
Download
RESEARCH ARTICLE
Fungal bioconversion of brewery by-products: assessment of fatty acids and sterols profiles
Fatma Boukid1, 3*, Joan Pera2, Javier Parladé2, Massimo Castellari1
1Food Safety and Functionality Programme, Institute of Agriculture and Food Research and Technology (IRTA), Monells, Spain;
2Sustainable Plant Protection, IRTA, Centre de Cabrils, Barcelona, Spain;
3ClonBio Group LTD, 6 Fitzwilliam Pl, Dublin, Ireland
Abstract
Waste management of brewery by-products is economically and environmentally problematic. In the frame of bio-recycling, this study aims to investigate the bioconversion of brewery by-products by filamentous fungi. Pleurotus ostreatus and Lentinula edodes were grown on different substrates based on brewer’s spent grains (fresh and dry). Afterwards, fatty acids and sterols were determined. Following the selection of the suitable substrate composition for fungal growth, results showed that fatty acids composition of fungal biomasses varied significantly as a function of substrate and fungal strain. Interestingly, fungal fat might be used for human nutrition due to low SFA/UFA ratios (~0.2–0.4) within the same range of vegetal oils. Sterols profile of fungi biomass revealed the predominance of ergosterol. Also, it was found that the fungi growing on by-products slightly reduced the cholesterol contents. As such, this approach focusing on the bioconversion of by-products using fungi can provide biomasses with a fat composition suitable for feed and human consumption.
Key words: bioconversion, fatty acids, Lentinula edodes, Pleurotus ostreatus, sterols
*Corresponding Author: Fatma Boukid, Food Safety and Functionality Programme, Institute of Agriculture and Food Research and Technology (IRTA), Monells, Spain; and ClonBio Group LTD, 6 Fitzwilliam Pl, Dublin, Ireland. Email: [email protected]
Received: 11 September 2022; Accepted: 13 October 2022; Published: 11 November 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
By-products from the agro-industrial sector present crescent environmental and economic problems because handling large amounts of organic material requires high expenses and can negatively affect the environment (Barcelos et al., 2020; Correddu et al., 2020; Radenkovs et al., 2018). In most cases, this material is reutilized as fertilizers in the field, or burnt or drained as a waste (Bhuvaneshwari et al., 2019; Duque-Acevedo et al., 2020; Puglia et al., 2021). The proper valorization of by-products can efficiently reduce their environmental impact and obtain new strategic solutions to increase the productivity and sustainability of agro-industrial systems (Boukid et al., 2021a; Ferreira et al., 2022; Pauletto et al., 2020; Hamam et al., 2021; Pauletto et al. 2020; Boukid et al., 2021). In the European Union, the use and valorization of waste and by-products are considered the key elements in the Work Programme 2021–2022 (EU 2021).
The brewing industry is an important global business with huge annual revenues (US$ 651,398 million in 2021) with a compound annual growth rate of 7.4% from 2021 to 2025 (Statistica 2021). On the other hand, malting and brewing generate high amounts of by-products including brewer’s spent grain, hot trub, wastewater, spent hops, and residual brewer’s yeast (Jaeger et al., 2020; Karlović et al., 2020). Brewer’s spent grain, the main by-product (~85%), is known for its rich composition of proteins (20–30%), fiber (30–70%), lipids (5–10%), vitamins, and minerals (Fărcaş et al., 2017). Given this nutritious composition and affordability, the biotechnological valorization of such by-products is deemed necessary for economic and environmental motives (Berglund et al., 2016; Karlović et al., 2020; Rachwał et al., 2020; Severini et al., 2015). From a circular economy viewpoint, an integral bioremediation and valorization of brewery by-products relies on implementing bioreactors and identifying suitable microorganisms (e.g., yeast and fungi) to degrade this material and produce valuable compounds (Bianco et al., 2020; Gmoser et al., 2020; Mohajeri et al., 2021; Xiros and Studer 2017).
Filamentous fungi were demonstrated to be efficient decomposers of different substances and complexes biomass (Dai et al., 2018). Fungi could produce numerous extracellular enzymes, mainly hydrolases and oxidore-ductases. Phytase, laccase, pectinase, cellulases, xylanases, lipases, and tannase are among the most important enzymes produced by fungi (Campioni et al., 2019; Linhartová et al., 2020; Londoño-Hernandez et al., 2020). One of the main activities of these enzymes is the bioconversion of a wide variety of agricultural and/or industrial biomass (El-Gendi et al., 2022; Fernandes et al., 2020; Xu et al., 2018). Thus, fungi can be considered as cell factories that produce high-value compounds including cellulose, antibiotics, organic acids, pectin, inulin, proteins, and lipids (Liu et al., 2020). Furthermore, fungal enzymes are of high value and dominate nearly half of the enzyme market due to their availability, compatibility, cost-efficiency, and versatility of applications (e.g., medicine, biotechnology, food, leather, and textile (El-Gendi et al., 2022; Meyer et al., 2020; Pellegrino et al., 2022; Wang et al., 2018). Therefore, the use of fungi is a promising green approach toward the production of sustainable compounds, reduction in the cost of production, and mitigation of the impact of wastes on the environment which contributes to bio-economies and human health (Boukid and Gagaoua 2022; Meyer et al., 2020).
Brewer’s spent grains are lignocellulosic biomass rich in arabinoxylan (22−28%), cellulose (17−25%), and lignin (12−28%) (Fărcaş et al., 2017; Reis et al., 2015), and thus represent a good source of carbon for fungal growth. Several studies focused on the use of this by-product as a low-cost substrate for edible mushrooms (saprophytic fungi) able to degrade lignocellulosic materials (Fărcaş et al., 2017; Rachwał et al., 2020). Thus, the bioconversion of brewer’s spent grain using fungi can be considered a promising sustainable strategy (Dias et al., 2018; Marcus and Fox 2021). The most cultivated edible fungi are Lentinula (shiitake and relatives), Pleurotus (oyster mushroom), Auricularia (wood ear mushroom), and Agaricus (button mushroom and relatives) (Kumla et al., 2020). Fungi such as Rhizopus spp., Trichoderma spp., and Mucor spp. have been used to hydrolyze brewer’s grain spent, which resulted in producing nitrogen and carbon and releasing proteins and sugars for their growth that required no extra nutrients (Bekatorou et al., 2015; Cooray and Chen 2018). Aspergillus brasiliensis and Aspergillus ibericus produced lignocellulolytic enzymes using brewer’s grain spent as substrate (da Silva Menezes et al., 2017; Sousa et al., 2018). The enzymes such as xylanase and cellulase contribute to the increase in the release of primary and secondary metabolites (Verni et al., 2020). The nutritional value of the by-products was improved in terms of increasing protein content and producing value-added products such as lignocellulolytic enzymes and antioxidant peptides (da Silva Menezes et al., 2017; Sousa et al., 2018). Fungi Neurospora intermedia and Rhizopus oryzae were used to produce new protein- enriched products out of this by-product (Gmoser et al., 2020).
