
Download
ORIGINAL ARTICLE
Variation of chlorophyll and carotenoids in different varieties and organs of Chinese kale
Chenlu Zhang†, Sha Liang†, Yilin Wang†, Sha Luo, Weijia Yao, Hao He, Yuxiao Tian, Huanxiu Li, Fen Zhang*, Bo Sun*
College of Horticulture, Sichuan Agricultural University, Chengdu, China
Abstract
The concentrations of chlorophyll and carotenoids were analyzed in two organs (leaves and bolting stems) of 19 varieties of Chinese kale (including four maturity periods and two flower colors). Two chlorophylls and four carotenoids were identified in Chinese kale. The concentrations of chlorophyll and carotenoids varied widely in different organs and varieties. JL-03 was a good candidate for the future breeding programs, since it contained the highest concentrations of chlorophyll and carotenoids (except violaxanthin) in leaves. Lutein was the main component of carotenoids and accounted for approximately 50% of total carotenoids. The concentrations of chlorophyll and carotenoids in leaves significantly exceeded those in bolting stems. Principal component analysis showed that organ was the main source of differences of chlorophyll and carotenoid concentrations in different varieties of Chinese kale, while the maturity and flower color have little effect. Correlation analysis identified a significantly positive correlation between chlorophyll and carotenoids in Chinese kale. These results provided evidence for improving human dietary nutrition and breeding of Chinese kale.
Key words: carotenoids, Chinese kale, chlorophyll, organs, varieties
*Corresponding authors: Fen Zhang and Bo Sun, College of Horticulture, Sichuan Agricultural University, Chengdu 611130, China. Emails: [email protected]; [email protected]
†Contributed equally to this work and are first co-authors.
Received: 22 February 2022; Accepted: 14 June 2022; Published: 15 October 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Chinese kale (Brassica oleracea var. alboglabra) belongs to the Brassicaceae family, and its edible parts are bolting stems and tender leaves (Sun et al., 2012a, 2019). It is rich in bioactive compounds, including vitamin C, total phenolics, glucosinolates and carotenoids, which can enhance the immunity in consumers and reduce their risk of cancer (Sun et al., 2011, 2012b). Chinese kale belongs to long-day plants and favors a cold, cool and humid climate. It is an important vegetable crop in South China and is cultivated widely (Lei et al., 2017).
Chlorophyll is the major photosynthesis pigment and is widely distributed in green plants (Zhang et al., 2019). Chlorophyll and its various derivatives can be absorbed by the human body in diet rich in chlorophyll pigments, such as green vegetables. In addition, chlorophyll pigments have numerous health benefits (Ferruzzi and Blakeslee 2007), such as their good ability to chelate with several chemical carcinogens and mutagens (Jubert et al., 2009), reduce cancer risk (Cervantes-Paz et al., 2014) and scavenge free radicals (Chen and Roca, 2018).
Carotenoids are the second most abundant natural pigments with more than 750 members (Quinlan et al., 2012). Carotenoids are isoprenoid metabolites that are synthesized by all photosynthetic organisms. They are essential for photoprotection and participate in light harvesting in photosynthetic systems (Rodriguez-Concepcion et al., 2018). The colors of carotenoids range from colorless to yellow, orange and red, with variations reflected in many flowers, fruits and vegetables (Cazzonelli and Pogson, 2010). Carotenoids play an important role in both plants and humans (Amin et al., 2009). In plants, carotenoids and their derivatives participate in many critical biological functions, such as biosynthesis of phytohormones, photosynthesis and photo-protection (Cazzonelli and Pogson, 2010). However, in human body, carotenoids could enhance the antioxidant and anti-inflammatory activity, and prevent several reactive oxygen species (ROS)-mediated chronic diseases (Rodriguez-Concepcion et al., 2018). Previous studies have reported that carotenoids can prevent and treat several cancers and chronic diseases (Amin et al., 2009). Lutein and zeaxanthin can help to prevent eye diseases (Caseli et al., 2013). In addition, several of the carotenoids can be used as an origin of vitamin A. β-carotene is the most important precursor of vitamin A, which can be degraded into vitamin A when the human body requires it. It can cure dry eye disease, night blindness and other diseases caused by a lack of vitamin A. In general, the antioxidant properties of chlorophyll and carotenoids are associated with a lower risk of developing cancer (Britton and Khachik, 2009).
Chinese kale has a number of varieties with a long history of cultivation. Based on maturity, it can be divided into prematurity, medium maturity, late maturity and extremely late maturity types. It can also be classified into two types according to whether the petals are white or yellow (Lei et al., 2017). Numerous studies have reported large differences in phytochemicals among different species, types, varieties and organs of vegetables (Chen et al., 2019; Sun et al., 2011; Takaidza et al., 2018). However, no studies have been reported on pigments of Chinese kale to date. The present experiment measured the differences of chlorophyll and carotenoids content in leaves and bolting stems of 19 Chinese kale varieties with different maturity and color to explore the effects of variety and organ on the pigment content of Chinese kale.
Materials and Methods
Plant materials
The experiment investigated 19 varieties of Chinese kale, including four maturity types (prematurity, medium maturity, late maturity and extremely late maturity) and two flower colors (white and yellow) (Table 1). These varieties of Chinese kale were sown on 10 September 2015, with a row spacing of 30 cm and a plant spacing of 40 cm. A total of 10,000 kg of decomposed fertilizer was applied to each hectare of field planting. After four days of field planting, 75 kg of urea was applied to each hectare, with 150 kg of urea applied to each hectare every 15 days and watered at the same time. Carbendazim was sprayed at seedling stage to prevent sclerotinia and soft rot, and metoclopramide was sprayed to control diamondback moth. All plants were harvested according to different maturities on 15 November 2015, 5 December 2015, 5 January 2016 and 25 January 2016. At each harvesting, strong and healthy plants with identical maturity levels and no pests and diseases were chosen and transported to laboratory immediately. Fifteen plants were collected per variety, five plants were pooled as one repetition, and three replicates were used in total. After wiping the material surface clean, samples were divided according to edible parts into leaves and bolting stems. After that, all samples were lyophilized for 72 h using a vacuum freeze dryer (Songyuan Huaxing, Beijing, China), and grounded to a fine powder and stored at -80°C.
