
Download
% Fat yield = Weight of extracted oil g Weight of dried sample g × 100 AV = Vol. of KOH × Normality of KOH Weight of sample g × 56.1 Saponification value SV = 56.1 B-S × Normality or HCI Weight of sample g IV = B-S × Normality of Na 2 S 2 O 3 × 0.127 Weight of sample g × 100 PV = S-B × Normality of Na 2 S 2 O 3 × 1 , 000 Weight of sample g SG = Density of fat Kg/m 3 Density of water Kg/m 3 η 1 = η 1 × ρ 1 × t 1 ρ 2 × t 2 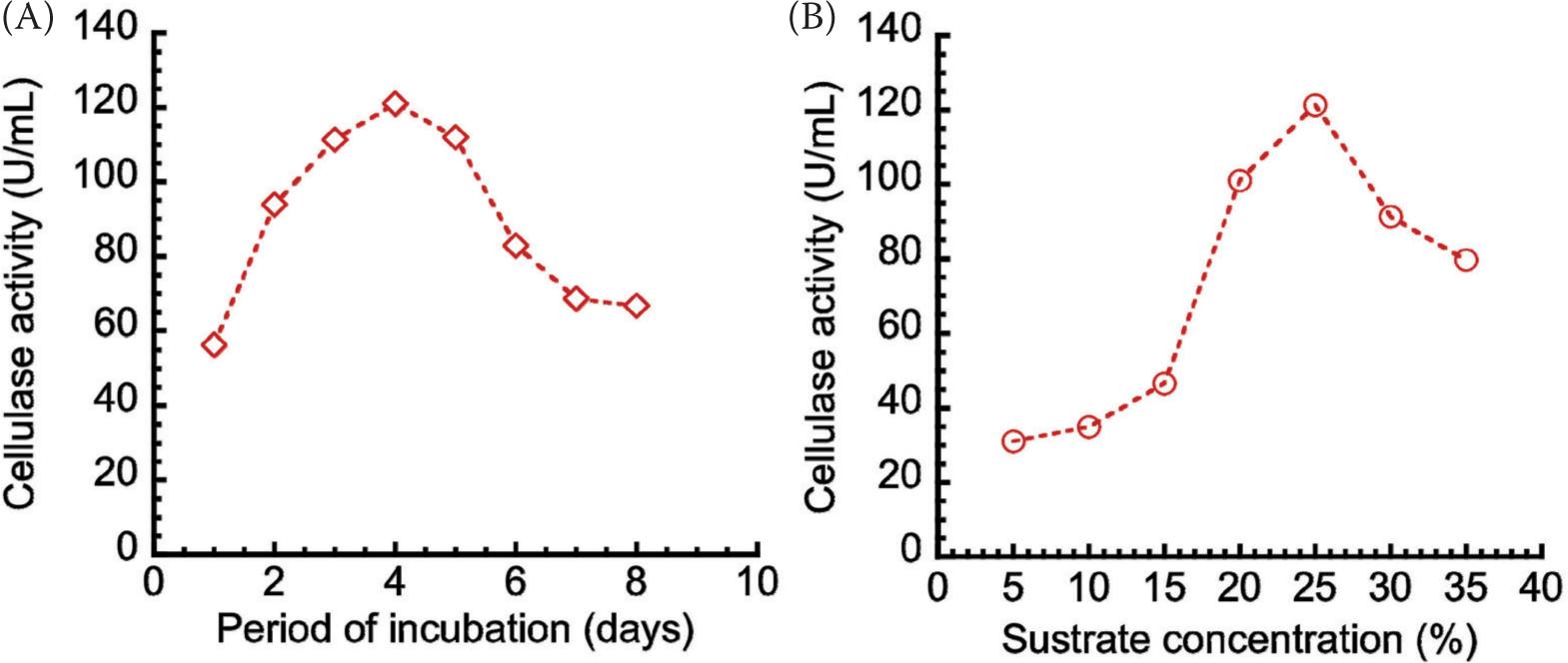
ORIGINAL ARTICLE
An enhanced fat extraction from Pycnanthus angolensis (African nutmeg) seeds using cellulase from Aspergillus niger strain BC23
Marius Obiwanne Chiwetalu1, Emmanuel Chekwube Ossai1*, Osmond Chukwuma Enechi1, Christian Nnabuchi Ugwu1, Vincent E.O. Ozogwu1, Charles Odilichukwu R. Okpala2*, Obioma Uzoma Njoku1
1Department of Biochemistry, Faculty of Biological Sciences, University of Nigeria, Nsukka, Enugu State, Nigeria;
2Department of Functional Food Products Development, Wrocław University of Environmental and Life Sciences, Wrocław, Poland
Abstract
A promising extraction method for obtaining fat from Pycnanthus angolensis seeds through Aspergillus niger strain BC23 enzyme pretreatment and particle size reduction is presented in this work. The employed A. niger enzyme was produced from corn bagasse waste material, which served as the only source of carbon. P. angolensis seeds were dried, pulverized and subsequently sieved into two different particle sizes of 1.0 mm and 1.4 mm. This was followed by enzyme pretreatment prior to fat extraction. Results demonstrated that while the peak enzyme activity (121.036 U/mL) occurred on day 4 of fermentation, the maximum cellulase activity could be achieved at 60% saturation of ammonium sulfate purification. Compared with the results of water-treated and untreated seed samples, the fat yield, free fatty acid and iodine values of the enzyme-treated seed samples were considerably higher (P < 0.05). As peroxide value was less in the enzyme-assisted extracted fat, which pointed to a good-quality oil, the decreased refractive index suggested peak unsaturation. Despite considerable differences (P < 0.05) in refractive indices, fat’s specific gravity matched (P > 0.05) when enzyme-treated, water-treated and untreated samples were compared. On the whole, size reduction of P. angolensis seeds pretreated with A. niger cellulase enzyme to improve both fat quality and yield implied it as a useful inventory for confectionary as well as cosmetic industries.
Key words: acid value, Aspergillus niger, cellulase, peroxide value, Pycnanthus angolensis, saponification
*Corresponding Authors: Emmanuel Chekwube Ossai, Department of Biochemistry, Faculty of Biological Sciences, University of Nigeria, Nsukka, Enugu State, Nigeria. Email: [email protected];
Charles Odilichukwu R. Okpala, Department of Functional Food Products Development, Wrocław University of Environmental and Life Sciences, Wrocław, Poland. Email: [email protected]
Received: 26 October 2021; Accepted: 2 March 2022; Published: 6 August 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Pycnanthus angolensis (Welw) Warb. (family Myristicaceae) is a widely distributed species of tree in Central and West Africa, with its different parts traditionally serving in management and treatment of wide range of ailments such as mouth ulcer, anemia, malaria, schistosomiasis and fungal infections (Gustafson et al., 2013; Mapongmetsem, 2007; Sofidiya and Awolesi, 2015). The seeds of the plant resemble those of true nutmeg (Myristica fragrans), although the latter belongs to a different genus; hence P. angolensis is literally known as African nutmeg. Its bitter-tasting inedible seeds serve as spice ingredient in Central Africa (Mapongmetsem, 2007). The seeds have an ample fat content of roughly 56–61.6%, with myristic (58.1–64.4%) and myristoleic (19.4–26.3%) acids as their main fatty acid components (Gustafson et al., 2013). The fat, also called kombo butter, is a useful component of cosmetic/soap production, as it is a potential source of cetylmyristoleate (Gustafson et al., 2013) and some terpenoid quinonic acids (Mapongmetsem, 2007), with antioxidant/anti-inflammatory properties. That is why, kombo butter is used extensively in cosmetic/pharmaceutical industries to make ointments used to relieve pain of body joints and muscles. Nonetheless, as common to most plant seeds, fats from P. angolensis seeds is extracted through mechanical and solvent (aqueous, organic) methods (Gustafson et al., 2013; Lourith et al., 2016; Mu et al., 2021; Sirisompong et al., 2011).