Past studies on the bioconversion of brewery by-products using fungi focused on the impact of substrate on fungal growth, yield, and nutritional composition of the biomass with emphasis on proteins, amino acids, fibers, and minerals (Hoa et al., 2015; Koutrotsios et al., 2014; Wang et al., 2001), while lipids were rarely targeted. Noteworthy, fungal lipids are gaining attraction in production of healthier and more sustainable alternatives for edible plant and/or animal lipids (Athenaki et al., 2018; Zhang et al., 2022). The interest in fungi as a source of proteins is not new and keeps increasing (Ciani et al., 2021; Ibarruri et al., 2021). This can be attributed to their white color and bland taste, and most importantly their filamentous texture that suits meat analog applications such as Quorn™ made using biomass fermentation (Fusarium venenatum) (Boukid et al., 2021b). Further understanding of the fat component of edible fungi could be of interest as a potential fat replacer to make healthy and sustainable plant-based products.
Therefore, this work aims to expand the knowledge on fungal efficiency and the bioconversion of by-products to fatty acids and sterols using edible fungi. Remarkably, Pleurotus ostreatus and Lentinula edodes, two widely used edible fungi, were cultivated using substrates made using brewery by-products. The first step consisted of the selection of suitable substrates for fungal growth. Afterward, the focus was attributed to assessing fatty acids and sterols profiles as a function of selected substrates.
Material and Methods
Material
For the purposes of this study, the fungal strains P. ostreatus (Jacq.) P. Kumm. (common name: Winter Oyster Mushroom) M2191 and L. edodes (Berk.) Pegler (common name: Shiitake) M3790 were purchased from Mycelia BVBA® (Veldeken, Belgium). Brewer’s spent grains were collected from a local brewery (Mahou San Miguel, Spain). Fresh brewer’s spent grain contained 75.3% moisture, 2.4% total fat, and 7.6% protein, while dried brewer’s spent grain contained 6.7% moisture content, 8.6% total fat, and 32.4% protein.
Substrate preparation and mushroom cultivation process
For cultivation, heat-resistant polypropylene bags (Sac O2®) were used. The formulation of substrates is reported in Table 1. All culture bags were filled with a final quantity of 4 L of substrate and autoclaved at 100°C for 2 h. The mushroom cultivation was performed with an inoculum concentration of 10% (dwt/dwt) (Hultberg et al., 2018). In all cases, the inoculation ratio used was 1:30 (inoculum:substrate, v:v). The applied dose was higher than that usually used in the industrial cultivation of mushrooms, which allowed to shorten the incubation times (Aranaz et al., 2021; Hultberg et al., 2018). The viability of all the inoculums was verified in each experimental batch by seeding an aliquot on 3% malt extract agar plates.
Table 1. Formulation of substrates based on selected by-products.
| Substrate | Composition |
|---|---|
| Control | Chestnut wood chips (1500 mL) + chestnut sawdust (1000 mL) + cereal seeds (equal parts of corn, barley, and wheat) (92.6 g) adjusted to 60% humidity and a pH of 5.5–6 |
| A* | Dry brewer’s spent grain (3 L) + H2O (1.8 L) + control (1:3, v:v) |
| B* | Dry brewer’s spent grain (1 L) + yeast lysate (500 mL) + H2O (500 mL) + control (1:3, v:v) |
| C* | Fresh brewer’s spent grain + control (1:3, v:v) |
| D* | Fresh brewer’s spent grain + control (1:1, v:v) |
*The pH of the substrates made with brewer’s spent grain was adjusted by adding calcium carbonate (40 g/kg substrate) to reach 5.4.
Once inoculated, the bags were incubated at a temperature of 22–25°C for 5–7 weeks. After the incubation period, the substrate and the apparent absence of contaminants were verified, and the bags were transferred to fruiting chambers (temperature: 15 ± 3°C; relative humidity: 80–90%) to produce mushrooms. In all the experiments, a commercial substrate of edible mushrooms was used as a control. For each experiment, four replications were carried out.
Fatty acids profile
Fatty acids profile was assessed using a method reported in a previous work by Riudavets et al., (2020). In brief, samples (500 mg) were extracted using a mixture of chloroform and methanol (2:1, v/v), derivatized with a mixture of toluene and 3 N HCl in methanol (1:4, v/v) at 80°C for 1 h and added with 10% NaCl in water and hexane (10:3, v/v). Fatty acids methyl esters were recovered in the organic phase and then separated on a gas chromatograph, Agilent 6890 Series II (Hewlett Packard SA, Barcelona, Spain), equipped with a capillary column DB23 (30 m × 0.25 mm i.d., 0.25 μm; Agilent, Santa Clara, USA), a split or splitless injector, and a flame ionization detector. Identification of single methyl esters was performed by comparing retention time of the peaks with those of pure standards; while quantification was carried out using tripentadecanoin (Merck KGaA, Darmstadt, Germany) as an internal standard.
Sterol profile
The lyophilized fungi (500 mg) were hydrolyzed in Soxcap (FOSS IBERIA, S.A., Barcelona, Spain) with 4 N HCl for 5 h. Subsequently, an extraction was carried out with 350 mL of hexane: diethyl ether (2:1, v/v). The internal standard (5α-cholestan-3β-ol) was added to the evaporated extract, and saponification was carried out using 8 mL of 9 N KOH for 3 h at 80°C. For the extraction of the unsaponified fraction, three aliquots of 12 mL of hexane and diethyl ether (2:1) and 4 mL of ethanol were added. The apolar fractions of the three extractions were dried and then derivatized with 50 μL of silanizing solution (Silylating mixture I according to Sweeley, Sigma Aldrich, Missouri, US) for 1 h at 80°C. The derivatized sample was dried using nitrogen flow, resuspended with 1 mL isooctane and 2-propanol and injected into the chromatographic equipment.
For chromatographic analysis, a CP-3800 gas chromatograph (Varían) equipped with a DB-5MS column (length: 30 m, diameter: 0.250 mm diameter, film thickness: 0.25 μm; Agilent Technologies, Santa Clara, US) was used. For chromatographic separation, an initial oven temperature of 80°C was used, with a gradient of 10°C/min up to 160°C, a gradient of 5°C/min up to 250°C, and a gradient of 1°C/min up to 285°C.
Statistical analysis
Fatty acids and sterols composition was determined in triplicate, and data were expressed as means ± standard deviations (SD). The Kruskal–Wallis test was performed to verify the substrate on fatty acids, sterols, and lipid indices as well as sterols. These tests were performed at a significance level of α = 0.05. Finally, to verify the influence of fungi on fatty acids profile, principal components analysis was performed. All the statistical analyses were performed using IBM SPSS 24 statistical software (SPSS Inc., Chicago, IL, USA).
Results and Discussion
Substrate selection
Fungi showed different behaviors toward the substrates as illustrated in Table 2. P. ostreatus was able to grow on all substrates and yielded higher biomass than the control. This fungus is known to produce enzymes including lipase, phytase, xylanase, β-galactosidase, and cellulase that can break down organic biomass and use for its growth (Naim et al., 2020). The quantity of biomass cultivated on dry brewer’s spent grains was higher than that that on fresh brewer’s spent grains. This can be explained by the higher amount of nutrients in dry substrate compared to that in the fresh (less dry amount) one. Particularly, Substrate D ensured the highest biomass production. On the other hand, L. edodes was able to grow on all substrates except B. Substrate C made with a ratio of 1:3 (v/v) yielded the highest biomass, while the remaining substrates (A and D) had low yields. Compared to P. ostreatus, L. edodes showed lower growth rate, which can be attributed to the higher enzymatic activity in P. ostreatus favoring substrate degradation and thus fungal growth (Challa et al., 2019).