Table 1. Maturity and flower color in different varieties of Chinese kale.
| Variety | Maturity | Flower color |
|---|---|---|
| JL-01 | Prematurity | White |
| JL-02 | Prematurity | White |
| JL-03 | Prematurity | White |
| JL-05 | Prematurity | White |
| JL-06 | Prematurity | White |
| JL-07 | Prematurity | White |
| JL-08 | Prematurity | White |
| JL-11 | Prematurity | White |
| JL-13 | Medium maturity | White |
| JL-14 | Medium maturity | White |
| JL-16 | Medium maturity | White |
| JL-17 | Medium maturity | White |
| JL-18 | Late maturity | White |
| JL-19 | Late maturity | Yellow |
| JL-21 | Late maturity | White |
| JL-22 | Late maturity | Yellow |
| JL-24 | Extremely late maturity | Yellow |
| JL-25 | Extremely late maturity | White |
| JL-26 | Extremely late maturity | White |
Analysis of chlorophyll and carotenoids
Polypropylene tubes containing 0.2 g of sample powder were filled with 25 mL of acetone for extraction in darkness at 4°C for 12 h. Samples were then centrifuged to 8,000 g at 4°C for 10 min. Cellulose acetate filters (0.22 μm) were used to filter supernatant into a brown chromatographic flask. The high-performance liquid chromatography (HPLC) analysis of chlorophyll and carotenoids was conducted using an Agilent 1260 instrument equipped with a model variable wavelength absorbance detector (VWD). The samples (10 μL) were separated at 30°C on a Waters Spherisorb C18 column (150 × 3.9 mm) using 80% acetonitrile–water and isopropanol at a flow rate of 0.5 mL min–1. Absorbance was detected at 428 and 448 nm (Shi et al., 2014). The peaks of the samples were qualitatively assessed based on the retention period of individual standards. The standards of chlorophylls (a and b) and carotenoids (neoxanthin, violaxanthin, lutein and β-carotene) were obtained from Solarbio Science & Technology Co. Ltd. (Beijing, China). Chlorophyll and carotenoid concentrations in the samples were calculated according to the standard curve of standards.
Statistical analysis
The raw data were processed by Excel 2018, and the results were shown as mean values of three replicates. The statistical analysis and correlation analysis were performed using SPSS 18.0. The least significant difference test was applied to assess differences (P < 0.05). In order to determine relationships among samples, principal component analysis (PCA) and partial least-square discriminant analysis (PLS-DA) were performed using SIMCA-P 11.5 with unit variance scaling. Variance and correlation analyses were also performed. Hierarchical clustering analyses (HCA) were conducted using the TIGR MeV software (version 4.1) (Sun et al., 2018).
Results
Identification of chlorophyll and carotenoids
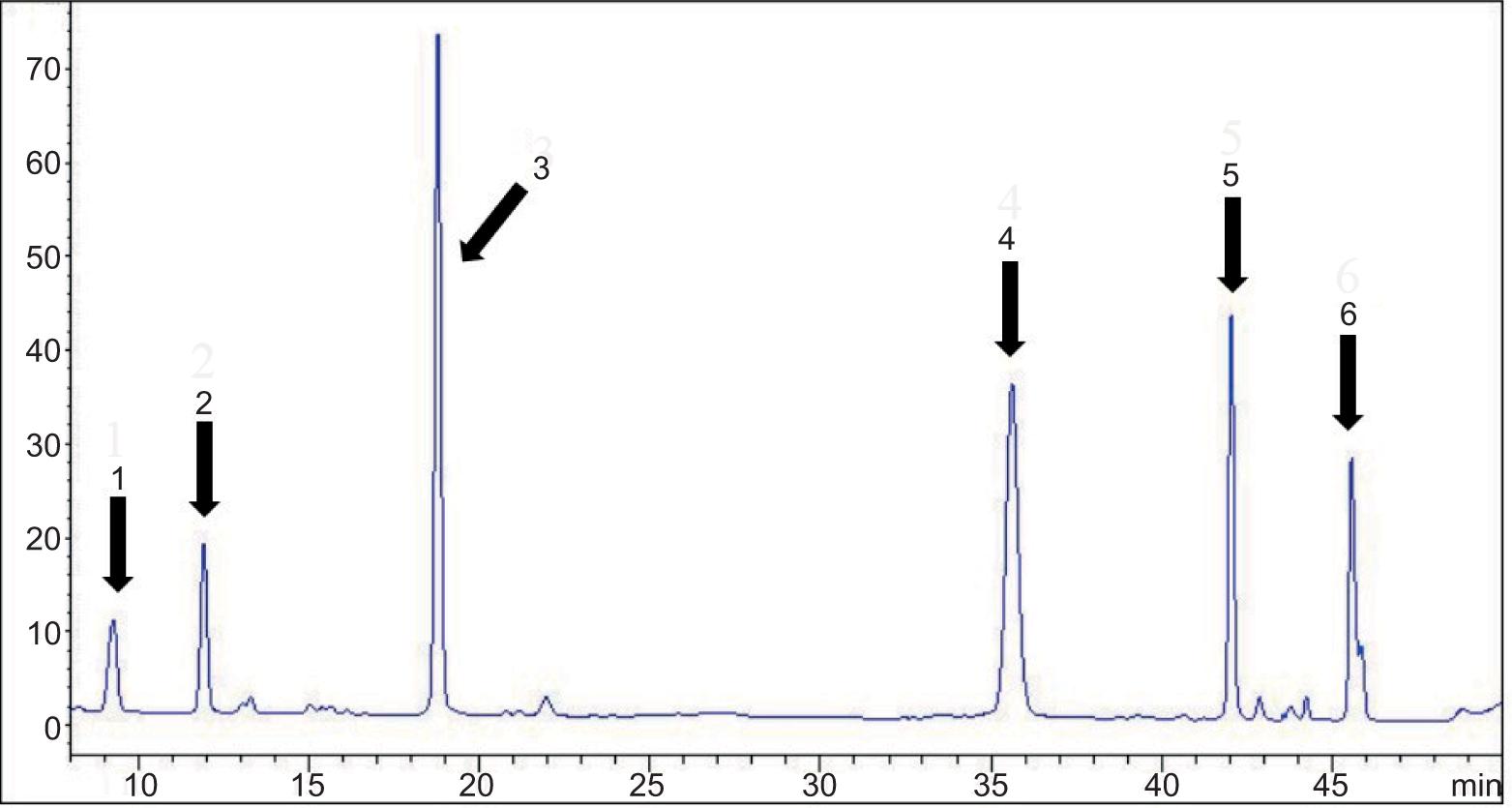
The composition and concentrations of chlorophyll and carotenoids in leaves and bolting stems of Chinese kale were analyzed by HPLC. Figure 1 shows that both chlorophyll a and chlorophyll b and four carotenoids (neoxanthin, violaxanthin, lutein and β-carotene) were detected in all samples of 19 varieties and two organs.
Figure 1. HPLC of chlorophyll and carotenoids in the leaves of Chinese kale. 1: neoxanthin, 2: violaxanthin, 3: lutein, 4: chlorophyll b, 5: chlorophyll a and 6: β-carotene.
