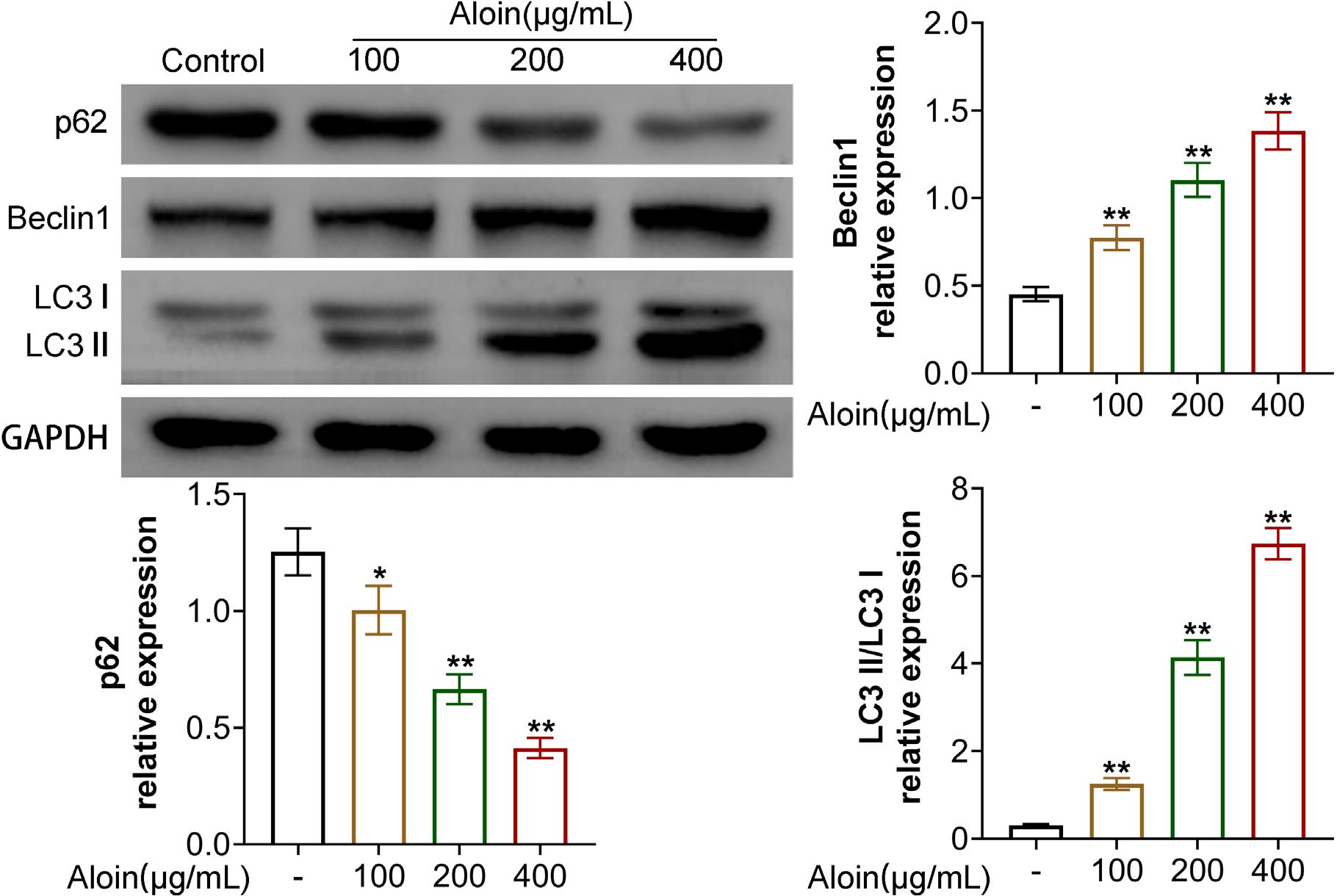
Levels of p62, Beclin-1, LC3-I and LC3-II proteins are closely related to autophagy (Wei, Wu, Liu, & Xie, 2020; Xu & Qin, 2019). Compared to the control group, the expressions of p62 and Beclin-1, and LC3-I/LC3-II have all changed (Figure 4, P < 0.01). The expression of p62 was down-regulated by Aloin, while Beclin-1 and LC3-I/LC3-II were up-regulated after Aloin treatment (Figure 4, P < 0.01). The evidence indicated that Aloin could induce autophagy in OSCC.
