
Download




Original Article
Crocin ameliorates peritoneal fibrosis in rat induced by peritoneal dialysis via Wnt5a/β-Catenin pathway
Jingyuan Lu1, 2#, Danye Shi3#, Xinhui Zhao2, Minhui Xi2, Hualin Qi2, Qiang He1, 4*
1Medical College, Soochow University, Suzhou, China;
2Department of Nephrology, Pudong New Area People’s Hospital, Shanghai, China;
3Department of Nephrology, Minhang Hospital, Fudan University, Shanghai, China;
4Department of Nephrology, the First Affiliated Hospital of Zhejiang Chinese Medical University (Zhejiang Provincial Hospital of Traditional Chinese Medicine), Hangzhou, China
Abstract
Peritoneal dialysis is used in the treatment of patients with kidney diseases. Long-term peritoneal dialysis could result in peritoneal fibrosis and recurrent peritonitis, thus leading to failure of ultrafiltration. Crocin is a bioactive carotenoid and isolated from stigma of Crocus sativus, and ameliorates pulmonary and myocardial fibrosis. The role of crocin in peritoneal fibrosis was assessed. Firstly, rats model with peritoneal dialysis was treated with 4.25% peritoneal dialysate. Results showed that injection with peritoneal dialysate induced obvious hyperplasia and increased thickness in peritoneum structure. Rats with peritoneal dialysis were injected with increasing concentrations of crocin at 10, 20, or 40 mg/kg. Crocin ameliorated the pathological changes in the peritoneum of peritoneal dialysate-induced rats. Secondly, crocin attenuated peritoneal dialysate-induced decrease of E-cadherin, increase of fibronectin, α-smooth muscle actin (α-SMA), and collagen I. Moreover, crocin enhanced ultrafiltration volume and reduced glucose transport in rats model with peritoneal dialysis. Thirdly, crocin also reduced levels of Interleukin (IL)-1β, Tumor Necrosis Factor-α (TNF –α), and IL-6 in peritoneal tissues of rats model with peritoneal dialysis. Lastly, protein expression of Wnt5a and β-Catenin in rats model with peritoneal dialysis were also downregulated by crocin. In conclusion, crocin exerted anti-inflammatory and anti-fibrotic effects on rats model with peritoneal dialysis through inactivation of Wnt5a/β-Catenin pathway.
Key words: crocin, peritoneal dialysis, peritoneal fibrosis, inflammation, Wnt5a/β-Catenin
Corresponding Author: Qiang He, Department of Nephrology, the First Affiliated Hospital of Zhejiang Chinese Medical University (Zhejiang Provincial Hospital of Traditional Chinese Medicine), No. 54, Youdian Road, Shangcheng District, Hangzhou City, Zhejiang Province, China. Email: [email protected]
#These authors contributed equally to the work and should be considered co-first authors.
Received: 21 July 2022; Accepted: 4 August 2022; Published: 1 September 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Prevalence and incidence of chronic kidney diseases, especially end-stage renal disease, has increased recently (Tsuzuki et al., 2018). End-stage renal disease, accompanied by cystic diseases, is a major risk of renal cell carcinoma (Tsuzuki et al., 2018). Peritoneal dialysis is widely used as an effective renal replacement therapy to prolong survival and improve quality of life in patients with end-stage renal disease (Khan and Rosner, 2018). However, continuous peritoneal dialysis induces contact between peritoneum and peritoneal dialysis fluid containing low pH, glucose degradation products, and high concentrations of glucose and lactate, results in peritoneal fibrosis, and finally leads to failure of ultrafiltration and withdrawal of peritoneal dialysis (Liu et al., 2020). Moreover, some patients post peritoneal dialysis would develop encapsulating peritoneal sclerosis with high mortality (Korte et al., 2011). Therefore, effective therapies to inhibit process of peritoneal fibrosis are urgently needed for the treatment of end-stage renal disease.
Emerging evidence has shown that high concentrations of glucose during peritoneal dialysis induces expression of transforming growth factor-β1 (TGF-β1), and TGF-β1 functions as a profibrotic cytokine to stimulate peritoneal epithelial-mesenchymal transition and triggers peritoneal fibrosis (Liu et al., 2008). TGF-β1 is an indispensable factor to maintain homeostasis of the immune system, and blocking of the TGF-β1 pathway induces long-term problems (Heo et al., 2021). Epithelial-mesenchymal transformation with the transdifferentiation of epithelial cells into mesenchymal cells is an initial reversible step in the development of peritoneal fibrosis (Kang, 2020). Thus, targeting epithelial-mesenchymal transition is regarded as a better approach for the treatment of peritoneal fibrosis (Kang, 2020).