Table 2. Biomass resulting from fungi growth on different by-products substrates.
| Substrates | Pleurotus ostreatus | Lentinula edodes | ||
|---|---|---|---|---|
| Growth | Biomass (g) | Growth | Biomass (g) | |
| Control | + | 173 | + | 217 |
| A | + | 243 | ± | 99 |
| B | + | 208 | - | 0 |
| C | + | 300 | + | 293 |
| D | + | 324 | ± | 84 |
+ Growth and production comparable to control; ± Growth and production much lower than control; - No growth or production.
In the following sections, brewery by-products substrates yielding the highest biomass were retrieved and compared to that of the control. Therefore, for P. ostreatus, C and D are considered, and C was considered for L. edodes.
Impact of selected substrates on fatty acids profile
Irrespective to the substrates, six fatty acids (C14:0, C15:0, C16:0, C18:0, C18:1 (n-6c), and C18:2 (n-6c)) were identified in P. ostreatus biomass (Table 3). The main fatty acids found were C18:2 (n-6c) followed by C16:0 and C18:2 (n-6c), which was consistent with the previous works (Cardoso et al., 2020; Gnanwa et al., 2021; Pedneault et al., 2007). Results showed variations in the percentages of the single components and their concentrations, without significantly modifying the general profiles. More specially, substrates did not impact the percentage of fatty acids (C14:0, C15:0, C16:0, C18:1[n-9c] and C18:2[n-6c]) except C18:0. This indicates that by-products did not change the fatty acids as compared to the control. The concentration of C14:0 was significantly higher in the control than in biomasses grown on brewery by-products, while C16:0 was higher in C than in the control and D. As for C18:2(n-6c), C had significantly high values than that of the control and D. Total fatty acids varied significantly with higher values in C than those of the control and D, which showed that lower concentration of fresh spent grains favored formation of fatty acids. Similar trend was observed for monounsaturated fatty acids (MUFA), polyunsaturated fatty acids (PUFA), and ω6. Total saturated fatty acid (SFA) was significantly higher in C. No significant differences were found in UFA and SFA/UFA as a function of substrates. The study results revealed PUFA as the dominant fatty acids, and this finding aligned with the previous studies focused on the same fungus but using different substrates (Cardoso et al., 2020; Gnanwa et al., 2021). Overall, growing P. ostreatus on brewery by-products did not affect the composition of fatty acids, suggesting their potential use instead of others expensive substrates. Noteworthy, all SFA/UFA were lower than 0.5, indicating that the fat of these biomasses could be a good candidate for use in human and/or animal foods (Sinanoglou et al., 2015; Wołoszyn et al., 2020). Indeed, the fatty acids profile was found comparable to that of soybean (C16:0: 5.5%; C18:2 [n-6c]: 33.2%) and peanut oils (C16:0: 9.55%; C18:2 [n-6c]: 65.8%) (Adjepong et al., 2017).
Table 3. Percentages and concentration of fatty acids and lipid indices in Pleurotus ostreatus biomasses grown on different substrates.
| Fatty acids | Control | C | D | Significance |
|---|---|---|---|---|
| C14:0 (%) | 0.281 ± 0.001 | 0.332 ± 0.071 | 0.401 ± 0.070 | NS |
| C15:0 (%) | 4.12 ± 0.69 | 3.75 ± 0.21 | 4.08 ± 0.11 | NS |
| C16:0 (%) | 18.09 ± 1.75 | 20.9 ± 0.65 | 21.65 ± 1.83 | NS |
| C18:0 (%) | 1.75 ± 0.45b | 0.96 ± 0.13a | 1.1 ± 0.08a | * |
| C18:1(n-9c) (%) | 9.88 ± 0.89 | 6.17 ± 0.35 | 6.32 ± 0.35 | NS |
| C18:2(n-6c) (%) | 65.87 ± 2.25 | 67.88 ± 1.19 | 66.45 ± 2.37 | NS |
| C14:0 (mg/100 mg) | 0.002 ± 0.001a | 0.004 ± 0.001b | 0.004 ± 0.001b | * |
| C15:0 (mg/100 mg) | 0.036 ± 0.003 | 0.042 ± 0.004 | 0.036 ± 0.003 | NS |
| C16:0 (mg/100 mg) | 0.157 ± 0.008a | 0.234 ± 0.023b | 0.194 ± 0.030a | * |
| C18:0 (mg/100 mg) | 0.015 ± 0.004 | 0.011 ± 0.002 | 0.010 ± 0.001 | NS |
| C18:1(n-9c) (mg/100 mg) | 0.086 ± 0.015 | 0.069 ± 0.010 | 0.057 ± 0.007 | NS |
| C18:2(n-6c) (mg/100 mg) | 0.579 ± 0.039a | 0.757 ± 0.062b | 0.594 ± 0.017a | * |
| Lipid indices | ||||
| Total | 0.875 ± 0.106a | 1.116 ± 0.097b | 0.895 ± 0.059a | * |
| SFA | 0.210 ± 0.005a | 0.290 ± 0.028a,b | 0.245 ± 0.035b | * |
| MUFA | 0.086 ± 0.015 | 0.069 ± 0.011 | 0.057 ± 0.007 | NS |
| PUFA | 0.579 ± 0.039a | 0.757 ± 0.062b | 0.594 ± 0.017a | * |
| ω6 | 0.580 ± 0.150a | 0.760 ± 0.06b | 0.5911 ± 0.017a | * |
| UFA | 0.665 ± 0.089 | 0.827 ± 0.072 | 0.651 ± 0.024 | NS |
| SFA/UFA | 0.321±0.041 | 0.351 ± 0.017 | 0.375 ± 0.039 | NS |
MUFA=sum of all monounsaturated fatty acids; PUFA =sum of all polyunsaturated fatty acids; SFA=sum of all saturated fatty acids; UFA = MUFA + PUFA; SFA/UFA = ratio of total saturated fatty acids/total monounsaturated fatty acids + total polyunsaturated fatty acids. Values are shown as means ± standard deviation.
Means within a row with different letter differ significantly, *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; NS = not significant.
Table 4 shows the fatty acids profile and lipid indices of L. edodes as a function of different substrates. Regardless of substrates, of the eight fatty acids (C14:0, C15:0, C16:0, C16:1 [n-7c], C18:0, C18:1 [n-9c], C18:2 [n-6c], and C18:3 [n-6c]), C18:2 (n-6c) was the most abundant, followed by C16:0. This aligns with the previous study findings of L. edodes cultivated using oak tree sawdust and rice (or wheat) bran (Chung et al., 2020). No significant differences were found among the percentages of fatty acids as a function of substrate. Nevertheless, the biomass of concentrations C16:0, C18:0, and C18:1(n-9c) was found to be significantly higher when grown on the control than on by-products. Total fatty acids, PUFA, ω6, and UFA were found to be unaffected by the substrate. As for SFA and MUFA, the control had higher values than the biomass grown on brewery by-products. Noteworthy, low SFA/UFA values (around 0.2) were obtained, which were within the same range of that of canola oil (Tartrakoon et al., 2016). Further investigation is needed to study the application of L. edodes grown on brewery by-products as potential animal-free fat substitute.