Chlorophyll concentrations in leaves and bolting stems of Chinese kale
The leaves of distinct varieties of Chinese kale had significant differences in chlorophyll concentrations, and the distribution of chlorophyll a, chlorophyll b and total chlorophyll was identical (Table 2). The lowest concentrations of chlorophyll a (3.11 mg g–1 DW), chlorophyll b (2.14 mg g–1 DW) and total chlorophyll (5.24 mg g–1 DW) were observed in JL-06, while the highest levels of all these were found in JL-03 (12.60, 9.40 and 22.01 mg g–1 DW, respectively). These were four-fold higher than those in JL-06. However, the ratio of chlorophyll b to chlorophyll a in the leaves of different varieties of Chinese kale remained almost identical, at approximately 0.7.
Table 2. Chlorophyll concentrations (mg g–1 DW) in the leaves of Chinese kale.
| Variety | Chlorophyll a | Chlorophyll b | Total chlorophyll |
|---|---|---|---|
| JL-01 | 7.04h | 5.64h,i | 12.68h |
| JL-02 | 10.64d | 7.28d,e | 17.92d |
| JL-03 | 12.60a | 9.40a | 22.01a |
| JL-05 | 11.63b,c | 8.05b | 19.68b,c |
| JL-06 | 3.11k | 2.14l | 5.24k |
| JL-07 | 9.28e,f | 6.21f,g,h | 15.49e,f |
| JL-08 | 11.02c,d | 7.65b,c,d | 18.67c,d |
| JL-11 | 9.62e | 6.81e,f | 16.43e |
| JL-13 | 11.04c,d | 7.93b,c | 18.97c,d |
| JL-14 | 8.39g | 5.85g,h,i | 14.23f,g |
| JL-16 | 8.15g | 5.57i | 13.72g,h |
| JL-17 | 8.93e,f,g | 6.28f,g | 15.21e,f |
| JL-18 | 8.81f,g | 6.22f,g,h | 15.03f |
| JL-19 | 12.02a,b | 8.82a | 20.84a,b |
| JL-21 | 5.02i | 3.26j,k | 8.28i |
| JL-22 | 4.13j | 2.73k,l | 6.86j |
| JL-24 | 5.09i | 3.57j | 8.66i |
| JL-25 | 10.46d | 7.41c,d,e | 17.87d |
| JL-26 | 8.62f,g | 5.92g,h,i | 14.54f,g |
Data are mean values (n = 3). Values followed by different superscript letters in the same column are significant differences for P < 0.05.
Significant differences were found in chlorophyll concentrations of bolting stems of different varieties of Chinese kale (Table 3). The distribution of chlorophyll in bolting stems was identical to those in leaves; however, the concentrations were notably lower than in leaves. Among the tested varieties, the lowest concentrations of chlorophyll a (0.53 mg g–1 DW), chlorophyll b (0.44 mg g–1 DW) and total chlorophyll (0.97 mg g–1 DW) were observed in JL-06, while the highest levels were observed in JL-03 (1.84, 1.30 and 3.14 mg g–1 DW, respectively). These were approximately three-fold higher than those observed in JL-06. Moreover, no significant difference in the ratio of chlorophyll b to chlorophyll a was found in the distinct varieties of Chinese kale, ranging from 0.67 to 0.83.
Table 3. Chlorophyll concentrations (mg g–1 DW) in the bolting stems of Chinese kale.
| Variety | Chlorophyll a | Chlorophyll b | Total chlorophyll |
|---|---|---|---|
| JL-01 | 0.83g,h | 0.64g,h,i | 1.47h,i |
| JL-02 | 1.20c,d | 0.87c,d | 2.07c |
| JL-03 | 1.84a | 1.30a | 3.14a |
| JL-05 | 1.23c | 0.85c,d | 2.08c |
| JL-06 | 0.53j | 0.44k | 0.97k |
| JL-07 | 1.09c,d,e | 0.78c,d,e,f | 1.87c,d,e,f |
| JL-08 | 1.16c,d | 0.82c,d,e | 1.98c,d,e |
| JL-11 | 1.12c,d,e | 0.82c,d,e | 1.94c,d,e |
| JL-13 | 1.16c,d | 0.88c | 2.04c,d |
| JL-14 | 0.92f,g,h | 0.69f,g,h,i | 1.61f,g,h,i |
| JL-16 | 0.98e,f,g | 0.75d,e,f,g | 1.73e,f,g,h |
| JL-17 | 0.83g,h | 0.69f,g,h,i | 1.52g,h,i |
| JL-18 | 1.06d,e,f | 0.72e,f,g,h | 1.78d,e,f,g |
| JL-19 | 1.44b | 1.02b | 2.46b |
| JL-21 | 0.78h,i | 0.58i,j | 1.36i,j |
| JL-22 | 0.79h,i | 0.60h,i,j | 1.38i,j |
| JL-24 | 0.66i,j | 0.51j,k | 1.16j,k |
| JL-25 | 1.18c,d | 0.89c | 2.06c |
| JL-26 | 0.91f,g,h | 0.69f,g,h,i | 1.59g,h,i |
Data are mean values (n = 3). Values followed by different superscript letters in the same column are significant differences for P < 0.05.
Carotenoid concentrations in leaves and bolting stems of Chinese kale
Significant differences in carotenoid concentrations were found between the leaves of different varieties of Chinese kale (Table 4). Among the tested varieties, the lowest concentrations of neoxanthin, violaxanthin, lutein, β-carotene and total carotenoids were in JL-06, which were 0.29, 0.06, 0.66, 0.21 and 1.23 mg g–1 DW, respectively. The highest concentrations of neoxanthin (1.11 mg g–1 DW), lutein (2.53 mg g–1 DW), β-carotene (0.74 mg g–1 DW) and total carotenoids (4.51 mg g–1 DW) were found in JL-03, and were more than three-fold than found in JL-06. Moreover, the highest concentration of violaxanthin (0.18 mg g–1 DW) was observed in JL-25, and was 2.8-fold higher than that in JL-06. The differentiation of carotenoid proportion in the leaves of different varieties of Chinese kale was small. Among carotenoids, lutein was predominant, accounting for more than 50% of total carotenoid concentrations, followed by neoxanthin and β-carotene, which accounted for more than 20% and 10%, respectively. The lowest proportion was that of violaxanthin, accounting for 2.7-5.8%, in different varieties.