Crocin is a bioactive carotenoid and isolated from stigma of Crocus sativus, and exerts anti-oxidant and anti- inflammatory properties on bone and cartilage diseases (Vafaei et al., 2022). Crocin also exhibited protective effects against lipid and glucose metabolic dysfunction through amelioration of insulin sensitivity (Fang and Gu, 2020). In addition, crocin isreduced levels of fibrogenic factor (TGF-β and α-SMA) in carbon tetrachloride- induced rats to inhibit liver fibrosis (Chhimwal et al., 2020), reduced pulmonary inflammation and vascular dysfunction to attenuate pulmonary fibrosis in bleomycin- induced rats (Zaghloul et al., 2019). Isoprenaline-induced up-regulation of connective tissue growth factor and TGF-βwas also ameliorated by crocin, thus suppressing myocardial fibrosis (Jin et al., 2020). Epithelial-mesenchymal transition of cancer cells were repressed by crocin (Wu and Hui, 2020; Zhou et al., 2019). However, the role of crocin in peritoneal fibrosis remains unknown.
In this study, the effects of crocin on inflammation and fibrosis of rats model with peritoneal fibrosis are studied. The results might provide potential agent for the treatment of end-stage renal disease in combination with peritoneal dialysis.
Materials and Methods
Animal models
Fifty specific pathogen-free Sprague Dawley rats (weighing 200–250 g and 8–10 weeks old) were acquired from Shanghai Slack Laboratory Animal Co., Ltd (Shanghai, China). Rats were divided into five groups: blank (n = 10), peritoneal dialysis group (PD, n = 10), PD with 10 mg/kg crocin (n = 10), PD with 20 mg/kg crocin (n = 10), and PD with 40 mg/kg crocin (n = 10). Rats in the blank group were intraperitoneally injected with 10 mL phosphate- buffered saline buffer solution daily for 8 weeks. Rats were then intraperitoneally injected with 4.25% dextrose peritoneal dialysis solution (5B9866; Baxter, Deerfield, Illinois) daily for 8 weeks. Rats in the PD with crocin groups were also injected with different concentrations of crocin (PHL80391; Sigma-Aldrich, St. Louis, MO, USA) daily for 8 weeks. The experiment was approved by the Medical Ethics Committee of Pudong New Area People’s Hospital and in accordance with the National Institutes of Health Laboratory Animal Care and Use Guidelines.
Histopathological analysis
After the end of dosing cycle, rats in each group (n = 5) were sacrificed, peritoneal tissues were isolated and fixed in 10% formalin. The tissues were then embedded in paraffin and sliced into 5 μm sections. Sections were deparaffinized in toluene and hydrated in gradient concentrations of alcohol. Sections were then stained with hematoxylin (H3136) and eosin (E4009) (Sigma-Aldrich) or Masson’s trichrome (HT15; Sigma-Aldrich). The sections were observed under microscope (Olympus, Tokyo, Japan).
Immunofluorescence
The peritoneal sections were probed with Cy3-conjugated rabbit anti-rat E-cadherin (ab181296), anti-rat CD31 (ab222783), and anti-rat α-SMA (ab5694; Abcam, Cambridge, MA, USA) overnight. Sections were then incubated with goat anti-rabbit fluorescein isothiocyanate-conjugated immunoglobulin G (ab6717; Abcam), and the nuclei were counterstained with DAPI. Sections were observed under Zeiss LSM 510 Confocal Imaging System (Zeiss, Jena, Germany).
Peritoneal function test
Rats in each group (n = 5) were intraperitoneally injected with 4.25% dianeal peritoneal dialysate at the end of dosing cycle. Four hours later, rats were anesthetized, the peritoneum were opened, and the peritoneal fluid was taken out for determination of ultrafiltration volume (ultrafiltration volume = amount of peritoneal fluid removed 4 hours later minus amount of fluid given). For glucose transport, glucose concentration of tail vein blood in each group at 0 and 4 h were measured by automatic biochemical analyzer. Mass transfer of glucose (mmol/kg) was calculated as: (glucose concentration at 0 h × initial volume of injected dialysate) – (glucose concentration at 4 h × final volume of reserved dialysate).
Quantitative reverse transcription PCR (RT-qPCR)
Peritoneal tissues were lysed in Trizol (15596026; Invitrogen, Carlsbad, CA, USA) to isolate total RNAs. The RNAs were reverse-transcribed into cDNAs, and cDNAs were subjected to qRT-PCR analysis with SYBR Green Master (04707516001; Roche, Mannheim, Germany). GAPDH (Forward: 5’-GGATTTGGTCGTATTGGG-3’ and Reverse: 5’-GGAAGATGGTGATGGGATT-3’) was used as endogenous control. Expression of IL-1β (Forward: 5’-CACCTT CTTTTCCTTCATCTTTG-3’ and Reverse: 5’-GTCGTT GCTTGTCTCTCCTTGTA-3’), TNF-α (Forward: 5’-ACTG AACTTCGGGGTGATTG-3’ and Reverse: 5’-GCTTGGT GGTTTGCTACGAC-3’), and IL-6 (Forward: 5’-TG ATGG ATGCTTCCAAACTG-3’ and Reverse: 5’-GAGCA TTGG AAGTTGGGGTA-3’) were determined by 2 -△△CT method.