Figure 4. Aloin promoted autophagy of CAL-27 cells. Western blot test was performed to detect the protein expressions of p62, Beclin-1, light chain 3 (LC3)-I and LC3-II to assess autophagy. GAPDH was used as an internal reference. **P < 0.01 vs. -.
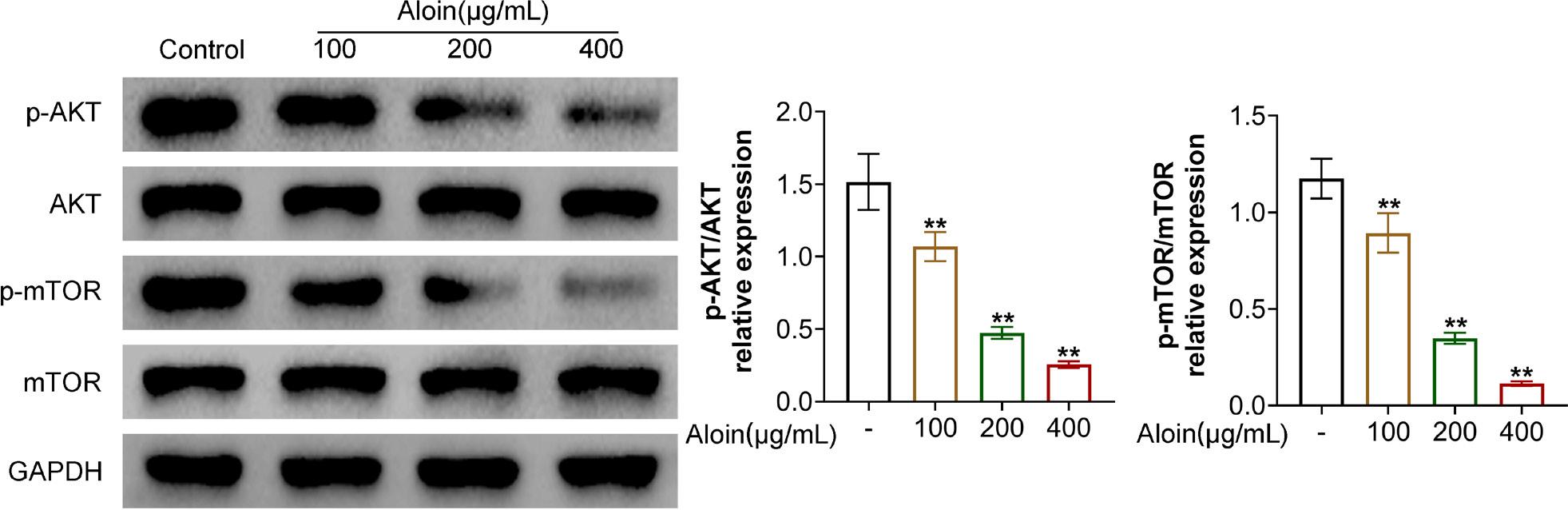
Aloin blocked the Akt/mTOR pathway
The Akt/mTOR pathway is the hub of a variety of signals, and is also an important pathway related to autophagy. We detected the phosphorylation levels of Akt and mTOR pathways. As expected, the values of p-Akt/Akt and p-mTOR/mTOR were decreased (Figure 5, P < 0.01), suggesting that the Akt/mTOR pathway might be one of the pathways in which Aloin inhibited OSCC cell functioning.
Figure 5. Aloin inhibited the Akt/mTOR pathway. Western blot test was performed to detect the protein expressions of AKT, phosphorylated (p)-AKT, mTOR, and p-mTOR. GAPDH was used as an internal reference. **P < 0.01 vs. -.