Table 4. Carotenoid concentrations (mg g–1 DW) in the leaves of Chinese kale.
| Variety | Neoxanthin | Violaxanthin | Lutein | β-carotene | Total carotenoids |
|---|---|---|---|---|---|
| JL-01 | 0.70d | 0.07f | 1.57e | 0.36g,h | 2.70e |
| JL-02 | 0.89b | 0.16a,b | 1.95c | 0.58b,c,d | 3.58c |
| JL-03 | 1.11a | 0.13a,b,c,d | 2.53a | 0.74a | 4.51a |
| JL-05 | 0.94b | 0.15a,b,c | 2.02b,c | 0.61b,c | 3.72c |
| JL-06 | 0.29f | 0.06f | 0.66g | 0.21i | 1.23g |
| JL-07 | 0.73d | 0.17a,b | 1.71e | 0.49d,e,f | 3.09d |
| JL-08 | 0.91b | 0.14a,b,c | 1.98c | 0.61b,c | 3.64c |
| JL-11 | 0.74c,d | 0.13a,b,c,d | 1.74d,e | 0.49d,e,f | 3.10d |
| JL-13 | 0.84b,c | 0.11c,d,e,f | 1.98c | 0.57b,c,d | 3.50c |
| JL-14 | 0.75c,d | 0.12b,c,d,e | 1.69e | 0.43f,g | 2.98d,e |
| JL-16 | 0.68d | 0.14a,b,c,d | 1.62e | 0.46e,f | 2.90d,e |
| JL-17 | 0.75c,d | 0.14a,b,c | 1.68e | 0.52c,d,e | 3.09d |
| JL-18 | 0.73d | 0.16a,b | 1.64e | 0.49d,e,f | 3.03d,e |
| JL-19 | 1.06a | 0.16a,b | 2.19b | 0.65a,b | 4.06b |
| JL-21 | 0.43e | 0.09d,e,f | 0.91f | 0.29h,i | 1.71f |
| JL-22 | 0.35e,f | 0.08e,f | 0.81f,g | 0.23i | 1.47f,g |
| JL-24 | 0.45e | 0.10c,d,e,f | 0.92f | 0.30h,i | 1.78f |
| JL-25 | 0.90b | 0.18a | 1.92c,d | 0.55c,d,e | 3.54c |
| JL-26 | 0.73d | 0.12b,c,d,e | 1.65e | 0.49d,e,f | 2.99d,e |
Data are mean values (n = 3). Values followed by different superscript letters in the same column are significant differences for P < 0.05.
Significant differences were found in carotenoid concentrations of bolting stems in different varieties of Chinese kale (Table 5). Among varieties, JL-06 has the lowest concentrations of neoxanthin, lutein, β-carotene and total carotenoids, which were 0.11, 0.14, 0.04 and 0.34 mg g–1 DW, respectively. In addition, concentrations of β-carotene and total carotenoids were also relatively low in JL-24. JL-02 and JL-17 had the lowest concentration of violaxanthin, which was 0.03 mg g–1 DW for both. JL-03 had the highest concentrations of neoxanthin (0.19 mg g–1 DW), violaxanthin (0.06 mg g–1 DW), lutein (0.36 mg g–1 DW), β-carotene (0.12 mg g–1 DW) and total carotenoids (0.73 mg g–1 DW). These were, respectively, 1.8-, 1.8-, 2.6-, 3.1- and 2.2-fold of the corresponding lowest content. No significant difference was found in the proportion of individual carotenoid in different varieties of Chinese kale; lutein accounted for the highest proportion, with approximately 50% of total carotenoid concentration, followed by Neoxanthin and β-carotene accounting for approximately 20% and 10–20%, respectively. Violaxanthin had the lowest proportion, with only 6–16%.
Table 5. Carotenoid concentration (mg g–1 DW) in the bolting stems of Chinese kale.
| Variety | Neoxanthin | Violaxanthin | Lutein | β-carotene | Total carotenoids |
|---|---|---|---|---|---|
| JL-01 | 0.14b,c,d | 0.05a,b | 0.20f,g,h,i | 0.05d,e,f,g | 0.45d,e,f,g |
| JL-02 | 0.15b,c | 0.03b | 0.25c,d,e,f | 0.08b,c | 0.51c,d |
| JL-03 | 0.19a | 0.06a | 0.36a | 0.12a | 0.73a |
| JL-05 | 0.15b,c | 0.04a,b | 0.28b,c,d | 0.08c,d | 0.54b,c |
| JL-06 | 0.11d | 0.06a,b | 0.14j | 0.04g | 0.34h |
| JL-07 | 0.15b,c | 0.04a,b | 0.28b,c | 0.07c,d,e | 0.53b,c |
| JL-08 | 0.15b,c | 0.04a,b | 0.23d,e,f,g | 0.06c,d,e,f | 0.48c,d,e |
| JL-11 | 0.13b,c,d | 0.04a,b | 0.27b,c,d,e | 0.06c,d,e,f | 0.50c,d |
| JL-13 | 0.12b,c,d | 0.04a,b | 0.23c,d,e,f,g | 0.06c,d,e,f | 0.46d,e,f |
| JL-14 | 0.13b,c,d | 0.04a,b | 0.23d,e,f,g | 0.05e,f,g | 0.45d,e,f,g |
| JL-16 | 0.12b,c,d | 0.05a,b | 0.22e,f,g,h | 0.06c,d,e,f | 0.45d,e,f,g |
| JL-17 | 0.12c,d | 0.03b | 0.19g,h,i | 0.06d,e,f,g | 0.40f,g,h |
| JL-18 | 0.15b | 0.03a,b | 0.23d,e,f,g | 0.07c,d,e | 0.49c,d,e |
| JL-19 | 0.15b | 0.05a,b | 0.30b | 0.10a,b | 0.59b |
| JL-21 | 0.13b,c,d | 0.04a,b | 0.17i,j | 0.06d,e,f,g | 0.40f,g,h |
| JL-22 | 0.12b,c,d | 0.04a,b | 0.19h,i | 0.04f,g | 0.39g,h |
| JL-24 | 0.12b,c,d | 0.03a,b | 0.16i,j | 0.04g | 0.35h |
| JL-25 | 0.14b,c,d | 0.05a,b | 0.26b,c,d,e | 0.06d,e,f,g | 0.51c,d |
| JL-26 | 0.13b,c,d | 0.04a,b | 0.20g,h,i | 0.06d,e,f,g | 0.43e,f,g |
Data are mean values (n = 3). Values followed by different superscript letters in the same column are significant differences for P < 0.05.