ELISA
Peritoneal tissues were lysed in radio-immunoprecipitation assay buffer (P0045; Beyotime, Beijing, China) to isolate tissue lysates. Levels of IL-1β (EK301B/2), TNF-α (70-EK382), and IL-6 (70-EK306) were determined by commercial ELISA kits (MultiSciences Biotech Co., Ltd., Wuhan, China).
Western blot
Tissue lysates were segregated using SDS-PAGE, and then transferred onto nitrocellulose membranes. Membranes were blocked in 5% dry milk, and then incubated with primary antibodies: anti-fibronectin (ab268020) and anti-GAPDH (ab181602) (1:2000), anti-α-SMA (ab124964) and anti-collagen I (ab270993) (1:2500), anti-E-cadherin (ab76319) and anti-Wnt5a (ab227229) (1:3000), anti- β-Catenin (ab32572) and anti-Axin (ab109307) (1:3500), anti-COX-2 (ab179800) (1:4000). The membranes were then incubated with secondary antibodies (ab6721) (1:5000), and subjected to chemiluminescence reagent kit (Beyotime). All the proteins were purchased from Abcam.
Statistical analysis
All the data were expressed as mean ± SEM and analyzed by student’s t test or one-way analysis of variance under SPSS 19.0 software (SPSS, Chicago, IL, USA). P < 0.05 was considered as statistically significant.
Results
Crocin alleviated peritoneal fibrosis in peritoneal dialysate-induced rats
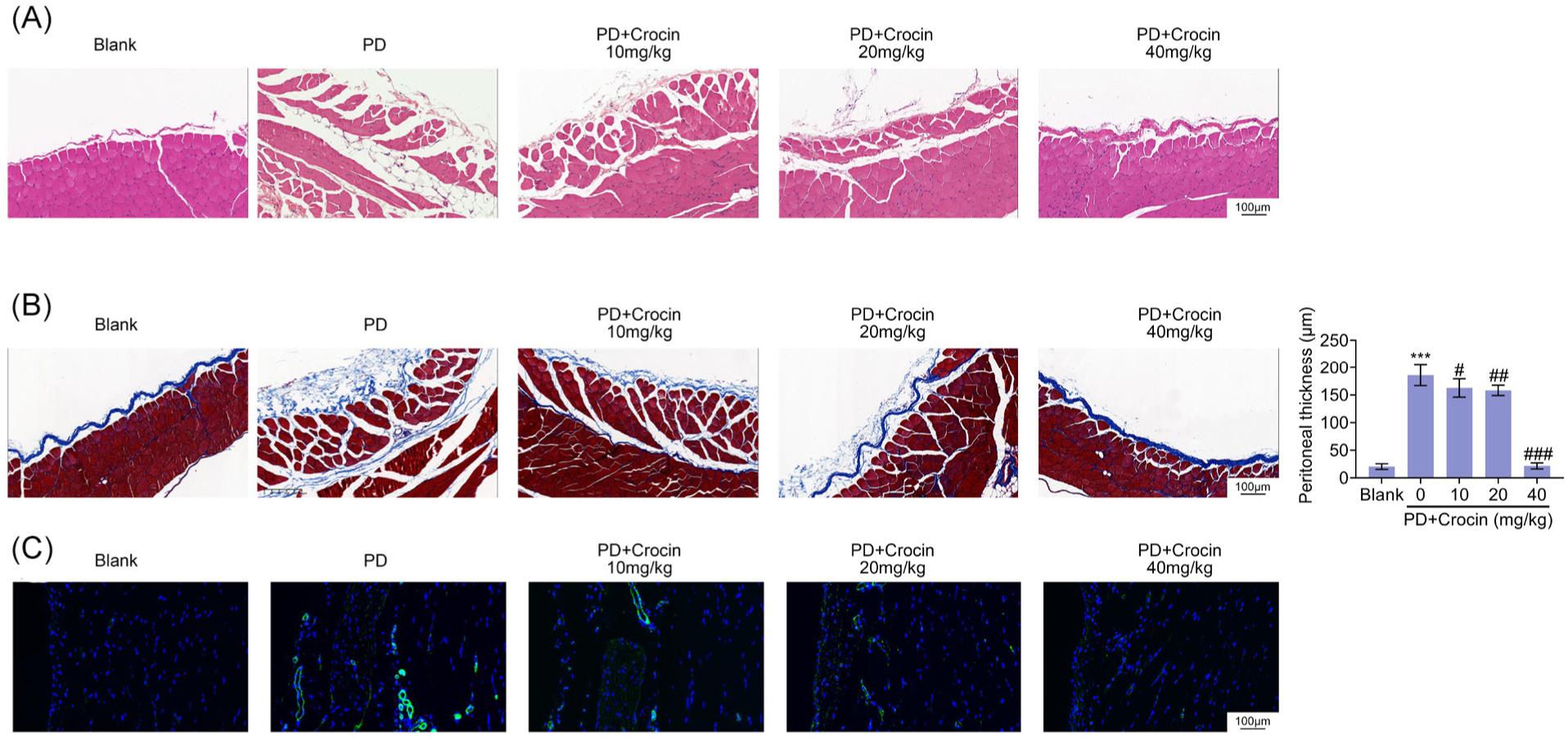
To induce peritoneal fibrosis, rats were injected with peritoneal dialysis solution. Peritoneal tissues in peritoneal dialysate-induced rats showed disordered and thickened peritoneum structure (Figure 1A). Obvious hyperplasia and vascular proliferation were also observed in peritoneal dialysate-induced rats (Figure 1A). Injection with crocin attenuated the histopathological changes in peritoneum of peritoneal dialysate-induced rats (Figure 1A). Moreover, Masson’s staining demonstrated fibrosis in peritoneum of rats post injection with peritoneal dialysis solution (Figure 1B). However, crocin also attenuated peritoneal fibrosis in peritoneal dialysate-induced rats (Figure 1B). Up-regulation of vascular marker, CD31, was also identified in peritoneal tissues of peritoneal dialysate-induced rats (Figure 1C). Crocin reduced the level of CD31 (Figure 1C).
Figure 1. Crocin alleviated peritoneal fibrosis in peritoneal dialysate-induced rats. (A) Peritoneal dialysate induced disordered and thickened peritoneum structure, as well as obviously hyperplasia and vascular proliferation, in peritoneal tissues. Injection with crocin attenuated the histopathological changes in peritoneum of peritoneal dialysate-induced rats. (B) Peritoneal dialysate induced fibrosis in peritoneum of rats, while crocin attenuated peritoneal fibrosis in peritoneal dialysate-induced rats. (C) Crocin reduced level of CD31 in peritoneal tissues of peritoneal dialysate-induced rats. *** vs. blank, Pp < 0.001. #, ##, ### vs. PD, Pp < 0.05, Pp < 0.01, Pp < 0.001.