However, owing to ever-increasing demand of fats from vegetable sources (Borawski et al., 2016), and compounding urbanization, civilization, and industrialization, there is a requirement to maximize fat yield, especially from oil seeds. A potential strategy is the use of enzymatic pretreatment of seeds and feedstocks for improved yields of fat and oil (Díaz-Suárez et al., 2021; He et al., 2020; Li et al., 2012). Enzymatic breakdown of structural polysaccharides covering cells of fat-bearing seeds would enhance the release of fats/oil stored in the cells, and that also with less energy (Nhi and Tuan, 2016). Hence, this would enhance the penetration of extracting solvent and eventual release of fats into the medium.
As an environment-friendly approach, the enzyme- assisted extraction process would produce fat that would possess high oxidative stability and low phospholipid concentrations. This approach/strategy would minimize product loss during downstream processing (Kalia et al., 2001; Zhang et al., 2011). Despite having advantages, the application of enzyme-assisted fat extraction in relevant industries, especially in developing countries such as Nigeria, has been hampered by long processing time and high cost of enzyme purification. Hence, there is a requirement to explore the use of crude or partially purified enzymes for extraction of fat from seeds (Liu et al., 2016). Studies utilizing enzyme pretreatment for extracting lipids from various plant materials have shown enhancement in lipid recovery. Enzymes such as cellulases, hemicellulases and pectinases, individually or in combination, have been employed in the pretreatment of plant materials for the hydrolysis of cell wall components, thereby increasing their accessibility to extracting solvents and improving the yield of fats/oil (Didia et al., 2018; Nhi and Tuan, 2016; Qiu et al., 2019; Tano-Deborah and Ohta, 1994; Zhang et al., 2018).
Globally, agro-waste remains among the major solid wastes, which include corn cob, corn bagasse, sugarcane bagasse, grape vines, etc., and their use as an alternative source has become the focus of interest for many researchers (Ashori et al., 2014; Atoyebi et al., 2019). Production of amylase from Aspergillus niger has been previously reported through the fermentation of biomass bagasse waste substrates (Aliyah et al., 2017). To the best of our knowledge, no study has investigated how cellulase produced from the fungus A. niger grown from waste material like corn baggase can help to enhance extraction of fat specifically from P. angolensis seeds through the context of particle size of seeds. To supplement the existing information, therefore, the present work investigated the use of partially purified cellulase from A. niger for enzymatic pretreatment of P. angolensis seeds prior to fat extraction, specifically aimed to improve (fat) quality and yield. The hypothesis is that fat yield can be improved based on the use of different particle size seeds. More so and as a cocktail of enzymes, the use of near-crude cellulase extract would play an important role, particularly to avoid the loss of (some) enzyme components that can happen during the extensive purification processes.
Materials and method
Schematic overview of the experimental program
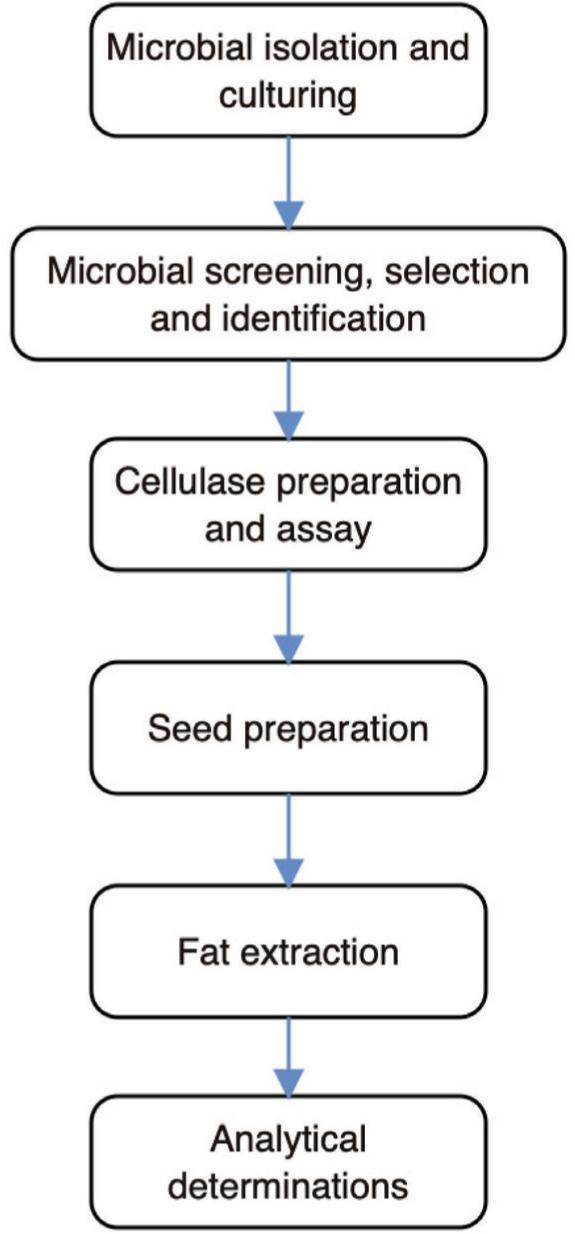
Figure 1 shows the schematic overview of the experimental program, from the microbial isolation and culturing, microbial screening, selection and identification, cellulase preparation and assay to analytical determinations. For emphasis, the present work was purposed to investigate the use of partially purified cellulase from A. niger for the pretreatment of P. angolensis seeds prior to fat extraction for improving yield and quality of fat. Besides, cellulase was to be produced from A. niger using the corn bagasse—a domestic waste which is to serve as the only carbon source. Additionally, the temperature and pH optima for enzyme activity were also ascertained. In all cases, the measurements were performed thrice using different samples. In addition, all chemicals and reagents used were of analytical grade, and were procured from certified retailers.
Figure 1. The schematic overview of the experimental program from the microbial isolation and culturing, microbial screening, selection and identification, cellulase preparation and assay to analytical determination.
Isolation and culturing of microorganism
Soil samples were collected from dumpsites in Nsukka area of Enugu State. The soil was serially diluted and used to inoculate sterilized agar plates using spread plate method. About 0.1 mL of each sample was aseptically transferred to Sabouraud dextrose agar (SDA) plates using a micropipette. Samples were evenly distributed around the plate using a sterilized spreader, and incubated at 30°C for 4 days for fungi growth. Distinct colonies were sub-cultured repeatedly on SDA to get pure fungi isolates.
Screening, selection and identification of cellulase producing fungi
Basal salt medium was prepared according to the method adopted by Pointing (1999). The media were sterilized in an autoclave at 121°C for 15 min, transferred aseptically to sterilized petri dishes and allowed to solidify. The test organisms were inoculated into the media and incubated at 30°C in darkness for 5 days. In order to identify the cellulase-producing fungi, the plates were covered with 2% (w/v) aqueous Congo red stain and left for 20 min. The stain was poured off, and the agar surface was washed with distilled water (Bradner et al., 1999). Thereafter, the plates were inundated with 1-M NaCl for 20 min to remove the stain. The positive result of colonies of cellulase-producing fungi was identified as a yellow-opaque region with red background. Isolated fungi (A. niger) were identified based on cultural and morphological characteristics using lactophenol cotton blue staining method and strain BC23 (species C. elegans) in accordance with Pitt and Hocking’s (2009) taxonomic descriptions.