Discussion
In recent years, the importance of drug therapy for malignant tumors has increased with passing years (J. Wang et al., 2020; W. E. Yang et al., 2019). Previous studies have demonstrated that natural plant products could be used as chemopreventive or therapeutic agents for human cancer cells (Khan et al., 2019). As per our knowledge, our study for the first time has investigated the role of Aloin in OSCC. The in vitro data have initially revealed that Aloin plays a tumor-suppressor activity in OSCC. It inhibits tumor proliferation, migration, and invasion potential, and stimulates cancer cells’ apoptosis and autophagy.
A number of studies have indicated that the disorder of cell proliferation, apoptosis and metastasis is a major inducement for the occurrence and development of cancer, especially tumor metastasis, which is one of the primary problems faced during the clinical treatment of various cancers (Irani, 2016). Lymph node metastasis of OSCC in the neck reduces patient’s survival rate by approximately 50% (Sharma, Kim, & Paeng, 2018). Targeted autophagy has been emphasized as a feasible treatment strategy with clinical utility in cancer treatment (Khan et al., 2020). In this study, based on the study of Wang et al. (Z. Wang et al., 2020), we selected the concentration range of 100–400 μg/mL Aloin and evaluated the effects of the gradient Aloin on cell proliferation, migration, invasion and apoptosis in CAL-27 cells. Encouragingly, Aloin treatments significantly inhibited the proliferation, migration and invasion potential of cancer cells, and stimulated cancer cells to undergo apoptosis and autophagy. Consistently, the higher the concentration of Aloin, the more pronounced the anticancer effect. The evidence suggested that Aloin is expected to become a potential molecule for the treatment of OSCC. It was reported that high-mobility group protein B1 (HMGB1) could be used as a target for Aloin (Li et al., 2020; Tao et al., 2019). Nevertheless, Aloin still lacks a lot of mechanism research.
Autophagy is a lysosome-dependent cellular degradation process involving the degradation of intracellular components to maintain cell (White, 2012). We utilized Western blot test to detect p62, Beclin-1, LC3-I and LC3-II levels to assess autophagy in CAL-27 cells. The multidomain protein p62, also known as sequestosome 1, occupies a central position in autophagy and apoptosis, and is an autophagy receptor and a selective substrate for autophagy (Ichimura, Kominami, Tanaka, & Komatsu, 2008; Islam, Sooro, & Zhang, 2018). In OSCC, high cytoplasmic p62 expression could be a poor prognostic marker for OSCC patients (Liang et al., 2018). Beclin-1, LC3-I and LC3-II are the hallmark proteins of autophagy (Wei et al., 2020; Xu & Qin, 2019). It has been reported that activated Beclin-1 can induce autophagy (Kinarivala, Patel, Boustany, Al-Ahmad, & Trippier, 2017). We found that Aloin treatment caused the up-regulation of Beclin-1 and LC3 I/LC3-II proteins, while it downregulated P62 protein. Therefore, we suggest that Aloin induces autophagy in OSCC.
Unlike apoptosis, autophagy is a mechanism by which cells try to survive. In some cases, autophagy constitutes a stress adaptation that avoids cell death (M Chiara Maiuri, Einat Zalckvar, Adi Kimchi, & Guido Kroemer, 2007). Interestingly, accumulated studies indicate that there is a complex relationship between autophagy and apoptosis (D’Arcy, 2019; Maiuri, Zalckvar, Kimchi, & Kroemer, 2007). They play seemingly opposite biological roles in response to genotoxic or pharmacological stress but share common regulatory elements, including Akt/mTOR signal pathway (Lin et al., 2019). The classic Akt/mTOR signal transduction pathway is the most common dysregulated cell pathway in human cancers, regulating the development of many cancers, including proliferation, apoptosis, chemoresistance and autophagy (Xia & Xu, 2015). Akt/mTOR pathway negatively regulates the process of autophagy. Western blot test results demonstrated that Aloin treatment significantly reduced the levels of p-AKT and p-mTOR in CAL-27 cells as described previously. Autophagy and apoptosis may regulate each other after treatment to achieve anticancer effects (Adhauliya, Kalappanavar, Ali, & Annigeri, 2016). This finding confirmed that Aloin could inhibit the activation of AKT/mTOR pathway to induce autophagic cell death.