Crocin alleviated epithelial-mesenchymal transformation in peritoneal dialysate-induced rats
Protein expression of E-cadherin was reduced in peri-toneal tissues of peritoneal dialysate-induced rats (Figure 2A). Moreover, peritoneal dialysis solution stimulated upregulation of fibronectin, α-SMA, and collagen I (Figure 2A). Immunofluorescence of α-SMA was enhanced while E-cadherin was reduced in peritoneal dialysate-induced rats (Figure 2B). Crocin increased protein expression of E-cadherin, and decreased fibronectin, α-SMA, and collagen I in peritoneal tissues of peritoneal dialysate-induced rats (Figure 2B). Moreover, crocin enhanced the immunofluorescence of E-cadherin, while it reduced α-SMA to inhibit epithelial-mesenchymal transformation (Figure 2B).
Figure 2. Crocin alleviated epithelial-mesenchymal transformation in peritoneal dialysate-induced rats. (A) Crocin increased protein expression of E-cadherin, decreased fibronectin, α-SMA and collagen I in peritoneal tissues of peritoneal dialysate-induced rats. (B) Crocin enhanced the immunofluorescence of E-cadherin while reduced α-SMA in peritoneal tissues of peritoneal dialysate-induced rats. *** vs. blank, P < 0.001. ### vs. PD, P < 0.001.
Crocin alleviated peritoneal transport function in peritoneal dialysate-induced rats
Injection with peritoneal dialysate solution downregulated ultrafiltration volume (Figure 3A) and upregulated glucose transport (Figure 3B) in rats model with peritoneal dialysis. Crocin ameliorated peritoneal transport function through upregulation of ultrafiltration volume (Figure 3A) and downregulation of glucose transport (Figure 3B).
Figure 3. Crocin alleviated peritoneal transport function in peritoneal dialysate-induced rats. (A) Crocin increased ultrafiltration volume in peritoneal dialysate-induced rats. (B) Crocin decreased glucose transport in peritoneal dialysate-induced rats. *** vs. blank, P < 0.001. #, ### vs. PD, P < 0.05, P < 0.001.
Crocin alleviated peritoneal inflammation in peritoneal dialysate-induced rats
Levels of IL-1β, TNF-α, and IL-6 were increased in peritoneal tissues of peritoneal dialysate-induced rats (Figure 4A and B). However, crocin reduced IL-1β, TNF-α, and IL-6 in peritoneal dialysate-induced rats (Figure 4A and B) to inhibit inflammatory response.