Table 4. Percentages and concentration of fatty acids and lipid indices in the biomasses of Lentinula edodes grown on different substrates.
| Fatty acids | Control | C | Significance |
|---|---|---|---|
| C14:0 | 0.190 ± 0.040 | 0.260 ± 0.031 | NS |
| C15:0 | 1.68 ± 0.73 | 1.48 ± 0.28 | NS |
| C16:0 | 14.24 ± 1.41 | 13.75 ± 0.72 | NS |
| C16:1(n-7c) | 0.340 ± 0.071 | 0.361 ± 0.030 | NS |
| C18:0 | 1.57 ± 0.45 | 0.75 ± 0.04 | NS |
| C18:1(n-9c) | 1.62 ± 0.24 | 1.75 ± 0.35 | NS |
| C18:2(n-6c) | 79.88 ± 2.73 | 81.01 ± 1.42 | NS |
| C18:3(n-6c) | 0.47 ± 0.21 | 0.63 ± 0.02 | NS |
| C14:0 (mg/100 mg) | 0.003 ± 0.001 | 0.003 ± 0.001 | NS |
| C15:0 (mg/100 mg) | 0.029 ± 0.011 | 0.017 ± 0.001 | NS |
| C16:0 (mg/100 mg) | 0.246 ± 0.002 | 0.164 ± 0.025 | * |
| C16:1(n-7c) (mg/100 mg) | 0.006 ± 0.001 | 0.004 ± 0.001 | NS |
| C18:0 (mg/100 mg) | 0.027 ± 0.005 | 0.009 ± 0.001 | * |
| C18:1(n-9) (mg/100 mg) | 0.028 ±0.001 | 0.021 ± 0.000 | * |
| C18:2(n-6c) (mg/100 mg) | 1.388 ± 0.194 | 0.973 ± 0.217 | NS |
| C18:3(n-6c) (mg/100 mg) | 0.008 ± 0.004 | 0.008 ± 0.002 | NS |
| Lipid indices | |||
| Total | 1.735 ± 0.184 | 1.199 ± 0.246 | NS |
| SFA | 0.304 ± 0.013 | 0.193 ± 0.027 | * |
| MUFA | 0.034 ± 0.002 | 0.025 ± 0.001 | * |
| PUFA | 1.397 ± 0.199 | 0.981 ± 0.218 | NS |
| ω6 | 1.4 ± 0.2 | 0.98 ± 0.22 | NS |
| UFA | 1.43 ± 0.197 | 1.005 ± 0.219 | NS |
| SFA/UFA | 0.215 ± 0.039 | 0.194 ± 0.015 | NS |
MUFA=sum of all monounsaturated fatty acids; PUFA =sum of all polyunsaturated fatty acids; SFA=sum of all saturated fatty acids; UFA = MUFA + PUFA; SFA/UFA = ratio of total saturated fatty acids/total monounsaturated fatty acids + total polyunsaturated fatty acids. Values are shown as means ± standard deviation.
Significance: *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; NS = not significant.
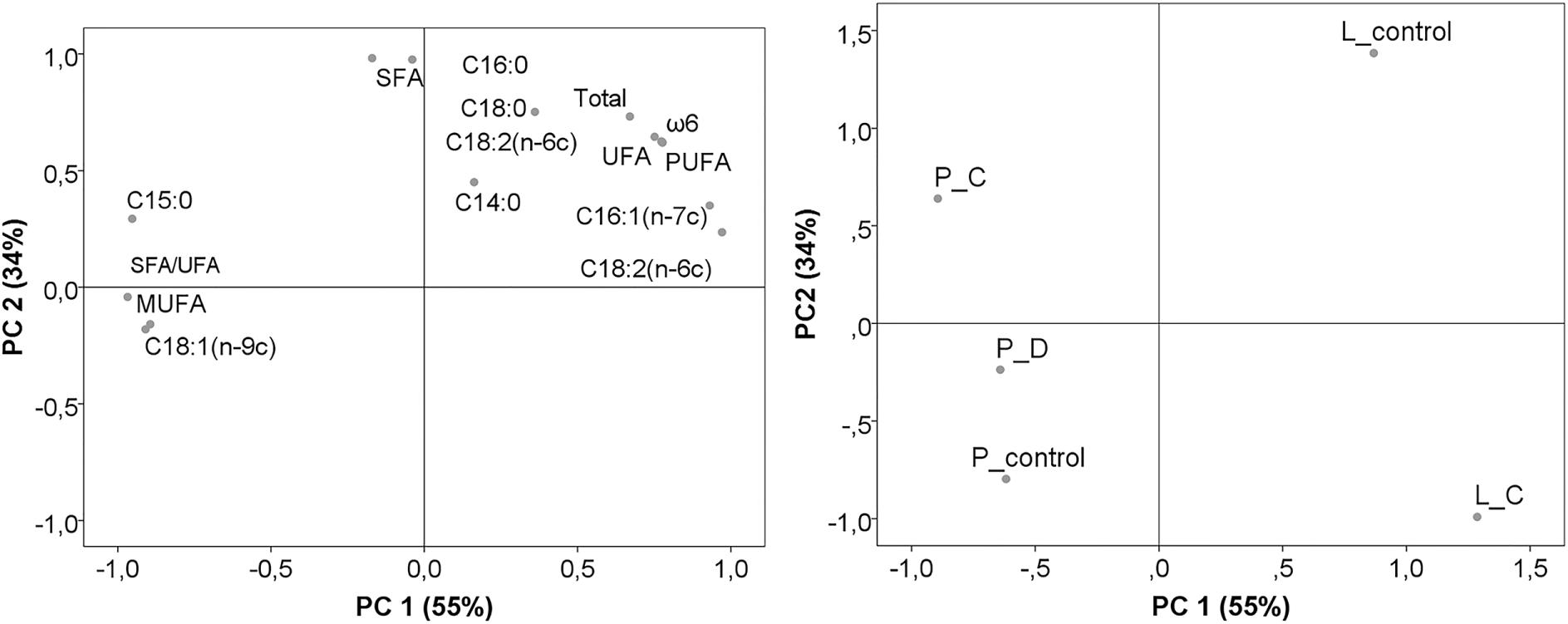
Principal component analysis based on fatty acid concentrations and lipid indices enabled a better understanding of the impacts of substrate and fungi simultaneously. The overall variability explained by the first two components was 89%, where the first component (PC1) accounted for 55% and the second (PC2) for 34%. As illustrated in Figure 1A, PC1 was explained as a function of C18:3(n-6c), SFA/UFA, C15:0, C16:1(n-7c), C18:1(n-9c), MUFA, ω6, PUFA, C18:2(n-6c) and UFA; whereas PC2 was a function of SFA, C16:0, C18:0, total fatty acids, and C14:0.