Culture medium
Culture medium was prepared using dried and ground corn bagasse as the sole source of carbon to produce cellulase from A. niger. Basal salt media prepared according to the Pointing (1999) method were put into separate conical flasks (250-mL capacity), each containing basal medium (100 mL). This was autoclaved at 121°C for 15 min. After cooling, A. niger was introduced. Replicate cultures were left incubated at 30°C for several days. Crude enzyme was harvested from flasks every day, and assays of cellulase activity were conducted for determining the day of maximum enzyme production. In addition, cultures of different concentrations of corn bagasse were incubated to obtain the optimum concentration of substrate for cellulase production.
Cellulase preparation and activity assay
Solid-state fermentation
Cellulase was produced by solid-state fermentation. In all, 25 g of corn bagasse was placed in different flasks. The Pointing (1999) medium was added to each flask and autoclaved. After cooling, the flasks were inoculated and incubated at 30°C for 4 days before extraction of crude enzyme, which was done by adding 0.1-M sodium phosphate buffer (50 mL, pH 6.0) into each flask. This was followed by agitation at 150 rpm and 30°C for 30 min. Slightly wet cheese cloth was used for extract filtration, and the filtrate was centrifuged at 6,000 rpm for 15 min to get clear supernatant (crude enzyme), which was kept at 4°C prior to use.
Cellulase activity assay
The partially purified cellulase was mixed with substrate, 1% carboxymethylcellulose (CMC) in phosphate buffer (pH 6.5) at 50°C for 15 min. At 540 nm, dinitrosalicylic acid (DNS) assay was applied to determine enzyme activity by quantifying the level of reducing sugar produced per milliliter. Similarly, protein content was determined by the Lowry method (Lowry et al., 1951) using BSA as protein standard.
Ammonium sulfate precipitation
Ammonium sulfate precipitation was performed to ascertain the percentage of salt required to precipitate maximum protein from the stock (crude) of enzyme solution. Various concentrations of salt (20% w/v to 100% w/v) were added to crude enzymes (10 mL each) in nine test tubes. After thorough mixing of the content in each tube, they were kept aside at 4°C for 30 h. The eafter, they were centrifuged at 4,000 rpm for 30 min, followed by decanting of supernatants and re-dissolution of pellets in equal volume of 0.1-M phosphate buffer (pH 6.0). Estimation of cellulase activity was done in each tube. The percentage of salt for which the activity of cellulase was highest was noted, and was scaled up to 1 L for mass precipitation of the enzyme for dialysis.
Temperature and pH optima of cellulase activity
The activity of cellulase from A. niger was determined through incubation of the enzyme with CMC solution (1% w/v) through 30–90°C range, at an interval of 10°C for 15 min and at pH 6.0 (0.1-M sodium phosphate buffer). The optimum temperature was obtained from a plot of activity of cellulase against temperature. In addition, the effect of pH on cellulase activity was estimated using 0.1-M sodium acetate buffer, pH 3.5–5.5, phosphate buffer, pH 6.0–7.5, and Tris-HCl buffer, pH 8.0–10.0 at intervals of 0.5. The optimum pH was estimated from a plot of activity of cellulase against pH.
Collection and preparation of the seeds
The P. angolensis seeds used in this study were collected from Umana Ndiuno in Ezeagu Local Government Area of Enugu State, Nigeria. The seeds were screened to remove the damaged ones; thereafter their shells were carefully removed to obtain kernels. The kernels were dried to a constant weight in an oven at 60°C for 96 h, ground using mechanical grinder and sieved into two particle sizes of 0.1 mm and 1.4 mm. Each of the two particle sizes of P. angolensis seeds was divided into three portions of 100 g each. The first portion was treated with 50-mL buffer and 50-mL partially purified enzyme (cellulase). The contents were thoroughly mixed for 1 h before incubation. The second portion was treated with 50-mL buffer and 50-mL distilled water to serve as control and thoroughly mixed for 1 h before incubation. The third portion was left without any treatment. The treated portions were incubated at 50°C for 24 h while the pH values of the treated samples were adjusted to 6.5 using sodium phosphate buffer. At the end of incubation, the treated samples were dried to constant weights in an oven and stored in a desiccator.
Extraction of fat from P. angolensis seeds
Fat extraction from P. angolensis seeds was carried out by the Soxhlet extraction technique using n-hexane as solvent. The percentage fat yield of P. angolensis seeds was determined after triplicate run (each at 65°C for 6 h) and the mean value was calculated. The percentage fat yield was calculated using the following equation below:
Physicochemical properties of fat
Physiochemical properties of fat were determined using the method mentioned by Association of Official Analytical Chemists (AOAC, 2005), which included the following: acid value, free fatty acids, saponification value, iodine value, peroxide value, refractive index and viscosity.
Acid value and determination of free fatty acids
For determining acid value (AV), 5.0 g of fat was taken in a conical flask containing 25-mL absolute ethanol; to this was added 2 drops of phenolphthalein indicator. The mixture was heated with continuous shaking in a water bath at 65°C for 10 min. The resulting solution was cooled and titrated against 0.1-N KOH until a pink color appeared. The AV was estimated following equation below:
The amount of free fatty acids (FFA) was calculated by the following equation below:
FFA = Acid value × 0.503.
Determination of saponification value
In order to determine the saponification value (SV) of fat, 25 mL of 0.5-N alcoholic potassium hydroxide was first added to 2-g fat in a 250-mL capacity conical flask. Thereafter, a reflux condenser was attached and the mixture was heated for 1 h in a boiling water bath, with intermittent shaking. Following the addition 3 drops of phenolphthalein indicator to the warm solution, titration was carried out against 0.1-N HCl. In addition, titration was done against HCl using the blank sample (25-mL alcoholic KOH). Saponification value was estimated using the following equation below:
where B = amount of HCl in milliliter required for the blank sample, and S = amount of HCl in milliliter required for sample.
Determination of iodine value
Determination of iodine value (IV) was done by adding 20-mL carbon tetrachloride and 25-mL Wijs reagent to 0.25-g fat in a 250-mL capacity Erlenmeyer flask. The flask was stoppered, swirled to mix the contents, and then stored in dark for 30 min. Thereafter, 10-mL 30% potassium iodide solution and 100-mL distilled water were added and the mixture was titrated against sodium thiosulfate solution. Following the disappearance of yellow color, 1-mL starch indicator was added and the solution (blue in color) was titrated until the endpoint (solution becomes colorless). Similarly, the procedure was repeated for blank solution (without fat) and the iodine value was calculated following the equation below:
where B = amount of sodium thiosulfate solution in milliliter required for blank sample, and S = amount of sodium thiosulfate solution in milliliter required for sample.
Determination of peroxide value
For determining peroxide value (PV), 30-mL acetic acid–chloroform solution was first added to 5-g fat in a 250-mL capacity stoppered Erlenmeyer flask. The mixture was swirled until the complete dissolution of fat. Thereafter, 0.5-mL solution of saturated potassium iodide was added, and the flask was stoppered and contents swirled for 1 min. Followed with immediate addition of 30-mL distilled water, the flask was restoppered and vigorously shaken to release iodine from the chloroform layer. A deep red-orange colored mixture was then titrated against 0.1-N sodium thiosulfate solution until the disappearance of color. The s was followed by the addition of 1-mL starch solution, and the mixture was further titrated with more of 0.1-N sodium thiosulfate solution until the disappearance of blue-gray color in the upper layer. The procedure was repeated for blank as well as PV (expressed in milli-equivalent of active oxygen per kg of sample) calculated using the following equation below:
where S = amount of sodium thiosulfate solution in milliliter required for sample, and B = amount of sodium thiosulfate solution in milliliter required for blank sample.