PCA and PLS-DA analysis
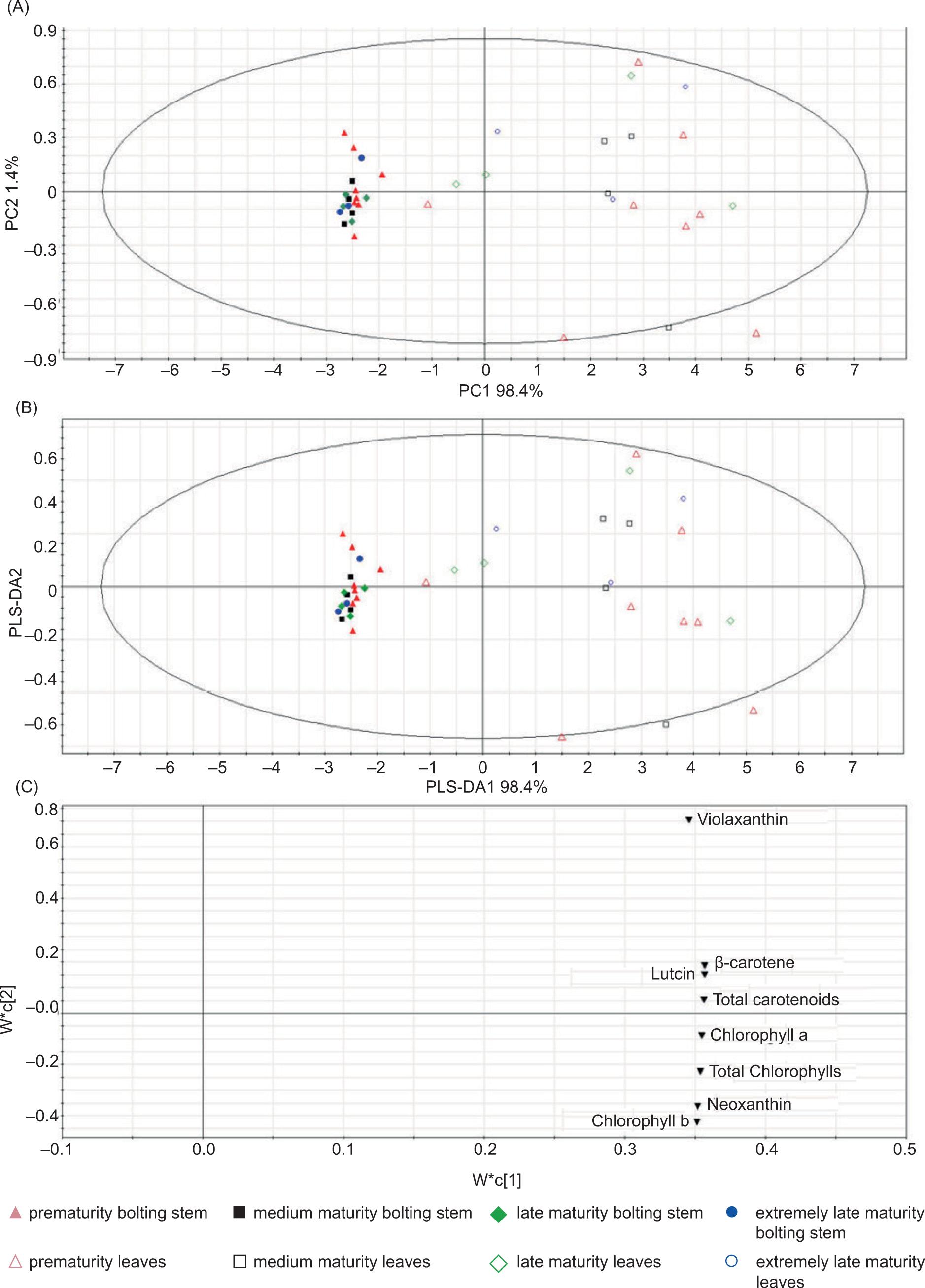
Principal component analysis of chlorophyll and carotenoids was performed to investigate directly variation in different organs, varieties and maturities of Chinese kale. As shown in Figure 2A, two principal components were obtained through dimension reduction. Principal component 1 (PC1) played a major role (accounting for 98.4%), while principal component 2 (PC2) played a minor role (accounting for only 1.4%). The sum of both PCs was nearly 100%. Leaves and bolting stems were discriminated from each other by PC1. PC1 and PC2 were located on the positive and negative sides of the abscissa axis, respectively, and distribution of the bolting stem was relatively compact. However, different varieties of Chinese kale could not be separated into different groups as organs or maturities by PC2, which had little effect, because of its low proportion. The results of PLS-DA analysis were consistent with that of PCA. Bolting stems were clearly separated from leaves by PLS-DA1, while different varieties of Chinese kale were not separated into different groups according to organs or maturities by PLS-DA2 (Figure 2B). Therefore, it was concluded that organs formed the main source of differences for chlorophyll and carotenoid concentrations in different varieties of Chinese kale, while maturities and flower colors exerted little effect.
Figure 2. PCA and PLS-DA analysis of leaves and bolting stems of Chinese kale. (A) PCA score plot; (B) PLS-DA score plot; (C) PLS-DA loading plot.
The results of PLS-DA loading plot showed that each index was located at the positive position of abscissa (Figure 2C), and leaves of Chinese kale were located at the positive position of abscissa in PLS-DA score plot (Figure 2B). This suggests that leaves of Chinese kale had relatively high chlorophyll and carotenoid concentrations.