Figure 4. Crocin alleviated peritoneal inflammation in peritoneal dialysate-induced rats. (A) Crocin reduced mRNA expression of IL-1β, TNF-α and IL-6 in peritoneal dialysate-induced rats. (B) Crocin reduced protein expression of IL-1β, TNF-α and IL-6 in peritoneal dialysate-induced rats. *** vs. blank, P < 0.001. ##, ### vs. PD, P < 0.01, P < 0.001.
Crocin regulated Wnt/β-catenin signaling in peritoneal dialysate-induced rats
Protein expression of Axin was decreased, while Wnt5a, β-Catenin, and COX-2 were increased in peritoneal tissues of peritoneal dialysate-induced rats (Figure 5). Crocin enhanced Axin, while it reduced Wnt5a, β- Catenin, and COX-2 (Figure 5) to suppress activation of Wnt/β-catenin signaling in peritoneal dialysate-induced rats.
Figure 5. Crocin regulated Wnt/β-catenin signaling in peritoneal dialysate-induced rats. Crocin enhanced Axin, while reduced Wnt5a, β-Catenin and COX-2 in peritoneal dialysate-induced rats. *** vs. blank, P < 0.001. ##, ### vs. PD, P < 0.01, P < 0.001.
Discussion
Ancient herbal medicine has been regarded as alternative therapy to treat chronic kidney diseases (Chen et al., 2021). For example, Saikosaponin D, a triterpene saponin, was isolated from herbal medicine Bupleurum falactum, and it suppressed peritoneal fibrosis in rats (Ruiqi et al., 2021). Crocus sativus has been shown to mediate macrophage polarization from M2 towards M1 and reduced peritoneal adhesion formation in rats (Rahmanian-Devin et al., 2021). Crocin, a bioactive carotenoid from Crocus sativus, exerted anti-fibrotic effect against carbon tetrachloride-induced liver fibrosis (Chhimwal et al., 2020), pulmonary fibrosis (Zaghloul et al., 2019), and isoprenaline-induced myocardial fibrosis (Jin et al., 2020). This study for the first time found that crocin also exerted anti-fibrotic and anti-inflammatory effects on peritoneal dialysate-induced rats.
Previous study has shown that peritoneal dialysis solution induced thickened compact zone, mesothelial denudation, and interstitial fibrosis in peritoneal tissues (Zhao et al., 2019). Moreover, the peritoneal transport function was also impaired by injection with peritoneal dialysis solution through downregulation of ultrafiltration volume and upregulation of mass glucose transport (Zhao et al., 2019). Therefore, peritoneal dialysate-induced rats were widely used as model of peritoneal fibrosis (Zhao et al., 2019). In this study, we also established a rat model with peritoneal fibrosis through administration with peritoneal dialysis solution. Peritoneal dialysis solution induced histopathological changes in peritoneum, and stimulated peritoneal fibrosis. Moreover, peritoneal dialysis solution also provoked impaired peritoneal function in rats. Crocin ameliorated histopathological changes in peritoneum of peritoneal dialysate-induced rats, attenuated peritoneal fibrosis, and improved the peritoneal function, suggesting a protective effect against peritoneal fibrosis in rats induced by peritoneal dialysis.
In clinical research, structure of the peritoneal membrane in patients with long-term continuous ambulatory peritoneal dialysis showed fibroblast characteristics and loss of mesothelial cell morphology due to the epithelial- mesenchymal transformation (Yáñez-Mó et al., 2003). High glucose concentration in peritoneal dialysate fluid altered normal polarity of peritoneal mesothelial cells, and induced myofibroblast phenotype of the mesothelial cells, thus leading to epithelial-mesenchymal transformation (Duan et al., 2021). The epithelial- mesenchymal transformation stimulated loss of tight junctions between mesothelial cells, and contributed to the development of peritoneal fibrosis (van baal et al., 2016). Downregulation of E-cadherin, and upregulation of fibronectin, α-SMA, and collagen I were typical features of peritoneal mesothelial cells during epithelial- mesenchymal transformation (Wei et al., 2019). Blockage of epithelial-mesenchymal transformation mitigated peritoneal fibrosis (Kim et al., 2019). Crocin has been reported to increase protein expression of E-cadherin, and decrease N-cadherin and Snail to suppress epithelial- mesenchymal transition of gastric cancer (Zhou et al., 2019). Results in this study also indicated that crocin enhanced the expression of E-cadherin, and reduced fibronectin, α-SMA, and collagen I to inhibit epithelial- mesenchymal transformation in peritoneal dialysate- induced rats.