Determination of refractive index and specific gravity
Refractive index was determined by using Abbe refract-tometer. In addition, specific gravity (SG) of fat was determined by using a 25-mL capacity density bottle at 50°C. It was calculated as the ratio of the weight of fat to that of equal volume of water at a specific temperature, depicted in the equation below:
Determination of viscosity
Viscosity (η1) was determined using Oswald viscometer. The stated viscometer has two marks, one at top of the bulb and another at the bottom. The sample was drawn from one arm to fill the bulb, after being poured from the opposite arm. On getting to the top mark, a stopwatch was set and the time the sample took to flow down to the bottom mark from the top mark was recorded as t1. This was used to calculate the viscosity using the following the equation below:
where η1 = viscosity of sample, η2 = viscosity of water = 1.005, ρ1 = density of sample and ρ2 = density of water.
Statistical analysis
SPSS version 22 for Windows was used to run the collected data. One way analysis of variance (ANOVA) was applied to the emergent data derived from three measurements taken from different samples. The results were presented in terms of mean ± standard deviation (SD) where appropriate, and the level of statistical significance was set at P < 0.05.
Results and discussion
Enzyme production, ammonium sulfate precipitation profile, and cellulase activities
Incubation time and substrate concentration were the critical factors considered for the efficiency of enzyme production, which Gregg and Saddler (1996) considered while reviewing the factors affecting cellulose hydrolysis and potential of enzyme recycle to enhance efficiency of integrated wood-to-ethanol process. The initial aspect of this study, which was performed to determine the day of peak enzyme production, could be considered to be a pilot feature. Figure 2 shows the effects of (a) incubation period and (b) substrate concentration on enzyme production. The peak enzyme activity (121.036 U/mL) was recorded (Figure 2A) specifically on day 4 of fermentation. This situation suggested a feasible harvest (enzyme) at about 96 h (on day 4) of fermentation, similar to the findings published elsewhere (Abubakar and Oloyede, 2013; Costa et al., 2021; Maftukhah and Abdullah, 2018; Okoye et al., 2013). The incubation period, however, would depend on a number of factors, such as the availability/level of nutrients, fermentation environment, nature of fermenting organisms and the physiological conditions influencing the (fermentation) process (Patil and Dayanand, 2006; Shivasharanappa et al., 2014). With the concentration of substrate (corn bagasse) being optimized, the cellulase activity peaked (121.331 U/mL) when the concentration of the corn bagasse reached 25% w/v (Figure 2B). It is important to observe that the crude cellulase production was conducted by growing the fungi for 4 days in a mineral medium that contained 25% w/v corn bagasse, the latter being the sole carbon source. Afterwards, the medium containing crude cellulase was harvested and purified partially, subsequently.
Figure 2. Effects of (A) incubation period, and (B) substrate concentration on enzyme production.
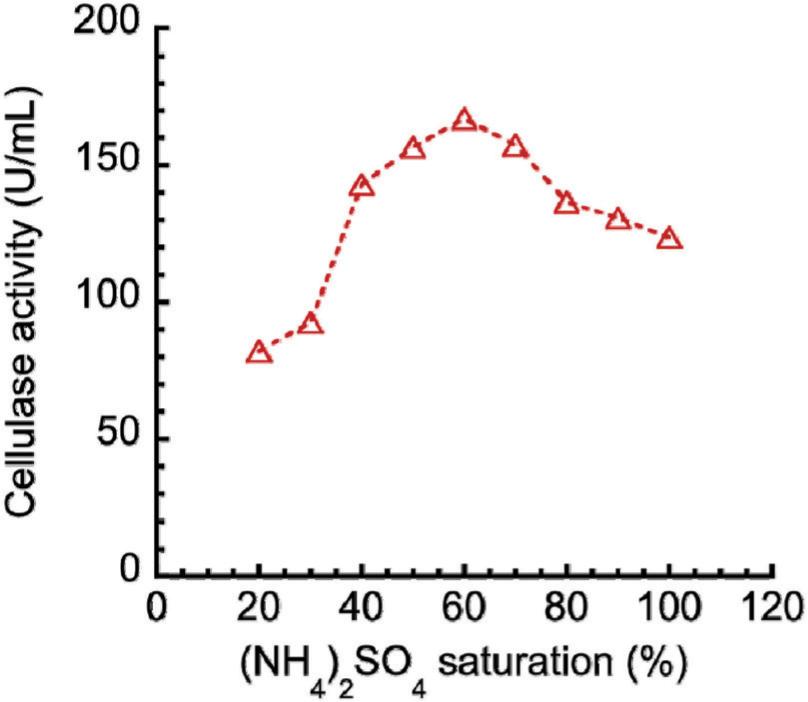
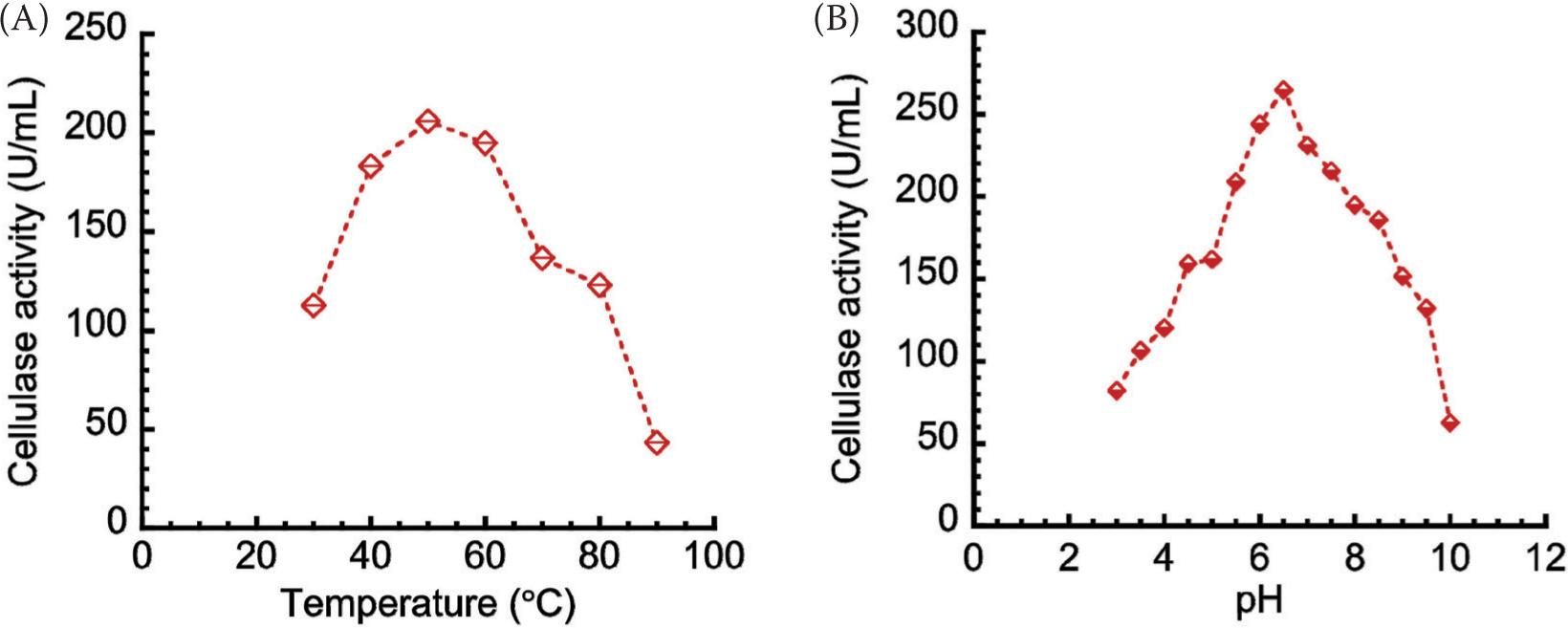
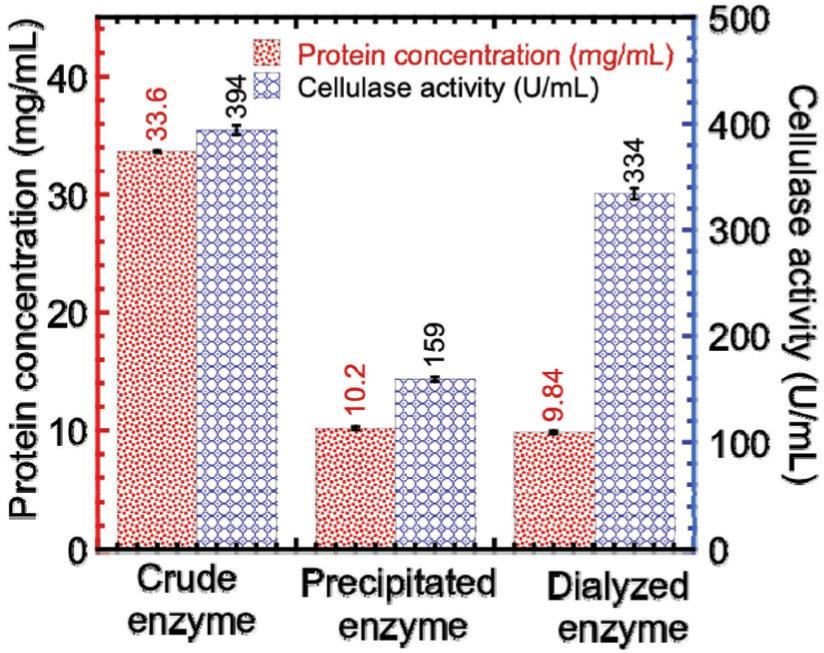
The ammonium sulfate precipitation profile is shown in Figure 3. Given the ammonium sulfate purification at 60% saturation, a maximum cellulase activity seemed apparent, which enabled its use for subsequent stages. After precipitation, the enzyme was dialyzed to remove excess salt. Based on the highest activity because of changes in temperature and pH, the dialyzed enzyme was interpreted. The effects of (a) temperature and (b) pH on cellulase activities are shown in Figure 4. The temperature increased with enzyme activity to attain the peak of 50°C (Figure 4A), which declined subsequently. Temperature increased with average kinetic energy and reaction rate of enzyme molecules. The enzyme activity was inclined to drop gradually after reaching an optimum temperature, suggesting change in enzyme conformation. Possibly, the disulfide bond interactions between hydrophobic domains might be disrupted, which might be followed by denatured/inactivated enzyme. Equally, the optimum cellulase activity (264.73 U/mL) was observed at pH 6.5 (Figure 4B), which suggested enzyme functioning in an acidic environment. Previous studies (Okoye et al., 2013; Srivastava et al., 2017) obtained optimum temperature of 50°C for other cellulase produced from Emericella variecolor and A. niger. Gilna and Khaleel (2011) and Srivastava et al. (2022) also obtained pH 6.5 as the optimum value for other cellulase produced from Cladosporium cladosporioides and Rhizopus oryzae coculture and A. fumigatus. In the present work, crude fractions, and precipitated and dialyzed enzymes were assayed for cellulase activities and protein contents. Comparisons of protein concentrations and activities of crude fractions, and precipitated and dialyzed cellulases are shown in Figure 5. Protein concentration decreased from 33.65 to 10.20 mg/mL after precipitation of ammonium sulfate, and further decreased from 10.20 to 9.84 mg/mL after dialysis, the latter being attributed to the removal of proteins of lower molecular weight. In addition, decrease in cellulase activity from 394.18 to 159.22 U/mL after precipitation but subsequent increase from 159.22 to 333.98 U/mL after dialysis could be due to the removal of impurities to alter enzyme activity.