HCA analysis
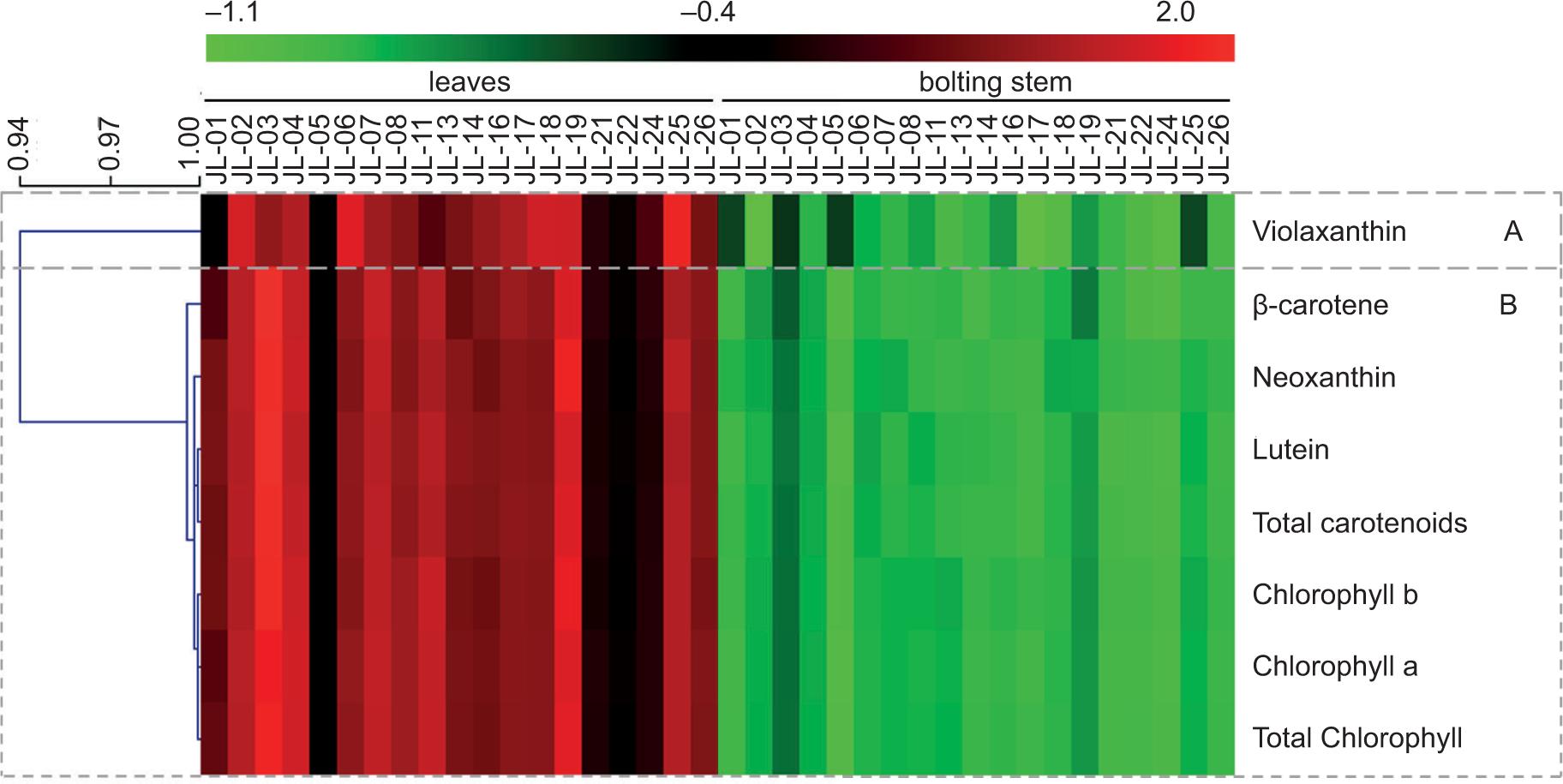
Hierarchical clustering analysis was performed to visualize chlorophyll and carotenoid concentrations in leaves and bolting stems for different varieties of Chinese kale. The concentration of each pigment was normalized, and the results are shown in Figure 3. The red color in Figure 3 represents relatively high concentrations, and the green color represents relatively low concentrations. The concentrations of chlorophyll and carotenoids in leaves of different varieties of Chinese kale were significantly higher than those in bolting stems. The pigments were divided into two parts. Group A contains violaxanthin, which was highest in the leaves of JL-25 and JL-07, and lowest in the bolting stems of JL-02 and JL-17. Group B contains β-carotene, neoxanthin, lutein, total carotenoids, chlorophyll b, chlorophyll a and total chlorophyll. These were higher in the leaves of JL-03 and JL-19, and lower in the bolting stems of JL-06.
Figure 3. Heatmap and HCA of chlorophyll and carotenoids in Chinese kale. Red and green colors reflect relative concentrations.
Correlation analysis
Correlation analysis was conducted to investigate correlations between chlorophyll and carotenoids in leaves and bolting stems of different varieties of Chinese kale (Table 6). A significantly positive correlation (P < 0.01) was observed between chlorophyll and carotenoids. All correlation coefficients were above 0.9, and the correlation coefficient between chlorophyll a and total chlorophyll was highest (1.000), while the correlation was relatively low between violaxanthin and others. Moreover, the lowest correlation coefficient was found between violaxanthin and total carotenoids (0.939), indicating that the contribution of violaxanthin to total carotenoids was relatively small.
Table 6. Correlation coefficients of chlorophylls and carotenoids in leaves and bolting stems of Chinese kale.
| Item | Chlorophyll a | Chlorophyll b | Total chlorophyll | Neoxanthin | Violaxanthin | Lutein | β-carotene |
|---|---|---|---|---|---|---|---|
| Chlorophyll b | 0.998 | – | – | – | – | – | – |
| Total chlorophyll | 1.000 | 0.999 | – | – | – | – | – |
| Neoxanthin | 0.996 | 0.997 | 0.997 | – | – | – | – |
| Violaxanthin | 0.942 | 0.930 | 0.937 | 0.933 | – | – | – |
| Lutein | 0.996 | 0.997 | 0.997 | 0.997 | 0.932 | – | – |
| β-carotene | 0.997 | 0.995 | 0.996 | 0.994 | 0.941 | 0.995 | – |
| Total carotenoids | 0.998 | 0.998 | 0.998 | 0.998 | 0.939 | 0.999 | 0.997 |
Discussion
Carotenoids have strong antioxidant capacity and play an important role in human health. Chinese kale is a good source of carotenoids and contains high concentrations of carotenoids. The results showed that the highest concentrations of neoxanthin, violaxanthin, lutein, β-carotene and total carotenoids in leaves of Chinese kale were 1.11, 0.18, 2.53, 0.74 and 4.51 mg g–1 DW, respectively. The highest concentrations of neoxanthin, violaxanthin, lutein, β-carotene and total carotenoids in bolting stems were 0.19, 0.06, 0.36, 0.12 and 0.73 mg g–1 DW, respectively. Individual and total carotenoid concentrations in Chinese kale were significantly higher than those of mature pepper (Lee et al., 2005). Lutein was identified as the predominant carotenoid, accounting for approximately 50% of total carotenoids, and this result was consistent with the results of Farnham and Kopsell (2009) for broccoli. In addition, JL-03 could be used to breed a functional variety because of its highest concentration of chlorophyll and carotenoids (except violaxanthin in leaves).
Numerous studies showed that significant differences in the composition and concentrations of phytochemicals exist in different varieties or organs of vegetables (Abu-Reidah et al., 2017; Agneta et al., 2014; Amri et al., 2017). This could be due to the genetic diversity of most characters in different varieties and organs (Hanson et al., 2011). In this study, the concentrations of chlorophyll and carotenoids in leaves were significantly higher than those in bolting stems. This result was consistent with previous studies in baby mustard (Brassica juncea var. gemmifera) (Sun et al., 2018) and bitter melon (Momordica charantia) (Tuan et al., 2011). Perhaps, it is because chlorophyll and carotenoids are important photosynthetic pigments in plants, and photosynthesis mainly occurs in leaves (Tuan et al., 2011).