Long-term exposure to peritoneal dialysate fluid–induced chronic inflammation in peritoneal tissues stimulated peritoneal vasculature changes and impairment of peritoneal membrane integrity, ultimately leading to peritoneal fibrosis (Witowski et al., 2018). Therefore, inflammation was implicated in the pathogenesis of peritoneal dialysis-induced fibrosis (Witowski et al., 2018). The anti-inflammatory effect of crocin has been widely found in carbon tetrachloride-induced liver fibrosis (Chhimwal et al., 2020), pulmonary fibrosis (Zaghloul et al., 2019), and isoprenaline-induced myocardial fibrosis (Jin et al., 2020). Crocin in this study also attenuated peritoneal dialysis-induced upregulation of IL-1β, TNF-α, and IL-6, revealing the anti-inflammatory effect of crocin against peritoneal dialysis-induced fibrosis. Oxidative stress was also implicated in the pathogenesis of peritoneal fibrosis, and herbal antioxidants showed a promising effect to treat peritoneal fibrosis (Asgharpour and Alirezaei, 2021). Crocin exerted anti-oxidant effect against myocardial infarction injury (Wang et al., 2018). Therefore, crocin might also exhibit anti-oxidant effect against peritoneal dialysis-induced fibrosis.
Wnt family contains a variety of secreted lipid-modified signaling glycoproteins that deliver signals to cells via the surface receptors (Khatoon et al., 2020). In typical reactions, Wnt ligands bind to receptor complex, resulting in nuclear translocation of β-catenin, and β-catenin has been shown to regulate the transcription of fibrosis- related genes, such as fibronectin, E-cadherin, α-SMA, and collagen I, in fibrotic diseases (Guo et al., 2012). Expression of Wnt1, Wnt5a, β-catenin, and lymphoid enhancer-binding factor 1 were upregulated in mesothelial cells of patients with peritoneal dialysis (Guo et al., 2017). Inhibition of Wnt/β-catenin signaling increased E-cadherin, and decreased α-SMA, collagen I, and fibronectin to attenuate mesothelial-mesenchymal transition in dialysate-induced peritoneal fibrosis (Guo et al., 2017). Crocin interfered with Wnt/β-catenin signaling to inhibit epithelial–mesenchymal transition of triple- negative breast cancer (Arzi et al., 2018; Dariushnejad et al., 2022). Moreover, crocin also inhibited activation of Wnt5a/β-catenin signaling to alleviate pain hyperalgesia in adjuvant-induced arthritis rats (Wang et al., 2020). Results in this study demonstrated that crocin reduced protein expression of Wnt5a and β-catenin in peritoneal dialysis-induced rats. Therefore, crocin might protect against peritoneal dialysis-induced fibrosis through inactivation of Wnt5a/β-catenin signaling.
Collectively, crocin attenuated peritoneal dialysate- induced histopathological changes and fibrosis in peritoneum of rats, and exerted anti-inflammatory and anti- fibrotic effects against peritoneal dialysis-induced fibrosis. Crocin inhibited epithelial-mesenchymal transformation in peritoneal dialysis-induced fibrotic rats through downregulation of Wnt5a/β-catenin signaling. Therefore, crocin might be a potential agent for the prevention of peritoneal dialysis-induced fibrosis.
Acknowledgements
Not applicable.
Funding
This work was supported by the Science and Technology Commission of Shanghai Municipality (No. 20ZR1449800).
Competing interests
The authors state that there are no conflicts of interest to disclose.
Ethics approval
Ethical approval was obtained from the Medical Ethics Committee of Pudong New Area People’s Hospital.
Data availability
The authors declare that all data supporting the findings of this study are available within the paper and any raw data can be obtained from the corresponding author upon request.
Contribution of authors
Jingyuan Lu and Danye Shi designed the experiments; Jingyuan Lu, Danye Shi, Xinhui Zhao, MInhui Xi, and Hualin Qi performed the experiments, and analyzed and interpreted the data; Qiang He prepared the manuscript with contributions from all co-authors. Jingyuan Lu and Danye Shi contributed equally to the work and should be considered co-first authors.