Figure 3. Ammonium sulfate precipitation profile.
Figure 4. Effect of (A) temperature, and (B) pH on cellulase activities.
Figure 5. Comparisons of protein concentrations and activities of crude, precipitated, and dialyzed cellulase.
Purification profile of cellulases from A. niger
The purification profile of cellulases from A. niger is shown in Table 1. These were the result of protein and enzyme assays of crude as well as precipitated and dialysed enzymes used to determine their respective activities and purification fold. Effect of particle sizes and enzyme treatment on fat yield is shown in Table 2. The percentage fat yield of P. angolensis seeds of particle size 1.0 mm demonstrated the peak of enzyme-treated sample (88.15 ± 0.82%), followed by water-treated sample (76.75 ± 0.42%) and untreated sample, which produced the least yield (72.08 ± 0.95%). The percentage fat yield of enzyme-treated sample appear considerably higher (P < 0.05) compared to water-treated and untreated samples. The same pattern seemed apparent when the particle size of P. angolensis seeds increased to 1.4 mm. The enzyme-treated sample obtained a fat yield of 83.91 ± 0.33%, significantly higher (P < 0.05) than the water-treated sample (74.87 ± 0.55%) and the untreated sample (68.34 ± 2.36%). Fat extracted from the enzyme-treated sample was also higher than the fat yield of 74.13% earlier obtained by Nagre et al. (2011), which asserted enzyme pretreatment with capacity to provide enhanced fat extraction. Similarly, pretreatment of biomass with enzymes prior to lipid extraction increased lipid yield relative to non-enzymatic pretreated samples (Díaz-Suárez et al., 2021; He et al., 2020; Liu et al., 2019). In another study, Otu et al. (2015) obtained a 4% increase in fat yield when the shea tree seeds were treated with crude enzyme. In the present study, the enzyme-treated samples depicted enhanced fat yield in the investigated P. angolensis seeds of two particle sizes. Probably, this could be due to the presence of enzymatic hydrolysis that digested complex cell walls of oil-bearing seeds, which by enhancing the permeability of solvent (n-hexane), favored the oil extraction. Furthermore, the smaller particle size of 1.0 mm, as shown in Table 2, produced higher fat yield in each group of enzyme-treated, water-treated and untreated samples. Reduction in particle size of seeds to have increased fat yield could be anticipated, given the enhanced surface area of smaller particle size (Dash et al., 2021; Ishak et al., 2021).
Table 1. Purification profile of cellulase from A niger.
| Purification steps | Vol (mL) | Protein concentration (mg/mL) | Activity (U/mL) | Total protein (mg) | Total activity (U) | Specific activity (U/mg) | Purification fold |
|---|---|---|---|---|---|---|---|
| Crude enzyme | 1,000 | 33.65 | 394.18 | 33,645 | 394,176 | 11.72 | 1 |
| 60% (NH4)2SO4 | 82 | 10.20 | 159.22 | 835.99 | 13,056 | 15.62 | 1.33 |
| Dialysed enzyme | 101 | 9.84 | 333.98 | 993.84 | 33,732 | 33.94 | 2.89 |
Table 2. Effect of particle size and enzyme treatment on fat yield.
| Particle size | Untreated sample | Fat yield (%) Water-treated sample | Enzyme-treated sample |
|---|---|---|---|
| 1.0 mm | 72.08 ± 1.65a | 76.75 ± 0.72b | 88.15 ± 1.42c |
| 1.4 mm | 68.34 ± 4.09a | 74.87 ± 0.96b | 83.91 ± 0.57c |
Values were mean ± standard deviation of triplicate run. Different superscripts alphabets (a, b and c) across the rows indicate significant difference between mean values at P < 0.05.
Physicochemical properties of extracted fat from P. angolensis seeds
The physicochemical properties of extracted fat from P. angolensis seeds are shown in Table 3. For emphasis, the physicochemical properties involved measurement of acid value, free fatty acid, specific gravity, viscosity, refractive index as well as iodine, saponification and peroxide values. The acid value of fat of the enzyme-treated sample (7.33 ± 0.06 mg KOH/g) was considerably (P < 0.05) higher than that of the water-treated sample (5.95 ± 1.22 mg KOH/g) and the untreated sample (5.72 ± 0.11 mg KOH/g). A considerable increase (P < 0.05) in free fatty acid of the enzyme-treated sample (3.69 ± 0.03%) occurred when compared with the water-treated (2.99 ± 0.11%) and untreated samples (2.88 ± 0.06%). Probably, the cellulase‘s hydrolytic action on the cell walls of oilseeds exposed the lipid bilayer of cell membranes, which might have enhanced the secretion of native seed enzyme lipase. Prolonged activities of lipase to degrade lipid bilayer would likely increase the release of free fatty acids (Kumar et al., 2021).