Many studies have indicated that plant species with different colors vary greatly in their carotenoid concentrations (Carvalho et al., 2013). Carotenoid concentrations in Brassica napus with yellow flowers were significantly higher than those with white flowers, and the predominant carotenoid was violaxanthin (Zhang et al., 2015). However, this study did not find significant differences in leaves or bolting stems of Chinese kale with different flower colors. It is possible that carotenoids have different metabolism in flowers with different colors, but the metabolism in other organs was not affected by that in petals (Perrin et al., 2017). This finding implied that flower color exerted a limited effect on chlorophyll and carotenoid concentrations of leaves and bolting stems of Chinese kale; however, it is necessary to analyze whether it affects other phytochemicals as well.
In recent years, many studies have investigated plant maturity, and mainly focused on genetic analysis (Li et al., 2019; Nuñez-Lillo et al., 2019) and cultivation techniques (Lu et al., 2017). The present study, for the first time, has measured chlorophyll and carotenoid concentrations of Chinese kale with different maturities. However, no significant correlations were found between pigment concentrations and maturities, indicating that maturity exerted a limited effect on chlorophyll and carotenoid concentrations of Chinese kale. It could be that the sowing period of Chinese kale in this experiment was not strictly separated, since the maturity of Chinese kale differed in response to the most suitable sowing period. The possible reason requires further study.
Many studies have reported a significant positive correlation between chlorophyll and carotenoids, and the correlation coefficient was highest compared to other indicators (Sun et al., 2018; Yousefi et al., 2019). The correlation analysis in the present experiment showed significant positive correlations between chlorophyll and carotenoids (P < 0.01), which was consistent with results of previous studies (Conesa et al., 2019; Sun et al., 2018).
Conclusion
This study reported the composition and concentrations of chlorophyll and carotenoids in different organs, varieties, maturities and flower colors of Chinese kale. The pigment concentrations were significantly higher in leaves than in bolting stems, indicating that organs exerted a major role in chlorophyll and carotenoid accumulation. This was followed by variety effect, while maturity and flower color exerted little effect. Lutein was the main component of carotenoids and accounted for approximately 50% of total carotenoids. JL-03 had the highest concentrations of chlorophyll and carotenoids, and was thus a promising variety for the future breeding initiatives.
Acknowledgements
This work was supported by National Natural Science Foundation of China (32072586, 31500247), Natural Science Foundation of Sichuan Province (2022NSFSC1689), and Project of New Varieties Breeding of Sichuan Vegetable Innovation Team (sccxtd-2022-05).
REFERENCES
Abu-Reidah, I.M., Gil-Izquierdo, Á., Medina, S. and Ferreres, F., 2017. Phenolic composition profiling of different edible parts and by-products of date palm (phoenix dactylifera L.) by using HPLC-DAD-ESI/MSn. Food Research International 100: 494–500. 10.1016/j.foodres.2016.10.018
Agneta, R., Lelario, F., De Maria, S., Möllers, C., Bufo, S.A. and Rivelli, A.R., 2014. Glucosinolate profile and distribution among plant tissues and phenological stages of field-grown horseradish. Phytochemistry 106: 178–187. 10.1016/j.phytochem.2014.06.019
Amin, A.R., Kucuk, O., Kuhari, F.R. and Shin, D.M., 2009. Perspectives for cancer prevention with natural compounds. Journal of Clinical Oncology 27: 2712–2725. /10.1200/JCO.2008.20.6235
Amri, Z., Zaouay, F., Lazreg-Aref, H., Soltana, H., Mneri, A., Mars, M. and Hammami, M., 2017. Phytochemical content, fatty acids composition and antioxidant potential of different pomegranate parts: comparison between edible and nonedible varieties grown in Tunisia. International Journal of Biological Macromolecules 104: 274–280. 10.1016/j.ijbiomac.2017.06.022
Britton, G. and Khachik, F., 2009. Carotenoids in food. Carotenoids 5: 45–66. 10.1007/978-3-7643-7501-0_3
Carvalho, E., Fraser, P.D. and Martens, S., 2013. Carotenoids and tocopherols in yellow and red raspberries. Food Chemistry 139: 744–752. 10.1016/j.foodchem.2012.12.047
Caseli, L., Sousa-Martins, D., Maia, M., Lima-Filho, A.A., Rodrigues, E.B. and Belfort, R. Jr., 2013. An intraocular dye solution based on Lutein and Zeaxanthin in a surrogate internal limiting membrane model: a Langmuir monolayer study. Colloids and Surfaces B: Biointerfaces 107: 124–129. 10.1016/j.colsurfb.2013.01.076
Cazzonelli, C.I. and Pogson, B.J., 2010. Source to sink: regulation of carotenoid biosynthesis in plants. Trends in Plant Science 15: 266–274. 10.1016/j.tplants.2010.02.003
Cervantes-Paz, B., Yahia, E.M., Ornelas-Paz, J.D.J., Victoria-Campos, C.I., Ibarra-Junquera, V., Pérez-Martínez, J.D., et al. 2014. Antioxidant activity and content of chlorophylls and carotenoids in raw and heat-processed Jalapeno peppers at intermediate stages of ripening. Food Chemistry 146: 188–196. 10.1016/j.foodchem.2013.09.060
Chen, K.W. and Roca, M., 2018. In vitro bioavailability of chlorophyll pigments from edible seaweeds. Journal of Functional Foods 41: 25–33. 10.1016/j.jff.2017.12.029
Chen, Y.S., Wang, E.P., Wei, Z.H., Zheng, Y.F., Yan, R. and Ma, X., 2019. Phytochemical analysis, cellular antioxidant and α-glucosidase inhibitory activities of various herb plant organs. Industrial Crops and Product 141: 111771. 10.1016/j.indcrop.2019.111771
Conesa, A., Manera, F.C., Brotons, J.M. and Fernandez-Zapata, J.C., 2019. Changes in the content of chlorophylls and carotenoids in the rind of Fino 49 lemons during maturation and their relationship with parameters from the CIELAB color space. Scientia Horticulturae 243: 252–260. 10.1016/j.scienta.2018.08.030