In conclusion, this study strongly demonstrated that Aloin, a natural plant ingredient, is a potential antitumor molecule , which can induce OSCC cell protective autophagy to exert anticancer effects. At present, the study of Aloin is still in its infancy. Further studies toward Aloin in OSCC are required to validate the performances and properties in vivo and in real practice.
Competing Interests
The authors state that there are no conflicts of interest to disclose.
Contribution of Authors
Yongxin Hu and Xiaosong Xiang designed the study and supervised data collection. Yuhui Zhang and Zhongqi Tian analyzed and interpreted the data. Lijian Wang prepared the manuscript for publication and reviewed its draft. All authors read and approved the final manuscript.
Funding
This work was supported by Special Clinical Technology of Guangzhou. (Grant No. 2019 TS41).
REFERENCES
Adhauliya, N., Kalappanavar, A.N., Ali, I.M. and Annigeri, R.G., 2016. Autophagy: a boon or bane in oral cancer. Oral Oncology 61: 120–126. 10.1016/j.oraloncology.2016.09.001
Birari, L.A., Mahajan, U.B., Patil, K.R., Patil, D.D., Bagul, N.A., Belemkar, S., et al. 2020. Aloin protects against arsenic trioxide-induced myocardial membrane damage and release of inflammatory cytokines. Naunyn Schmiedebergs Arch Pharmacology 393: 1365–1372. 10.1007/s00210-020-01833-1
Boudreau, M.D., Olson, G.R., Tryndyak, V.P., Bryant, M.S., Felton, R.P. and Beland, F.A., 2017. From the cover: aloin, a component of the aloe vera plant leaf, induces pathological changes and modulates the composition of microbiota in the large intestines of F344/N male rats. Toxicological Sciences 158: 302–318. 10.1093/toxsci/kfx105
Chang, R., Zhou, R., Qi, X., Wang, J., Wu, F., Yang, W., et al. 2016. Protective effects of aloin on oxygen and glucose deprivation-induced injury in PC12 cells. Brain Research Bulletin (BRB) 121: 75–83. 10.1016/j.brainresbull.2016.01.001
D’Arcy, M.S., 2019. Cell death: a review of the major forms of apoptosis, necrosis and autophagy. Cell Biology International 43: 582–592. 10.1002/cbin.11137
Fridman, E., Na’ara, S., Agarwal, J., Amit, M., Bachar, G., Villaret A.B., et al. 2018. The role of adjuvant treatment in early-stage oral cavity squamous cell carcinoma: an international collaborative study. Cancer 124: 2948–2955. 10.1002/cncr.31531
Ichimura, Y., Kominami, E., Tanaka, K. and Komatsu, M., 2008. Selective turnover of p62/A170/SQSTM1 by autophagy. Autophagy 4: 1063–1066. 10.4161/auto.6826
Irani, S., 2016. Distant metastasis from oral cancer: a review and molecular biologic aspects. Journal of International Society of Preventive and Community Dentistry 6: 265–271. 10.4103/2231-0762.186805
Islam, M.A., Sooro, M.A. and Zhang, P., 2018. Autophagic regulation of p62 is critical for cancer therapy. International Journal of Molecular Sciences 19: 1405. 10.3390/ijms19051405