Table 3. Physicochemical properties of the extracted fat of P angolensis seeds.
| Parameters | Untreated sample | Water-treated sample | Enzyme-treated sample |
|---|---|---|---|
| Acid value (mg KOH/g) | 5.72 ± 0.11a | 5.95 ± 1.22a | 7.33 ± 0.06b |
| Free fatty acid (%) | 2.88 ± 0.06a | 2.99 ± 0.11b | 3.69 ± 0.03c |
| Saponification value (mg KOH/g) | 223.37 ± 3.24a | 227.11 ± 7.07a,b | 236.46 ± 2.81b |
| Iodine value (gI2/g) | 62.99 ± 0.78a | 64.18 ± 1.06a | 65.36 ± 2.21a |
| Peroxide value (meq/kg) | 32.63 ± 1.53a | 27.97 ± 2.08b | 14.63 ± 1.53c |
| Specific gravity | 0.893 ± 0.010a | 0.864 ± 0.010a | 0.856 ± 0.010b |
| Viscosity (cP) | 2.498 ± 0.000a | 1.520 ± 0.037b | 1.369 ± 0.002c |
| Refractive index | 1.4927 ± 0.04a | 1.4856 ± 0.04b | 1.4811 ± 0.02c |
| Color | Reddish-brown | Reddish-brown | Reddish-brown |
Values were mean ± standard deviation of triplicate run. Different superscripts alphabets (a, b and c) across the rows indicate significant difference between mean values at P < 0.05.
Saponification value revealed the average molecular weight of triacylglycerol components of fat. Higher saponification values indicated shorter chain fatty acids in ester bonds with glycerol backbone. Despite that no noticeable difference (P > 0.05) occurred between the saponification values of fat of the enzyme-treated (236.46 ± 2.81 mg KOH/g) and water-treated (227.11 ± 7.06 mg KOH/g) samples, that of untreated (223.37 ± 3.24 mg KOH/g) sample appeared considerably (P < 0.05) lower (Table 3). Besides that Nagre et al. (2011) reported a saponification value of 238.0 mg KOH/g fat in P. angolensis seeds, other workers showed virgin coconut oil extracted using different processes would produce higher saponification values (254.10–264.04 mg KOH/g) highest at the enzyme-assisted method produced the highest values (Mohammed et al., 2021). Since the saponification values in this study were lower than those reported for virgin coconut oil, those from P. angolensis fat would accumulate in lower proportions of low molecular weight fatty acids; this suggests a smaller acidity content, and hence better feedstock for soap and confectionary applications.
Table 3 also reveals that, despite resemblances (P > 0.05), the iodine value of fat of the enzyme-treated samples was slightly higher (65.36 ± 2.21 g I2/g) than the water-treated (64.18 ± 1.06 g I2/g), followed thereafter by untreated (62.99 ± 0.78 g I2/g) sample. Further, these iodine values compared well with the data (64.9 g I2/g) of P. angolensis seeds reported by Nagre et al. (2011). Similarly, a higher iodine value was indicated for enzyme-assisted oil extracted from S. mukorossi seed kernels (113.15 ± 3.01 g I2/100 g) compared with the iodine value of solvent-extracted oil (107.39 ± 2.64 g I2/100 g) (Liu et al., 2019). Owing to low iodine values observed in the present study, the fat of P. angolensis seeds could be considered as nondrying lipid with promising storability. Relatively low degree of unsaturation of fat would depict an improved thermo-oxidative stability, which can potentially serve as biolubricants and bioplasticizers (Cecilia et al., 2020; Ikhuoria and Maliki, 2007).
The peroxide values of fat of enzyme-treated (14.63 ± 1.53 meq/kg), water-treated (27.97 ± 2.08 meq/kg) and untreated (32.63 ± 1.53 meq/kg) samples differed considerably (P < 0.05) (Refer to Table 3). The less peroxide value of the enzyme-assisted extracted fat suggested higher resistance to oxidative rancidity, and thus a good quality oil (Mohammed et al., 2021; Nandi and Bhattacharyya, 2015). Resistance to oxidative deterioration of enzyme-treated sample would help to substantiate the oxidative stabilities (Elagbar et al., 2016), given the presence of natural antioxidants. Moreover, the above situation is compared with onion oil extracted with cellulase with high phenolic content, which is an antioxidant, as reported by Salina et al. (2013). Thus, the enzyme used to enhance fat extraction could help liberate antioxidants (Nhi and Tuan, 2016). Besides the high antioxidant content of P. angolensis fat (Nagre et al., 2011), natural antioxidants could help to reduce the peroxide value of oil/fat (Ullah et al., 2003; Zhang et al., 2021).
Additionally, the specific gravity of fat resembled (P > 0.05) across enzyme-treated (0.856 ± 0.10), water-treated (0.864 ± 0.10) and untreated (0.893 ± 0.09) samples (Table 3). Specific gravity values of fat observed in the present work were less than the data reported by Nagre et al. (2011) for P. angolensis (0.922), which had been obtained via solvent extraction. Reduced specific gravity of fat of the enzyme-treated sample could be due to its lower molecular weight, considering its saponification values. At the temperature of 50°C, the viscosity of fat differed significantly (P < 0.05) between enzyme-treated (1.369 ± 0.02 cP), water-treated (1.520 ± 0.04 cP) and untreated (2.498 ± 0.04 cP) samples (Table 3). The least value of viscosity of fat of the enzyme-treated sample could be due to its slightly higher degree of unsaturation, reinforced by its iodine value. This phenomenon demonstrated that oil viscosity depended on the structure of molecules and increased with the saturation of fatty acid (Talkit et al., 2012). Additionally, the refractive index of fat of enzyme-treated (1.481 ± 0.0019), water-treated (1.485 ± 0.0005) and untreated (1.493 ± 0.0003) samples also differed considerably (P < 0.05). Decrease in the refractive index of enzyme-treated sample was probably at the highest level of unsaturation, not only as reflected by the iodine value but also by shortened fatty-acyl chain components as well as higher saponification number (Bayisa and Bullo, 2021). By increased refractive index of enzyme-treated samples, the fat hydrolysis of native enzyme lipase indicated a higher free fatty acid. Compared to their constituent free acids, however, the triacylglycerols possessed higher refractive indices (Ibeto et al., 2012).
Conclusions
The use of partially purified cellulase from A. niger for the enzymatic pretreatment of P. angolensis seeds prior to fat extraction, targeting improvement of (fat) yield and quality, was investigated. For emphasis, the fat yield was aimed to be enhanced based on the use of seeds of different particle size. The study demonstrated that P. angolensis seeds were high fat-yielding seeds, which could serve as a commercially rich source of fat. Pretreatment of P. angolensis seeds with cellulase and reduction in their particle size were found to improve fat yield. The physicochemical properties of enzyme-treated, water-treated and untreated seed samples differed significantly, indicating a promising impact of processing methods on quality of extracted fat. The fat obtained from enzyme-treated seeds was high, with appreciable saponification value and high oxidative stability, and thus could find applications in confectionery, soap and cosmetic industries.