The projection the substrate and fungal strains on the factorial spaces created by the first two components showed that each fungus took a side of PC1 (L. edodes was on the positive side and P. ostreatus on the negative side) (Figure 1B). Variability between fatty acid compositions of fungal biomass can be attributed to different enzyme activities and types (lipase) responsible for fatty acid release and degradation (Chandra et al., 2020; López-Fernández et al., 2020). L. edodes control and that grown on by-products were located in opposite sides of PC2, where the control had the highest values of C18:3(n-6c), C16:1(n-7c), ω6, SFA, PUFA, C18:0, C14:0, C16:0, C18:2(n-6c), UFA, and total fatty acids, contrary to fungus grown on C. As for P. ostreatus, three biomasses were characterized by high amounts of C15:0, MUFA, SFA/UFA, and C18:1(n-9c). These results confirmed the variability between substrates and fungus, yet no clear clustering was found.
Figure 1. Scattering the data of fatty acids and lipid indices by the first two principal components (PC1 and PC2) analysis of fungi grow on brewery by-products. (A) Biplot of the first two components created, considering fatty acids and lipid indices; (B) Rotated principal scores of fungi and their corresponding substrates projected into the first two principal components. P_control: P. ostreatus grown on the control substrate; P_C: P. ostreatus grown on C substrate; P_D: P. ostreatus grown on D substrate; L_control: L. edodes grown on control substrate; L_C: L. edodes grown on C substrate.
Impact of selected substrates on sterols profile
Sterol profiles of fungal biomass are summarized in Tables 5 and 6. Regardless of the used fungus, no significant differences were found among sterols as a function of substrate except for estigmastanol in the case of P. ostreatus. In this case, the control had higher value than that of biomass grown on by-products. In L. edodes, only cholesterol and campesterol contents were significantly different from the control. Cholesterol was found higher in the control, indicating that growing fungi on by- products ensured a low cholesterol, possibly due to the degradation of cholesterol deriving from the substrate by fungi to use for their metabolism (El-Gendy et al., 2016; Hyde et al., 2019). This aligns with the evidence from previous studies that indicated the effectiveness of fungi in lowering cholesterol levels (Keong 2015).
Table 5. Concentration (mg/g) of phytosterols and cholesterol in Pleurotus ostreatus biomasses as a function of substrate.
| Control | C | D | Significance | |
|---|---|---|---|---|
| Cholesterol | 0.043 ± 0.032 | 0.032 ± 0.018 | 0.027 ± 0.006 | NS |
| Ergosterol | 0.252 ± 0.035 | 0.190 ± 0.064 | 0.223 ± 0.020 | NS |
| Campesterol | 0.059 ± 0.019 | 0.025 ± 0.006 | 0.038 ± 0.014 | NS |
| Stigmaesterol | 0.002 ± 0.003 | n.d. | n.d. | NS |
| ß-sitoesterol | 0.018 ± 0.003 | 0.009 ± 0.001 | 0.013 ± 0.012 | NS |
| Estigmastanol | 0.02 ± 0.011b | 0.001 ± 0.002a | 0.003 ± 0.002a | ** |
Values are shown as means ± standard deviation.
Means within a row with different letter differ significantly, *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; NS = not significant.
Table 6. Concentration (mg/g) of phytosterols and cholesterol in Lentinula edodes biomasses as a function of substrate.
| Control | C | Significance | |
|---|---|---|---|
| Cholesterol | 0.048 ± 0.006 | 0.011 ± 0.004 | * |
| Ergosterol | 0.225 ± 0.117 | 0.274 ± 0.072 | NS |
| Campesterol | 0.070 ± 0.001 | 0.124 ± 0.018 | * |
| Stigmaesterol | n.d. | 0.019 ± 0.018 | NS |
| ß-sitoesterol | 0.056 ± 0.032 | 0.063 ± 0.019 | NS |
| Estigmastanol | 0.017 ± 0.015 | 0.029 ± 0.004 | NS |
Values are shown as means ± standard deviation.
Significance: *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001; NS = not significant.
Irrespective of substrate, P. ostreatus profile was characterized by the predominance of ergosterol and campesterol, and traces of other phytosterols and cholesterol similar to that of L. edodes. The amount of ergosterol was double that of fungi cultivated on sugarcane bagasse–based substrates (Cardoso et al., 2020), this finding was in alignment with that of the data reported in a systematic review by Weete et al., (2010). Ergosterol is a metabolite synthesized by fungi, which cannot be synthesized by plants (Baur et al., 2016). This suggests the potential valorization of these biomasses as a source of vitamin D in contrary to most vegetable oils (e.g., peanut, soybean, rapeseeds, and olives) characterized by β-sitosterol and campesterol as the predominant phytosterols (Yang et al., 2019). Furthermore, both fungi had relevant amounts of total phytosterol comparable to soybean, peanut, and olive oils (~0.300 mg/100 mg) but lower than rice bran (1.89 mg/100 mg), corn (0.990 mg/100 mg), and rapeseed oils (0.893 mg/100 g) (Yang et al., 2019).
Conclusion
To conclude, the use of brewery by-products as substrates was efficient to ensure the fungal growth. Fatty acids composition was significantly impacted by fungal strain and substrate formulation. In P. ostreatus biomass, six fatty acids were found, where C18:2 (n-6c) was the most dominant. As for L. edodes, eight fatty acids were identified, with C18:2 (n-6c) being the most abundant. Interestingly, both fungi used in this study resulted in different fatty acids profiles that are comparable to vegetable oils with the advantage of having low SFA/UFA ratios. Sterols profile showed that regardless of the fungal strain, no significant differences were found among sterols as a function of substrate except for estigmastanol (P. ostreatus). Ergosterol was found as the primary sterol suggesting the potential valorization of these biomasses as a source of vitamin D. In addition, fungal biomass showed a low cholesterol level suggesting the cholesterol-lowering effect of fungus. This might be used as a mitigation strategy and thus valorize a wider range of by-products having a high initial cholesterol. Overall, these findings confirm the efficiency of fungi as bioconvertors of agro-industrial by-products to produce fats with interesting properties. More investigations are required to determine their functionality and applicability as a nonanimal fat substitute in food reformulation.
Funding
The authors are grateful for the financial support of the CIEN project BIOPRO from “Centro para el Desarrollo Tecnológico Industrial” (CDTI), Ministry of Science and Innovation, Government of Spain. This research was also funded by the CERCA Program (Centres de Recerca de Catalunya, Spain) of the Generalitat de Catalunya.
Acknowledgments
The authors would like to thank Sr. Carles Díaz from Teb Verd / Bolet Ben Fet (Barcelona, Spain) for the technical advice, and Narcís Sais, Teresa Fernandez Felipe, and Victòria Barnés for their technical assistance. They also thank Mahou S.A. for providing the agro-industrial by-products.
Conflicts of Interest
The authors have no conflicts of interest to disclose.