REFERENCES
Arzi, L., Farahi, A., Jafarzadeh, N., Riazi, G., Sadeghizadeh, M. and Hoshyar, R., 2018. Inhibitory effect of crocin on metastasis of triple-negative breast cancer by interfering with Wnt/β-Catenin pathway in murine model. DNA and Cell Biology 37: 1068–1075. 10.1089/dna.2018.4351
Asgharpour, M. and Alirezaei, A., 2021. Herbal antioxidants in dialysis patients: a review of potential mechanisms and medical implications. Renal Failure 43: 351–361. 10.1080/0886022X.2021.1880939
Chen, J., Tsim, K.W.K. and Zhao, Y.-Y., 2021. Editorial: applications of herbal medicine to control chronic kidney disease. Frontiers in Pharmacology 12: 742407. 10.3389/fphar.2021.742407
Chhimwal, J., Sharma, S., Kulurkar, P.M. and Patial, V., 2020. Crocin attenuates CCl4-induced liver fibrosis via PPAR03 mediated modulation of inflammation and fibrogenesis in rats. Human & Experimental Toxicology 39: 1639–1649. 10.1177/0960327120937048
Dariushnejad, H., Aljaf, K.A.H., Wasman, H.M., Pirzeh, L. and Ghorbanzadeh, V., 2022. Crocin inhibit the metastasis of MDA-MB-231 cell line by suppressing epithelial to mesenchymal transition through WNT/β-catenin signaling pathway. Research Square. 10.21203/rs.3.rs-1539821/v1
Duan, Z., Yao, J., Duan, N., Wang, M. and Wang, S., 2021. Sulodexide prevents peritoneal fibrosis by downregulating the expression of TGF-β1 and its signaling pathway molecules. Evidence-Based Complementary and Alternative Medicine 2021: 2052787. 10.1155/2021/2052787
Fang, K. and Gu, M., 2020. Crocin improves insulin sensitivity and ameliorates adiposity by regulating AMPK-CDK5-PPARγ signaling. BioMed Research International 2020: 9136282. 10.1155/2020/9136282
Guo, Y., Sun, L., Xiao, L., Gou, R., Fang, Y., Liang, Y., Wang, R., Li, N., Liu, F. and Tang, L., 2017. Aberrant Wnt/Beta-Catenin pathway activation in dialysate-induced peritoneal fibrosis. Frontiers in Pharmacology 8: 774–774. 10.3389/fphar.2017.00774
Guo, Y., Xiao, L., Sun, L. and Liu, F., 2012. Wnt/beta-catenin signaling: a promising new target for fibrosis diseases. Physiological Research 61: 337–346. 10.33549/physiolres.932289
Heo, J.-Y., Do, J.-Y., Lho, Y., Kim, A.Y., Kim, S.-W. and Kang, S.-H., 2021. TGF-β1 receptor inhibitor SB525334 attenuates the epithelial to mesenchymal transition of peritoneal mesothelial cells via the TGF-β1 signaling pathway. Biomedicines 9: 839. 10.3390/biomedicines9070839
Jin, W., Zhang, Y., Xue, Y., Han, X., Zhang, X., Ma, Z., Sun, S., Chu, X., Cheng, J., Guan, S., Li, Z. and Chu, L., 2020. Crocin attenuates isoprenaline-induced myocardial fibrosis by targeting TLR4/NF-κB signaling: connecting oxidative stress, inflammation, and apoptosis. Naunyn-Schmiedeberg’s Archives of Pharmacology 393: 13–23. 10.1007/s00210-019-01704-4
Kang, D.-H., 2020. Loosening of the mesothelial barrier as an early therapeutic target to preserve peritoneal function in peritoneal dialysis. Kidney Research and Clinical Practice 39: 136–144. 10.23876/j.krcp.20.052
Khan, S. and Rosner, M.H., 2018. Peritoneal dialysis for patients with end-stage renal disease and liver cirrhosis. Peritoneal Dialysis International 38: 397–401. 10.3747/pdi.2018.00008
Khatoon, E., Deka, N., Deka, M., Saikia, K.K., Baruah, M.N. and Ahmed, G.N., 2020. Clinical significance of β-catenin, hTERT, p53, and Wnt7A as biomarkers for ovarian cancer. EJGO 41: 181–187. 10.31083/j.ejgo.2020.02.5138
Kim, Y.C., Kim, K.H., Lee, S., Jo, J.-W., Park, J.Y., Park, M.-S., Tsogbadrakh, B., Lee, J.P., Lee, J.W., Kim, D.K., Oh, K.-H., Jang, I.-J., Kim, Y.S., Cha, R.-H. and Yang, S.H., 2019. ST2 blockade mitigates peritoneal fibrosis induced by TGF-β and high glucose. Journal of Cellular and Molecular Medicine 23: 6872–6884. 10.1111/jcmm.14571
Korte, M.R., Sampimon, D.E., Betjes, M.G.H. and Krediet, R.T., 2011. Encapsulating peritoneal sclerosis: the state of affairs. Nature Reviews Nephrology 7: 528–538. 10.1038/nrneph.2011.93
Liu, J., Feng, Y., Sun, C., Zhu, W., Zhang, Q.-Y., Jin, B., Shao, Q.-Y., Xia, Y.-Y., Xu, P.-F., Zhang, M. and Jiang, C.-M., 2020. Valsartan ameliorates high glucose-induced peritoneal fibrosis by blocking mTORC1 signaling. Experimental Biology and Medicine (Maywood, N.J.) 245: 983–993. 10.1177/1535370220919364