We believe that this was the first study to specifically employ partially purified cellulase from A. niger for enzymatic pretreatment of P. angolensis seeds prior to fat extraction, thus targeting improvement in fat yield and quality. The work also established that corn bagasse, a common domestic waste, could be used as a sole carbon source of cellulase production from A. niger. The decreased refractive index of enzyme-treated samples would suggest unsaturation at its peak, supported by their saponification and iodine values, and shorter chain fatty acid components. Size reduction and treatment of P. angolensis seeds with A. niger cellulase improved both fat quality (improved saponification number, iodine value and refractive index) and yield, which suggested their potential application in cosmetic and confectionary processes. Considering the high levels of acidity, the extracted fat is not suitable for direct human consumption, and the future studies must target to ease this challenge.
Acknowledgements
The authors thank Profs. FC Chilaka and SOO Eze, both from the Department of Biochemistry, University of Nigeria, Nsukka, Nigeria, for their useful suggestions for the success of this work. The authors also thank the technical staff of the Department of Biochemistry, University of Nigeria, Nsukka, Nigeria, for their assistance in laboratory. No specific grant was received from any funding agency for this research.
Conflict of interest
The authors declared that they had no competing interest to disclose.
REFERENCES
Abubakar F.A. and Oloyede O.B., 2013. Production and activity of cellulase from Aspergillus niger using rice bran and orange peel as substrates. International Journal of Scientific Research and Management 1(5): 285–291.
Aliyah, A., Alamsyah, G., Ramadhani, R. and Hermansyah, H., 2017. Production of α-amylase and β-glucosidase from Aspergillus niger by solid state fermentation method on biomass waste substrates from rice husk, bagasse and corn cob. Energy Procedia 136, 418–423. 10.1016/j.egypro.2017.10.269
Ashori, A., Nourbakhsh, A. and Tabrizi, A.K., 2014. Thermoplastic hybrid composites using bagasse, corn stalk and E-glass fibers: fabrication and characterization. Polymer-Plastics Technology and Engineering, 53(1), 1–8. 10.1080/03602559.2013.832854
Association of Official Analytical Chemists (AOAC). 2005. Official methods of analysis, 18th edition. AOAC, MD, USA.
Atoyebi, O.D., Osueke, C.O., Badiru, S., Gana, A.J., Ikpotokin, I., Modupe, A.E. and Tegene, G.A., 2019. Evaluation of particle board from sugarcane bagasse and corn cob. International Journal of Mechanical Engineering and Technology (IJMET) 10(1), 1193–1200.
Bayisa Y.M. and Bullo T.A., 2021. Optimization and characterization of oil extracted from Crotonmacrostachyus seed for antimicrobial activity using experimental analysis of variance. Heliyon 7: e08095. 10.1016/j.heliyon.2021.e08095
Borawski A.B., Borawski P. and Dunn J.W., 2016. Factors of the development of oilseed rape production in Poland on the background of the world. Journal of Advanced Agricultural Technologies 3(3): 175–179. 10.18178/joaat.3.3.175–179
Bradner J.R., Gillings M. and Nevalainen K.M.H., 1999. Qualitative assessment of hydrolytic activities in Antarctic micro fungi grown at different temperatures on solid media. World Journal Microbiology and Biotechnology 15: 131–132. 10.1023/A:1008855406319
Cecilia J.A., Plata D.B., Saboya R.M.A., Luna F.M.T., Cavalcante C.L., Rodriguez-Castellon E., 2020. An overview of the biolubri-cant production process: Challenges and future perspectives. Processes 8: 257. 10.3390/pr8030257
Costa P.C., Reis E.E, Carvalho L.S., Fontana R.C., Schneider W.D.H. and Camassola M., 2021. Making the process of enzyme production in solid-state cultivation cleaner and more sustainable—reuse of raw materials and a syringe-type bioreactor enter in the scene. ACS Sustainable Chemistry & Engineering 42, 14134–14142. 10.1021/acssuschemeng.1c04257
Dash D.R., Pathak S.S. and Pradhan R.C., 2021. Extraction of oil from Terminalia chebulakernel by using ultrasound technology: influence of process parameters on extraction kinetics. Industrial Crops & Products 171: 113893. 10.1016/j.indcrop.2021.113893
Díaz-Suárez P., Rosales-Quintero A., Fernandez-Lafuente R., Pola-Sanchez E., Hernandez-Cruz M.C., Ovando-Chcon S.L., Rodrigues R.C. and Tacias-Pascacio V.G., 2021. Aqueous enzymatic extraction of Ricinuscommunis seeds oil using Viscozyme L. Industrial Crops and Products 170: 113811. 10.1016/j.indcrop.2021.113811
Didia B., Zakpaa H.D., Mills-Robertson F.C. and Abdul-Mumeen I, 2018. Qualitative analysis of enzyme-assisted traditionally extracted shea butter from different pretreated shea kernel substrates. Journal of Cereals and Oilseeds 9(2): 10–19. 10.5897/JCO2017.0177
Elagbar Z.A., Naik R.R., Shakya A.K. and Bardaweel S.K., 2016. Fatty acids analysis, antioxidant and biological activity of fixed oil of Annonamuricata L. seeds. Journal of Chemistry. 2016: ID 6948098. 10.1155/2016/6948098
Gilna VV. and Khaleel K.M., 2011. Cellulase enzyme activity of Aspergillus fumigatesfrom mangrove soil on lignocellulosic substrate. Recent Research in Science and Technology 3(1): 132–134.
Gregg, D.J. and Saddler, J.N., 1996. Factors affecting cellulose hydrolysis and the potential of enzyme recycle to enhance the efficiency of an integrated wood to ethanol process. Biotechnology and Bioengineering 51(4), 375–383.
Gustafson K., Wu Q.L., Asante-Dartey J. and Simon J.E., 2013. Pycnanthus angolensis: bioactive compounds and medicinal applications. In: Juliani H.R., Simon J.E. and Ho C.T. (eds.). African natural plant products, vol II: Discoveries and challenges in chemistry, health and nutrition. ACS Sumposium Series, Ashington DC, pp. 63–78. 10.1021/bk-2013-1127.ch005
He Y., Zhang B., Guo S., Guo Z., Chen B. and Wang M., 2020. Sustainable biodiesel production from the green microalgae Nannochloropsis: novel integrated processes from cultivation to enzyme-assisted extraction and ethanolysis of lipids. Energy Conversion and Management 209: 112618. 10.1016/j.enconman.2020.112618
Ibeto C.N., Okoye C.O.B. and Ofoefule A.U., 2012. Comparative study of the physicochemical characterization of some oils as potential feedstock for biodiesel production. ISRN Renewable Energy 2012: ID 621518. 10.5402/2012/621518
Ikhuoria E.U. and Maliki M., 2007. Characterization of avocado pear (Persea americana) and African pear (Dacryodes edulis) extract. African Journal of Biotechnology 7: 950–952.
Ishak I., Hussain N., Coorey R. and Ghani M.A., 2021. Optimization and characterization of chia seed (Salvia hispanica L.) oil extraction using supercritical carbon dioxide. Journal of CO2 Utilization 45: 101430. 10.1016/j.jcou.2020.101430
Kalia V.C., Rashmi L.S., and Gupta M.N., 2001. Using enzymes for oil recovery from edible seeds. Journal of Scientific & Industrial Research 60: 298–310.