REFERENCES
Adjepong, M., Valentini, K., Pickens, C.A., Li, W., Appaw, W. and Fenton, J., 2017. Quantification of fatty acid and mineral levels of selected seeds, nuts, and oils in Ghana. Journal of Food Composition and Analysis 59: 43–49. 10.1016/j.jfca.2017.02.007
Aranaz, P., Peña, A., Vettorazzi, A., Fabra, M.J., Martínez-Abad, A., López-Rubio, A., et al., 2021. Grifola frondosa (Maitake) extract reduces fat accumulation and improves health span in C. elegans through the Daf-16/Foxo and Skn-1/Nrf2 signalling pathways. Nutrients 13(11): 3968. 10.3390/NU13113968/S1
Athenaki, M., Gardeli, C., Diamantopoulou, P., Tchakouteu, S.S., Sarris, D., Philippoussis, A., et al., 2018. Lipids from yeasts and fungi: physiology, production and analytical considerations. Journal of Applied Microbiology 124(2): 336–367. 10.1111/JAM.13633
Barcelos, M.C.S., Ramos, C.L., Kuddus, M., Rodriguez-Couto, S., Srivastava, N., Ramteke, P.W., et al., 2020. Enzymatic potential for the valorization of agro-industrial by-products. Biotechnology Letters 42(10): 1799–1827. 10.1007/S10529-020-02957-3
Baur, A.C., Brandsch, C., König, B., Hirche, F. and Stangl, G.L., 2016. Plant oils as potential sources of vitamin D. Frontiers in Nutrition 3: 12. 10.3389/fnut.2016.00029
Bekatorou, A., Plessas, S. and Mantzourani, I., 2015. Biotechno-logical exploitation of brewery solid wastes for recovery or production of value-added products. In: Rai, V.R., editor. Advances in food biotechnology. Chichester: Wiley Blackwell. p. 395–413.
Berglund, L., Noël, M., Aitomäki, Y., Öman, T. and Oksman, K., 2016. Production potential of cellulose nanofibers from industrial residues: efficiency and nanofiber characteristics. Industrial Crops and Products 92: 84–92. 10.1016/j.indcrop.2016.08.003
Bhuvaneshwari, S., Hettiarachchi, H. and Meegoda, J.N., 2019. Crop residue burning in India: policy challenges and potential solutions. International Journal of Environmental Research and Public Health 16(5): 832. 10.3390/IJERPH16050832
Bianco, A., Budroni, M., Zara, S., Mannazzu, I., Fancello, F. and Zara, G., 2020. The role of microorganisms on biotransformation of brewers’ spent grain. Applied Microbiology and Biotechnology. 104(20): 8661–8678. 10.1007/s00253-020-10843-1
Boukid, F. and Gagaoua, M., 2022. Meat alternatives: a proofed commodity? Advances in Food and Nutrition Research 101: 213–236. 10.1016/BS.AFNR.2022.02.003
Boukid, F., Riudavets, J., Del Arco, L., Castellari, M., Savoldelli, S., Spranghers, T., et al., 2021a. Impact of diets including agro-industrial by-products on the fatty acid and sterol profiles of larvae biomass from Ephestia kuehniella, Tenebrio molitor and Hermetia illucens. Insects 12(8): 672. 10.3390/INSECTS12080672
Boukid, F., Rosell, C.M., Rosene, S., Bover-Cid, S. and Castellari, M., 2021b. Non-animal proteins as cutting-edge ingredients to reformulate animal-free foodstuffs: present status and future perspectives. Critical Reviews in Food Science and Nutrition 62(23): 6390–6420. 10.1080/10408398.2021.1901649
Campioni, T.S., de Jesus Moreira, L., Moretto, E., Nunes, N.S.S. and de Oliva Neto, P., 2019. Biobleaching of kraft pulp using fungal xylanases produced from sugarcane straw and the subsequent decrease of chlorine consumption. Biomass and Bioenergy 121: 22–27. 10.1016/J.BIOMBIOE.2018.12.014
Cardoso, R.V.C., Carocho, M., Fernandes, A., Zied, D.C., Cobos, J.D.V., González-Paramás, A.M., et al. 2020. Influence of calcium silicate on the chemical properties of Pleurotus ostreatus Var. Florida (Jacq.) P. Kumm. Journal of Fungi 6(4): 1–16. 10.3390/JOF6040299
Challa, S., Dutta, T. and Neelapu, N.R.R., 2019. Fungal white biotechnology applications for food security: opportunities and challenges. In: Yadav, A.N., Singh, S., Mishra, S., Gupta, A., editors. Recent advancement in white biotechnology through fungi. 1st ed. Cham: Springer. p. 119–148. 10.1007/978-3-030-14846-1_4
Chandra, P., Enespa, Singh, R. and Arora, P.K., 2020. Microbial lipases and their industrial applications: a comprehensive review. Microbial Cell Factories 19(1): 169. 10.1186/S12934-020-01428-8/FIGURES/8
Chung, Ill M., Kim, S.Y., Han, J.G., Kong, W.S., Jung, M.Y. and Kim, S.H., 2020. Fatty acids and stable isotope ratios in Shiitake mushrooms (Lentinula edodes) indicate the origin of the cultivation substrate used: a preliminary case study in Korea. Foods 9(9): 1210. 10.3390/FOODS9091210
Ciani, M., Lippolis, A., Fava, F., Rodolfi, L., Niccolai, A. and Tredici, M.R., 2021. Microbes: food for the future. Foods 10(5): 971. http://www.ncbi.nlm.nih.gov/pubmed/33925123
Cooray, S.T. and Chen, W.N., 2018. Valorization of brewer’s spent grain using fungi solid-state fermentation to enhance nutritional value. Journal of Functional Foods 42: 85–94. 10.1016/j.jff.2017.12.027
Correddu, F., Lunesu, M.F., Buffa, G., Atzori, A.S., Nudda, A., Battacone, G., et al., 2020. Can agro-industrial by-products rich in polyphenols be advantageously used in the feeding and nutrition of dairy small ruminants? Animals 10(1): 131. 10.3390/ANI10010131
Dai, Z., Zhou, H., Zhang, S., Gu, H., Yang, Q., Zhang, W., et al. 2018. Current advance in biological production of malic acid using wild type and metabolic engineered strains. Bioresource Technology 258: 345–353. 10.1016/j.biortech.2018.03.001
da Silva Menezes, B., Rossi, D.M. and Ayub, M.A.Z., 2017. Screening of filamentous fungi to produce xylanase and xylooligosaccharides in submerged and solid-state cultivations on rice husk, soybean hull, and spent malt as substrates. World Journal of Microbiology and Biotechnology 33(3): 1–12. 10.1007/s11274-017-2226-5
Dias, A.A., Fernandes, J.M.C., Sousa, R.M.O.F., Pinto, P.A., Amaral, C., Sampaio, A., et al., 2018. Fungal conversion and valorization of winery wastes. In: Prasad, R., editor. Mycoremediation and environmental sustainability. 1st ed. Cham: Springer. p. 239–252. 10.1007/978-3-319-77386-5_9
Duque-Acevedo, M., Belmonte-Ureña, L.J., Cortés-García, F.J. and Camacho-Ferre, F., 2020. Agricultural waste: review of the evolution, approaches and perspectives on alternative uses. Global Ecology and Conservation 22: e00902. 10.1016/J.GECCO.2020.E00902
El-Gendi, H., Saleh, A.K., Badierah, R., Redwan, E.M., El-Maradny, Y.A. and El-Fakharany, E.M., 2022. A comprehensive insight into fungal enzymes: structure, classification, and their role in mankind’s challenges. Journal of Fungi 8(1): 23. 10.3390/JOF8010023
El-Gendy, M.M.A.A., Al-Zahrani, H.A.A. and El-Bondkly, A.M.A., 2016. Genome shuffling of mangrove endophytic Aspergillus luchuensis MERV10 for improving the cholesterol-lowering agent lovastatin under solid state fermentation. Mycobiology 44(3): 171–179. 10.5941/MYCO.2016.44.3.171
EU, 2021. Horizon Europe—work programme 2021–2022. Food, bioeconomy, natural resources, agriculture and environment. European Commission. 571 p.