Liu, Q., Mao, H., Nie, J., Chen, W., Yang, Q., Dong, X. and Yu, X., 2008. Transforming growth factor β1 induces epithelial–mesenchymal transition by activating the Jnk–SMAD3 pathway in rat peritoneal mesothelial cells. Peritoneal Dialysis International: Journal of the International Society for Peritoneal Dialysis 28 (Suppl 3): S88–S95. 10.1177/089686080802803s18
Rahmanian-Devin, P., Rakhshandeh, H., Baradaran Rahimi, V., Sanei-Far, Z., Hasanpour, M., Memarzia, A., Iranshahi, M. and Askari, V.R., 2021. Intraperitoneal lavage with Crocus sativus prevents postoperative-induced peritoneal adhesion in a rat model: evidence from animal and cellular studies. Oxidative Medicine and Cellular Longevity 2021: 5945101. 10.1155/2021/5945101
Ruiqi, L., Ming, P., Qihang, S., Yangyang, L., Junli, C., Wei, L., Chao, G., Xinyue, L., Kang, Y. and Hongtao, Y., 2021. Saikosaponin D inhibits peritoneal fibrosis in rats with renal failure by regulation of TGFβ1/BMP7/Gremlin1/Smad pathway. Frontiers in Pharmacology 12: 628671. 10.3389/fphar.2021.628671
Tsuzuki, T., Iwata, H., Murase, Y., Takahara, T. and Ohashi, A., 2018. Renal tumors in end-stage renal disease: a comprehensive review. International Journal of Urology 25: 780–786. 10.1111/iju.13759
Vafaei, S., Wu, X., Tu, J. and Nematollahi-Mahani, S.N., 2022. The effects of crocin on bone and cartilage diseases. Frontiers in Pharmacology 12: 830331. 10.3389/fphar.2021.830331
van baal, J., Van de Vijver, K., Nieuwland, R., Noorden, C.J.F., Driel, W., Sturk, A., Kenter, G., Rikkert, L. and Lok, C., 2016. The histophysiology and pathophysiology of the peritoneum. Tissue and Cell 49: 95–105. 10.1016/j.tice.2016.11.004
Wang, J.-F., Xu, H.-J., He, Z.-L., Yin, Q. and Cheng, W., 2020. Crocin alleviates pain hyperalgesia in AIA rats by inhibiting the spinal Wnt5a/β-Catenin signaling pathway and glial activation. Neural Plasticity 2020: 4297483. 10.1155/2020/4297483
Wang, Y., Wang, Q., Yu, W. and Du, H., 2018. Crocin attenuates oxidative stress and myocardial infarction injury in rats. International Heart Journal 59: 387–393. 10.1536/ihj.17-114
Wei, X., Bao, Y., Zhan, X., Zhang, L., Hao, G., Zhou, J. and Chen, Q., 2019. MiR-200a ameliorates peritoneal fibrosis and functional deterioration in a rat model of peritoneal dialysis. International Urology and Nephrology 51: 889–896. 10.1007/s11255-019-02122-4
Witowski, J., Kamhieh-Milz, J., Kawka, E., Catar, R. and Jörres, A., 2018. IL-17 in peritoneal dialysis-associated inflammation and angiogenesis: conclusions and perspectives. Frontiers in Physiology 9: 1694. 10.3389/fphys.2018.01694
Wu, Z. and Hui, J., 2020. Crocin reverses 1-methyl-3-nitroso-1-nitroguanidine (MNNG)-induced malignant transformation in GES-1 cells through the Nrf2/Hippo signaling pathway. Journal of Gastrointestinal Oncology 11: 1242–1252. 10.21037/jgo-20-406
Yáñez-Mó, M., Lara-Pezzi, E., Selgas, R., Ramírez-Huesca, M., Domínguez-Jiménez, C., Jiménez-Heffernan, J.A., Aguilera, A., Sánchez-Tomero, J.A., Bajo, M.A., Álvarez, V., Castro, M.A., del Peso, G., Cirujeda, A., Gamallo, C., Sánchez-Madrid, F. and López-Cabrera, M., 2003. Peritoneal dialysis and epithelial-to-mesenchymal transition of mesothelial cells. New England Journal of Medicine 348: 403–413. 10.1056/NEJMoa020809
Zaghloul, M.S., Said, E., Suddek, G.M. and Salem, H.A., 2019. Crocin attenuates lung inflammation and pulmonary vascular dysfunction in a rat model of bleomycin-induced pulmonary fibrosis. Life Sciences 235: 116794. 10.1016/j.lfs.2019.116794
Zhao, J.-L., Guo, M.-Z., Zhu, J.-J., Zhang, T. and Min, D.-Y., 2019. Curcumin suppresses epithelial-to-mesenchymal transition of peritoneal mesothelial cells (HMrSV5) through regulation of transforming growth factor-activated kinase 1 (TAK1). Cellular & Molecular Biology Letters 24: 32. 10.1186/s11658-019-0157-x
Zhou, Y., Xu, Q., Shang, J., Lu, L. and Chen, G., 2019. Crocin inhibits the migration, invasion, and epithelial-mesenchymal transition of gastric cancer cells via miR-320/KLF5/HIF-1α signaling. Journal of Cellular Physiology 234: 17876–17885. 10.1002/jcp.28418