Kumar R.R., Bhargava D.V., Pandit K., Goswami S., Shankar S.M., Singh S.P., Rai G.K., Satyavathi C.T. and Praveen S., 2021. Lipase—the fascinating dynamics of enzyme in seed storage and germination—a real challenge to pearl millet. Food Chemistry 361: 130031. 10.1016/j.foodchem.2021.130031
Li F., Yang L., Zhao T., Zhao J., Zou Y. and Wu X., 2012. Optimization of the enzymatic pretreatment for n-hexane extraction of oil from Silybum marianum seed using response surface methodology. Food and Bioproduct Processing 90(2): 87–94. 10.1016/j.fbp.2011.02.010
Liu J., Gasmalla M.A.A., Li P. and Yang R., 2016. Enzyme-assisted extraction processing from oilseeds: principle, processing and application. Innovative Food Science and Emerging Technologies 35: 184–193. 10.1016/j.ifset.2016.05.002
Liu Z., Gui M., Xu T., Zhang L., Kong L., Qin L. and Zou Z., 2019. Efficient aqueous enzymatic-ultrasonication extraction of oil from Sapindusmukorossi seed kernels. Industrial Crops & Products 134: 124–133. 10.1016/j.indcrop.2019.03.065
Lourith N., Kanlayavattanakul M., Mongkonpaibool K., Butsaratrakool T. and Chinmuang T., 2016. Rambutan seed as a new promising unconventional source of specialty fat for cosmetics. Industrial Crops and Products 83: 149–154. 10.1016/j.indcrop.2015.12.045
Lowry O.H., Rosebrough N.J., Farr A.L. and Randall R.J., 1951. Protein measurement with the folin phenol reagent. Journal of Biology and Chemistry 193: 265–275.
Maftukhah S. and Abdullah A., 2018. Cellulase enzyme production from rice straw using solid state fermentation and fungi Aspergillus niger ITBCC L74. MATEC Web of Conferences 156, 01010. 10.1051/matecconf/201815601010
Mapongmetsem P.M., 2007. Pycnanthus angolensis (Welw.) Warb. In: Vossen H.A.M. and Mkamilo G.S. (editors). Plant resources of tropical Africa, 14: Vegetable oils. Backhuys, Wageningen, the Netherlands, pp. 137–141.
Mohammed N.K., Samir Z.T., Jassim M.A. and Saeed S.K., 2021. Effect of different extraction methods on physicochemical properties, antioxidant activity, of virgin coconut oil. Materials Today: Proceedings 42: 2000–2005. 10.1016/j.matpr.2020.12.248
Mu J., Wu G., Chen Z., Brennan C.S., Tran K., Dilrukshi H.N.N., Shi C., Zhen H. and Hui X., 2021. Identification of the fatty acids profiles in supercritical CO2 fluid and Soxhlet extraction of Samara oil from different cultivars of Elaeagnus mollis Diels seeds. Journal of Food Composition and Analysis 101: 103982. 10.1016/j.jfca.2021.103982
Nagre R.D., Oduro I. and Ellis W.O., 2011. Comparative physicochemical evaluation of kombo kernel fat produced by three different processes. African Journal of Food Science and Technology 2(4): 83–91.
Nandi S. and Bhattacharyya R., 2015. A parametric study for the enzymatic extraction of rice bran oil and its optimization. International Journal on Recent and Innovation Trends in Computing and Communication 3 (2): 21–25.
Nhi T.T.Y. and Tuan D.Q., 2016. Enzyme-assisted extraction of gac oil (Momordica cochinchinensis Spreng) from dried aril. Journal of Food and Nutrition Sciences 4(1): 1–6. 10.11648/j.jfns.20160401.11
Okoye I.G., Ezugwu A.L., Udenwobele D.I., Eze S.O.O., Anyawu C.U. and Chilaka F.C., 2013. Production and partial characterization of cellulases from Apergillus fumigates using two distinct parts of corn cob as carbon sources. Nigerian Journal of Biotechechnology 26: 50–59.
Otu S.A., Dzogbefia V.P., Kpikpi E.N. and Essuman E.K., 2015. Comparative effect of crude and commercial enzyme in shea fat extraction. IOSR Journal of Biotechnology and Biochemistry 1(3): 18–27.
Patil S.R. and Dayanand A., 2006. Production of pectinase from deseeded sunflower head by Aspergillus niger in submerged and solid-state conditions. Bioresources Technology 97: 2054–2058. 10.1016/j.biortech.2005.09.015
Pitt J.I. and Hocking A.D., 2009. Aspergillus and related teleo-morphs. In: Fungi and food spoilage, Pitt J.I. and Hocking A.D., Editors, Springer, Boston, MA, pp. 275–337. 10.1007/978-0-387-92207-2_8
Pointing S.P., 1999. Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Diversity 2: 17–33.
Qiu C., He Y., Huang Z., Li S., Huang J., Wang M. and Chen B., 2019. Lipid extraction from wet nannochloropsis biomass via enzyme-assisted three phase partitioning. Bioresource Technology 284: 381–390. 10.1016/j.biortech.2019.03.148
Salina H.F., Shima A.R.R., Masniza M. and Faeizah H.N., 2013. Enzyme-assisted aqueous extraction and phenolic antioxidants of onions oil. International Journal of Science, Environment and Technology 2(5): 949–955.
Shivasharanappa K., Hanchinalmath J.V., Sundeep Y.S., Borah D. and Talluri V.P. 2014. Optimization and production of alkaline proteases from agro byproducts using a novel trichoderma viridiae strain VPG 12, isolated from agro soil. International Letters of Natural Sciences 14: 77–84. 10.18052/www.scipress.com/ILNS.14.77
Sirisompong W., Jirapakkul W. and Klinkesorn U., 2011. Response surface optimization and characteristics of rambutan (Nepheliumlappaceum L.) kernel fat by hexane extraction. LWT–Food Science and Technology 44: 1946–1951. 10.1016/j.lwt.2011.04.011
Sofidiya M.O. and Awolesi A.O., 2015. Antinociceptive and antiulcer activities of Pycnanthus angolensis. Revista Brasileira de Farmacognosia 25: 252–257. 10.1016/j.bjp.2015.05.004
Srivastava N., Singh R., Mohammad A., Pal D.B., Syed A., Elgorban A.M., Mishra P.K., Yoon T., Srivastava M. and Gupta V.K., 2022. Graphene oxide mediated enhanced cellulase production using pomegranate waste following co-cultured condition with improved pH and thermal stability. Fuel 312: 122807. 10.1016/j.fuel.2021.122807
Srivastava N., Srivastava, M., Manikanta A., Singh P., Ramteke P.W., Mishra P.K. and Malhotra B.D., 2017. Production and optimization of physicochemical parameters of cellulase using untreated orange waste by newly isolated Emericella variecolor NS3. Applied Biochemistry and Biotechnology 183(2): 601–612. 10.1007/s12010-017-2561-x
Talkit K.M., Mahajan D.T. and Masand V.H., 2012. Study on physicochemical properties of vegetable oils and their blends use as possible ecological lubricant. Journal of Chemical and Pharmaceutical Research 4(12): 5139–5144.
Tano-Deborah K. and Ohta Y., 1994. Enzyme-assisted aqueous extraction of fat from kernels of the shea tree, Butyrospermum parkii. Journal of the American Oil Chemists’ Society 71: 979–983. 10.1007/BF02542265
Ullah J., Hamayoun M., Ahmad T., Ayub M. and Zafarullah M., 2003. Effect of light, natural and synthetic antioxidants on stability of edible oil and fats. Asian Journal of Plant Sciences 2: 1192–1194. 10.3923/ajps.2003.1192.1194
Zhang Y., Kong X., Wang Z., Sun Y., Zhu S., Li L. and Lv P., 2018. Optimization of enzymatic hydrolysis for effective lipid extraction from microalgae Scenedesmus sp. Renewable Energy 125: 1049–1057. 10.1016/j.renene.2018.01.078
Zhang S.B., Lu Q.Y., Yang H., Li Y. and Wang S., 2011. Aqueous enzymatic extraction of oil and protein hydrolysates from roasted peanut seeds. Journal of American Oil Chemists’ Society 88(5): 727–732. 10.1007/s11746-010-1711-x
Zhang C., Zhao J., Famous E., Pan S., Peng X. and Tian J., 2021. Antioxidant, hepatoprotective and antifungal activities of black pepper (Piper nigrum L.) essential oil. Food Chemistry 346: 128845. 10.1016/j.foodchem.2020.128845