Fărcaş, A.C., Socaci, S.A., Mudura, E., Dulf, F.V., Vodnar, D.C., Tofană, M., et al., 2017. Exploitation of brewing industry wastes to produce functional ingredients. In: Kanauchi, M., editor. Brewing technology. London, UK: InTechOpen. 10.5772/intechopen.69231
Fernandes, J.M.C., Fraga, I., Sousa, R.M.O.F., Rodrigues, M.A.M., Sampaio, A., Bezerra, R.M.F., et al., 2020. Pretreatment of grape stalks by fungi: effect on bioactive compounds, fiber composition, saccharification kinetics and monosaccharides ratio. International Journal of Environmental Research and Public Health 17(16): 1–13. 10.3390/ijerph17165900
Ferreira, D.S., Rocha, J.C.B., Arellano, D.B. and Pallone, J.A.L., 2022. Discrimination of South American grains based on fatty acid. Quality Assurance and Safety of Crops & Foods 14(3): 30–42. 10.15586/QAS.V14I3.1064
Gmoser, R., Fristedt, R., Larsson, K., Undeland, I., Taherzadeh, M.J. and Lennartsson, P.R., 2020. From stale bread and brewers spent grain to a new food source using edible filamentous fungi. Bioengineered 11(1): 582–598. 10.1080/21655979.2020.1768694
Gnanwa, J.M., Soro, L.C., Fagbohoun, J.B., Yorou, N.S. and Kouame, L.P., 2021. Assessment of minerals, vitamins, amino and fatty acids components of Pleurotus ostreatus mushrooms cultivated and sold in the village of M’Badon (Abidjan, Côte d’Ivoire). International Journal of Current Microbiology and Applied Sciences 10(09): 276–283. 10.20546/ijcmas.2021.1009.032
Hamam, M., Chinnici, G., Di Vita, G., Pappalardo, G., Pecorino, B., Maesano, G., et al., 2021. Circular economy models in agro-food systems: a review. Sustainability 13(6): 3453. 10.3390/SU13063453
Hoa, H.T., Wang, C.L. and Wang, C.H., 2015. The effects of different substrates on the growth, yield, and nutritional composition of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 43(4): 423–434. 10.5941/MYCO.2015.43.4.423
Hultberg, M., Prade, T., Bodin, H., Vidakovic, A. and Asp, H., 2018. Adding benefit to wetlands—valorization of harvested common reed through mushroom production. Science of the Total Environment 637–638: 1395–1399. 10.1016/J.SCITOTENV.2018.05.106
Hyde, K.D., Xu, J., Rapior, S., Jeewon, R., Lumyong, S., Niego, A.G.T., et al., 2019. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Diversity 97(1): 1–136. 10.1007/S13225-019-00430-9
Ibarruri, J., Cebrián, M. and Hernández, I., 2021. Valorisation of fruit and vegetable discards by fungal submerged and solid-state fermentation for alternative feed ingredients production. Journal of Environmental Management 281: 111901. 10.1016/J.JENVMAN.2020.111901
Jaeger, A., Arendt, E.K., Zannini, E. and Sahin, A.W., 2020. Brewer’s spent yeast (BSY), an underutilized brewing by-product. Fermentation 6(4): 123. 10.3390/FERMENTATION6040123
Karlović, A., Jurić, A., Ćorić, N., Habschied, K., Krstanović, V. and Mastanjević, K., 2020. By-Products in the malting and brewing industries—re-usage possibilities. Fermentation 6(3): 82. 10.3390/FERMENTATION6030082
Keong, C.Y., 2015. Medicinal values of selected mushrooms with special reference to anti-hypercholesterolemia. In: Kumar, S.A., editor. Hypercholesterolemia. London: IntechOpen. 10.5772/59424
Koutrotsios, G., Mountzouris, K.C., Chatzipavlidis, I. and Zervakis, G.I., 2014. Bioconversion of lignocellulosic residues by Agrocybe cylindracea and Pleurotus ostreatus mushroom fungi—assessment of their effect on the final product and spent substrate properties. Food Chemistry 161: 127–135. 10.1016/j.foodchem.2014.03.121
Kumla, J., Suwannarach, N., Sujarit, K., Penkhrue, W., Kakumyan, P., Jatuwong, K., et al., 2020. Cultivation of mushrooms and their lignocellulolytic enzyme production through the utilization of agro-industrial waste. Molecules 25(12): 2811. 10.3390/molecules25122811
Linhartová, L., Michalíková, K., Šrédlová, K. and Cajthaml, T., 2020. Biodegradability of dental care antimicrobial agents chlorhexidine and octenidine by ligninolytic fungi. Molecules 25(2): 400. 10.3390/MOLECULES25020400
Liu, S., Ahmed, S., Zhang, C., Liu, T., Shao, C. and Fang, Y., 2020. Diversity and antimicrobial activity of culturable fungi associated with sea anemone Anthopleura xanthogrammica” Electronic Journal of Biotechnology 44: 41–46. 10.1016/J.EJBT.2020.01.003
Londoño-Hernandez, L., Ruiz, H.A., Ramírez, T.C., Ascacio, J.A., Rodríguez-Herrera, R. and Aguilar, C.N., 2020. Fungal detoxification of coffee pulp by solid-state fermentation. Biocatalysis and Agricultural Biotechnology 23: 101467. 10.1016/J.BCAB.2019.101467
López-Fernández, J., Benaiges, M.D. and Valero, F., 2020. Rhizopus oryzae lipase, a promising industrial enzyme: biochemical characteristics, production and biocatalytic applications. Catalysts 10(11): 1277. 10.3390/CATAL10111277
Marcus, A. and Fox, G., 2021. Fungal biovalorization of a brewing industry byproduct, brewer’s spent grain: a review. Foods 10(9): 2159. 10.3390/FOODS10092159
Meyer, V., Basenko, E.Y., Benz, J.P., Braus, G.H., Caddick, M.X., Csukai, M., et al. 2020. Growing a circular economy with fungal biotechnology: a white paper. Fungal Biology and Biotechnology 7(1): 1–23. 10.1186/S40694-020-00095-Z
Mohajeri, S., Harsej, F., Sadeghpour, M. and Nia, J.K., 2021. Integrated reverse supply chain model for food waste based on industry 4.0 revolutions: a case study of producing the household waste recycling machine. Quality Assurance and Safety of Crops & Foods 13(4): 70–83. 10.15586/QAS.V13I4.1002
Naim, L., Alsanad, M.A., Shaban, N., El Sebaaly, Z., Fayssal, S.A. and Sassine, Y.N., 2020. Production and composition of Pleurotus ostreatus cultivated on Lithovit®-Amino25 Supplemented spent substrate. AMB Express 10(1): 1–10. 10.1186/S13568-020-01124-1/TABLES/10