Farnham, M.W. and Kopsell, D.A., 2009. Importance of genotype on carotenoid and chlorophyll levels in broccoli heads. Horticultue Science 44: 1248–1253. 10.21273/HORTSCI.44.5.1248
Ferruzzi, M.G. and Blakeslee, J., 2007. Digestion, absorption, and cancer preventative activity of dietary chlorophyll derivatives. Nutrition Research 27: 1–12. 10.1016/j.nutres.2006.12.003
Hanson, P., Yang, R.Y., Chang, L.C., Ledesma, L. and Ledesma, D., 2011. Carotenoids, ascorbic acid, minerals, and total glucosinolates in choysum (Brassica rapa cvg. parachinensis) and kailaan (B. oleraceae Alboglabra group) as affected by variety and wet and dry season production. Journal of Food Composition and Analysis 24: 950–962. 10.1016/j.jfca.2011.02.001
Jubert, C., Mata, J., Bench, G., Dashwood, R., Pereira, C., Tracewell, W., et al. 2009. Effects of chlorophyll and chlorophyllin on low-dose aflatoxin b(1) pharmacokinetics in human volunteers. Cancer Prevention Research 2: 1015–1022. 10.1158/1940-6207.CAPR-09-0099
Lee, J.J., Crosby, K.M., Pike, L.M., Yoo, K.S. and Leskovar, D.I., 2005. Impact of genetic and environmental variation on development of flavonoids and carotenoids in pepper (capsicum spp.). Scientia Horticulturae 106: 341–352. 10.1016/j.scienta.2005.04.008
Lei, J.J., Chen, G.J., Chen, C.M. and Cao, B.H., 2017. Germplasm diversity of Chinese kale in China. Horticultural Plant Journal 3: 101–104. 10.1016/j.hpj.2017.07.006
Li, J.W., Wang, Y.H. and Wen, G.H., 2019. Mapping QTL underlying tuber starch content and plant maturity in tetraploid potato. Crop Journal 7: 261–272. 10.1016/j.cj.2018.12.003
Lu, H.D., Xue, J.Q. and Guo, D.W., 2017. Efficacy of planting date adjustment as a cultivation strategy to cope with drought stress and increase rainfed maize yield and water-use efficiency. Agricultural Water Management 179: 227–235. 10.1016/j.agwat.2016.09.001
Nuñez-Lillo, G., Balladares, C., Pavez, C., Urra, C., Sanhueza, D., Vendramin, E., et al. 2019. High-density genetic map and QTL analysis of soluble solid content, maturity date, and mealiness in peach using genotyping by sequencing. Scientia Horticulturae 257: 108734. 10.1016/j.scienta.2019.108734
Perrin, F., Hartmann, L., Dubois-Laurent, C., Welsch, R., Huet, S., Hamama, L., et al. 2017. Carotenoid gene expression explains the difference of carotenoid accumulation in carrot root tissues. Planta 245: 737–747. 10.1021/jf4004576
Quinlan, R.F., Shumskaya, M., Bradbury, L.M., Beltrán, J., Ma, C., Kennelly, E.J., et al. 2012. Synergistic interactions between carotene ring hydroxylases drive lutein formation in plant carotenoid biosynthesis. Plant Physiology 160: 204–214. 10.1104/pp.112.198556
Rodriguez-Concepcion, M., Avalos, J., Bonet, M.L., Boronat, A., Gomez-Gomez, L., Hornero-Mendez, et al. 2018. A global perspective on carotenoids: metabolism, biotechnology, and benefits for nutrition and health. Progress in Lipid Research 70: 62–93. 10.1016/j.plipres.2018.04.004
Shi, Y.M., Wang, R., Luo, Z.P., Jin, L.F., Liu, P.P., Chen, Q.S., et al. 2014. Molecular cloning and functional characterization of the lycopene ε-cyclase gene via virus-induced gene silencing and its expression pattern in Nicotine abacus. International Journal of Molecular Science 15: 14766–14785. 10.3390/ijms150814766
Sun, B., Liu, N., Zhao, Y.T., Yan, H.Z. and Wang, Q.M., 2011. Variation of glucosinolates in three edible parts of Chinese kale (Brassica alboglabra Bailey) varieties. Food Chemistry 124: 941–947. 10.1016/j.foodchem.2010.07.031
Sun, B., Tian, Y.X., Jiang, M., Yuan, Q., Chen, Q., Zhang, Y., et al. 2018. Variation in the main health-promoting compounds and antioxidant activity of whole and individual edible parts of baby mustard (Brassica juncea var. gemmifera). RSD Advances 8: 33845–33854. 10.1039/C8RA05504A
Sun, B., Yan, H.Z., Liu, N., Wei, J. and Wang, Q.M., 2012a. Effect of 1-MCPC treatment on postharvest quality characters, antioxidants and glucosinolates of Chinese kale. Food Chemistry 131: 519–526. 10.1016/j.foodchem.2011.09.016
Sun, B., Yan, H.Z., Zhang, F. and Wang, Q.M., 2012b. Effects of plant hormones on main health-promoting compounds and antioxidant capacity of Chinese kale. Food Research International 48: 359–366. 10.1016/j.foodres.2012.04.021
Sun, B., Yuan, Q., Zheng, H., Liang, S., Jiang, M., Wang, M.M., et al. 2019. An efficient and economical protocol for isolating, purifying, and PEG-mediated transient gene expression of Chinese kale hypocotyl protoplasts. Plants 8: 385. 10.3390/plants8100385
Takaidza, S., Mtunzi, F. and Pillay, M., 2018. Analysis of the phytochemical contents and antioxidant activities of crude extracts from Tulbaghia species. Journal of Traditional Chinese Medicine 38: 272–279. 10.1016/j.jtcm.2018.04.005
Tuan, P.A., Kim, J.K., Park, N.I., Lee, S.Y. and Park, S.U., 2011. Carotenoid content and expression of phytoene synthase and phytoene desaturase genes in bitter melon (Momordica charantia). Food Chemistry 126: 1686–1692. 10.1016/j.foodchem.2010.12.058
Yousefi, F., Jabbarzadeh, Z., Amiri, J. and Rasouli-Sadaghiani, M.H., 2019. Response of roses (Rosa hybrida L. ‘Herbert Stevens’) to foliar application of polyamines on root development, flowering, photosynthetic pigments, antioxidant enzymes activity and NPK. Scientific Reports 9: 1–11. 10.1038/s41598-019-52547-1
Zhang, B., Liu, C., Wang, Y.Q., Yao, X., Wang, F., Wu, J.S., et al. 2015. Disruption of a carotenoid cleavage dioxygenase 4 gene converts flower colour from white to yellow in brassica species. New Phytologist 206: 1513–1526. 10.1111/nph.13335
Zhang, Z.H., Peng, H.D., Ma, H.L. and Zeng, X.A., 2019. Effect of inlet air drying temperatures on the physicochemical properties and antioxidant activity of whey protein isolate-kale leaves chlorophyll (WPI-CH) microcapsules. Journal of Food Engineering 245: 149–156. 10.1016/j.jfoodeng.2018.10.011