Johnson, D.E., Burtness, B., Leemans, C.R., Lui, V.W.Y., Bauman, J.E. and Grandis, J.R., 2020. Head and neck squamous cell carcinoma. Nature Reviews Disease Primers 6: 92. 10.1038/s41572-020-00224-3
Khan, T., Ali, M., Khan, A., Nisar, P., Jan, S.A., Afridi, S., et al. 2019. Anticancer plants: a review of the active phytochemicals, applications in animal models, and regulatory aspects. Biomolecules 10(1). 10.3390/biom10010047
Khan, T., Relitti, N., Brindisi, M., Magnano, S., Zisterer, D., Gemma, S., et al. 2020. Autophagy modulators for the treatment of oral and esophageal squamous cell carcinomas. Medicinal Research Reviews 40: 1002–1060. 10.1002/med.21646
Kim, Y.C. and Guan, K.L., 2015. mTOR: a pharmacologic target for autophagy regulation. Journal of Clinical Investigation 125: 25–32. 10.1172/JCI73939
Kinarivala, N., Patel, R., Boustany, R.M., Al-Ahmad, A. and Trippier, P.C., 2017. Discovery of aromatic carbamates that confer neuroprotective activity by enhancing autophagy and inducing the anti-apoptotic protein B-cell lymphoma 2 (Bcl-2). Journal of Medicinal Chemistry 60: 9739–9756. 10.1021/acs.jmedchem.7b01199
Kitamura, N., Sento, S., Yoshizawa, Y., Sasabe, E., Kudo, Y. and Yamamoto, T., 2020. Current trends and future prospects of molecular targeted therapy in head and neck squamous cell carcinoma. International Journal of Molecular Sciences 22: 240. 10.3390/ijms22010240
Lee, I.C. and Bae, J.S., 2021. Inhibitory effects of aloin on TGFBIp-mediated septic responses. Journal of Asian Natural Products Research 23: 189–203. 10.1080/10286020.2019.1711066
Lee, W., Yang, S., Lee, C., Park, E.K., Kim, K.M., Ku, S.K., et al. 2019. Aloin reduces inflammatory gene iNOS via inhibition activity and p-STAT-1 and NF-kappaB. Food and Chemical Toxicology 126: 67–71. 10.1016/j.fct.2019.02.025
Lei, J., Shen, Y., Xv, G., Di, Z., Li, Y. and Li, G., 2020. Aloin suppresses lipopolysaccharide-induced acute lung injury by inhibiting NLRP3/NF-kappaB via activation of SIRT1 in mice. Immunopharmacology and Immunotoxicology 42: 306–313. 10.1080/08923973.2020.1765373
Li, P., Ren, K., yin Liang, Y., kai Liu J., wen Liang, Z., et al. 2020. Aloin promotes cell apoptosis by targeting HMGB1-TLR4-ERK axis in human melanoma cells. EXCLI Journal 19: 641.
Liang, L., Luo, H., He, Q., You, Y., Fan, Y. and Liang, J., 2018. Investigation of cancer-associated fibroblasts and p62 expression in oral cancer before and after chemotherapy. Journal of Cranio-Maxillofacial Surgery 46: 605–610. 10.1016/j.jcms.2017.12.016
Lin, C.W., Chin, H.K., Lee, S.L., Chiu, C.F., Chung, J.G., Lin, Z.Y., et al. 2019. Ursolic acid induces apoptosis and autophagy in oral cancer cells. Environmental Toxicology 34: 983–991. 10.1002/tox.22769
Maiuri, M.C., Zalckvar, E., Kimchi, A. and Kroemer, G., 2007. Self-eating and self-killing: crosstalk between autophagy and apoptosis. Nature Reviews in Molecular Cell Biology 8: 741–752. 10.1038/nrm2239
McMullen, P. and McMullen, A., 1989. Ambulatory centers can be a source of liability, too. Nursing Connections 2(3): 42–44.