Pauletto, M., Elgendy, R., Ianni, A., Marone, E., Giantin, M., Grotta, L., et al. 2020. Nutrigenomic effects of long-term grape pomace supplementation in dairy cows. Animals 10(4): 714. 10.3390/ANI10040714
Pedneault, K., Angers, P., Avis, T.J., Gosselin, A. and Tweddell, R.J., 2007. Fatty acid profiles of polar and non-polar lipids of Pleurotus ostreatus and P. cornucopiae Var. “citrino-Pileatus” grown at different temperatures. Mycological Research 111(10): 1228–1234. 10.1016/j.mycres.2007.06.014
Pellegrino, R.M., Blasi, F., Angelini, P., Ianni, F., Alabed, H.B.R., Emiliani, C., et al. 2022. LC/MS Q-TOF metabolomic investigation of amino acids and dipeptides in Pleurotus ostreatus grown on different substrates. Journal of Agricultural and Food Chemistry 2022: 10371–1082. 10.1021/ACS.JAFC.2C04197/ASSET/IMAGES/LARGE/JF2C04197_0007.JPEG
{kind=link}
Puglia, D., Pezzolla, D., Gigliotti, G., Torre, L., Bartucca, M.L. and Del Buono, D., 2021. The opportunity of valorizing agricultural waste, through its conversion into biostimulants, biofertilizers, and biopolymers. Sustainability 13(5): 2710. 10.3390/SU13052710
Rachwał, K., Waśko, A., Gustaw, K. and Polak-Berecka, M., 2020. Utilization of brewery wastes in food industry. PeerJ 8: e9427. 10.7717/peerj.9427
Radenkovs, V., Juhnevica-Radenkova, K., Górnaś, P. and Seglina, D., 2018. Non-waste technology through the enzymatic hydrolysis of agro-industrial by-products. Trends in Food Science & Technology 77: 64–76. 10.1016/J.TIFS.2018.05.013
Reis, S.F., Coelho, E., Coimbra, M.A. and Abu-Ghannam, N., 2015. Improved efficiency of brewer’s spent grain Arabinoxylans by ultrasound-assisted extraction. Ultrasonics Sonochemistry 24: 155–164. 10.1016/J.ULTSONCH.2014.10.010
Riudavets, J., Castañé, C., Agustí, N., Del Arco, L., Diaz, I. and Castellari, M., 2020. Development and biomass composition of Ephestia kuehniella (Lepidoptera: Pyralidae), Tenebrio molitor (Coleoptera: Tenebrionidae), and Hermetia illucens (Diptera: Stratiomyidae) reared on different byproducts of the agri-food industry. Journal of Insect Science 20(4): 17. 10.1093/jisesa/ieaa085
Severini, C., Azzollini, D., Jouppila, K., Jussi, L., Derossi, A. and De Pilli, T., 2015. Effect of enzymatic and technological treatments on solubilisation of Arabinoxylans from brewer’s spent grain. Journal of Cereal Science 65: 162–66. 10.1016/j.jcs.2015.07.006
Sinanoglou, V.J., Zoumpoulakis, P., Heropoulos, G., Proestos, C., Ćirić, A., Petrovic, J., et al. 2015. Lipid and fatty acid profile of the edible fungus Laetiporus sulphurous. Antifungal and antibacterial properties. Journal of Food Science and Technology 52(6): 3264. 10.1007/S13197-014-1377-8
Sousa, D., Venâncio, A., Belo, I. and Salgado, J.M., 2018. Mediterranean agro-industrial wastes as valuable substrates for lignocellulolytic enzymes and protein production by solid-state fermentation. Journal of the Science of Food and Agriculture 98(14): 5248–5256. 10.1002/jsfa.9063
Statistica, 2021. Beer—worldwide | Statista market forecast. Available from: https://www.statista.com/outlook/10010000/100/beer/worldwide
Tartrakoon, W., Tartrakoon, T. and Kitsupee, N., 2016. Effects of the ratio of unsaturated fatty acid to saturated fatty acid on the growth performance, carcass and meat quality of finishing pigs. Animal Nutrition 2(2): 79–85. 10.1016/j.aninu.2016.03.004
Verni, M., Pontonio, E., Krona, A., Jacob, S., Pinto, D., Rinaldi, F., et al. 2020. Bioprocessing of brewers’ spent grain enhances its antioxidant activity: characterization of phenolic compounds and bioactive peptides. Frontiers in Microbiology 11: 1831. 10.3389/FMICB.2020.01831/TEXT
Wang, D., Sakoda, A. and Suzuki, M., 2001. Biological efficiency and nutritional value of Pleurotus ostreatus cultivated on spent beer grain. Bioresource Technology 78(3): 293–300. 10.1016/S0960-8524(01)00002-5
Wang, H., Kaur, G., Pensupa, N., Uisan, K., Du, C., Yang, X., et al., 2018. Textile waste valorization using submerged filamentous fungal fermentation. Process Safety and Environmental Protection 118: 143–151. 10.1016/J.PSEP.2018.06.038
Weete, J.D., Abril, M. and Blackwell, M., 2010. Phylogenetic distribution of fungal sterols. PLoS One 5(5): e10899. 10.1371/journal.pone.0010899
Wołoszyn, J., Haraf, G., Okruszek, A., Wereńska, M., Goluch, Z. and Teleszko, M., 2020. Fatty acid profiles and health lipid indices in the breast muscles of local Polish goose varieties. Poultry Science 99(2): 1216–1224. 10.1016/J.PSJ.2019.10.026
Xiros, C. and Studer, M.H., 2017. A multispecies fungal biofilm approach to enhance the celluloyltic efficiency of membrane reactors for consolidated bioprocessing of plant biomass. Frontiers in Microbiology 8: 1930. 10.3389/FMICB.2017.01930/BIBTEX
Xu, X., Lin, M., Zang, Q. and Shi, S., 2018. Solid state bioconversion of lignocellulosic residues by Inonotus obliquus for production of cellulolytic enzymes and saccharification. Bioresource Technology 247: 88–95. 10.1016/j.biortech.2017.08.192
Yang, R., Xue, L., Zhang, L., Wang, X., Qi, X., Jiang, J., et al., 2019. Phytosterol contents of edible oils and their contributions to estimated phytosterol intake in the Chinese diet. Foods 8(8): 334. 10.3390/foods8080334
Zhang, X.Y., Li, B., Huang, B.C., Wang, F.B., Zhang, Y.Q., Zhao, S.G., et al., 2022. Production, biosynthesis, and commercial applications of fatty acids from oleaginous fungi. Frontiers in Nutrition 9: 830. 10.3389/FNUT.2022.873657/BIBTEX