Quadri, M.F.A., Tadakamadla, S.K. and John, T., 2019. Smokeless tobacco and oral cancer in the Middle East and North Africa: a systematic review and meta-analysis. Tobacco Induced Diseases 17: 56. 10.18332/tid/110259
Sanchez, M., Gonzalez-Burgos, E., Iglesias, I. and Gomez-Serranillos, M.P., 2020. Pharmacological update properties of aloe vera and its major active constituents. Molecules 25: 1324. 10.3390/molecules25061324
Sharma, A., Kim, J.W. and Paeng, J.Y., 2018. Clinical analysis of neck node metastasis in oral cavity cancer. Journal of the Korean Association of Oral and Maxillofacial Surgeons 44: 282–288. 10.5125/jkaoms.2018.44.6.282
Sun, R., Zhai, R., Ma, C. and Miao, W., 2020. Combination of aloin and metformin enhances the antitumor effect by inhibiting the growth and invasion and inducing apoptosis and autophagy in hepatocellular carcinoma through PI3K/AKT/mTOR pathway. Cancer Medicine 9: 1141–1151. 10.1002/cam4.2723
Tandon, P., Dadhich, A., Saluja, H., Bawane, S. and Sachdeva, S., 2017. The prevalence of squamous cell carcinoma in different sites of oral cavity at our rural health care centre in Loni, Maharashtra—a retrospective 10-year study. Contemporary Oncology (Poznan, Poland) 21: 178–183. 10.5114/wo.2017.68628
Tao, H., Tang, T., Wang, S., Wang, Z., Ma, Y., Cai, T., et al. 2019. The molecular mechanisms of aloin induce gastric cancer cells apoptosis by targeting high mobility group box 1. Drug Design, Development and Therapy 13: 1221. 10.2147/DDDT.S201818
Torre, L.A., Bray, F., Siegel, R.L., Ferlay, J., Lortet-Tieulent, J. and Jemal, A., 2015. Global cancer statistics, 2012. Cancer Journal for Clinicians 65: 87–108. 10.3322/caac.21262
Wang, Z., Tang, T., Wang, S., Cai, T., Tao, H., Zhang, Q., et al. 2020b. Aloin inhibits the proliferation and migration of gastric cancer cells by regulating NOX2-ROS-mediated pro-survival signal pathways. Drug Design, Development and Therapy 14: 145–155. 10.2147/DDDT.S219247
Wang, J., Wang, S., Wang, Y., Wang, L., Xia, Q., Tian, et al. 2020a. Chemopreventive effect of modified zengshengping on oral cancer in a hamster model and assessment of its effect on liver. Journal of Ethnopharmacology 255: 112774. 10.1016/j.jep.2020.112774
Wei, M., Wu, Y., Liu, H. and Xie, C., 2020. Genipin induces autophagy and suppresses cell growth of oral squamous cell carcinoma via PI3K/AKT/MTOR pathway. Drug Design, Development and Therapy 14: 395–405. 10.2147/DDDT.S222694
White, E., 2012. Deconvoluting the context-dependent role for autophagy in cancer. Nature Reviews Cancer 12: 401–410. 10.1038/nrc3262
Xia, P. and Xu, X.Y., 2015. PI3K/Akt/mTOR signaling pathway in cancer stem cells: from basic research to clinical application. American Journal of Cancer Research 5(5): 1602–1609.
Xu, H.D. and Qin, Z.H., 2019. Beclin 1, Bcl-2 and autophagy. Advances in Experimental Medicine and Biology 1206: 109–126. 10.1007/978-981-15-0602-4_5
Yang, Y., Chen, D., Liu, H. and Yang, K., 2019b. Increased expression of lncRNA CASC9 promotes tumor progression by suppressing autophagy-mediated cell apoptosis via the AKT/mTOR pathway in oral squamous cell carcinoma. Cell Death and Disease 10: 41. 10.1038/s41419-018-1280-8
Yang, W.E., Ho, Y.C., Tang, C.M., Hsieh, Y.S., Chen, P.N., Lai, C.T., et al. 2019a. Duchesnea indica extract attenuates oral cancer cells metastatic potential through the inhibition of the matrix metalloproteinase-2 activity by down-regulating the MEK/ERK pathway. Phytomedicine 63: 152960. 10.1016/j.phymed.2019.152960
Yap, K.M., Sekar, M., Seow, L.J., Gan, S.H., Bonam, S.R., Rani, N.N.I.M., et al. 2021. Mangifera indica (Mango): a promising medicinal plant for breast cancer therapy and understanding its potential mechanisms of action. Breast Cancer: Targets and Therapy 13(13): 471–503. 10.2147/BCTT.S316667
Zhang, Y., Tang, K., Chen, L., Du, M. and Qu, Z., 2020. Exosomal CircGDI2 suppresses oral squamous cell carcinoma progression through the regulation of MiR-424-5p/SCAI axis.Cancer Management and Research 12: 7501–7514. 10.2147/CMAR.S255687
Zhong, J.S., Huang, Y.Y., Zhang, T.H., Liu, Y.P., Ding, W.J., Wu, X.F., Xie, Z.Y., Luo, H.B. and Wan, J.Z., 2015. Natural phosphodiesterase-4 inhibitors from the leaf skin of Aloe barbadensis Miller. Fitoterapia 100: 68–74. 10.1016/j.fitote.2014.11.018