
Download






Research Article
Artificial intelligence-based model for evaluating the inhibition of Listeria monocytogenes, Staphylococcus aureus, and Escherichia coli in kefir matrix
Kefyalew Chirkena Bali1, Fatma Kaya Yıldırım1,2*, Beyza Hatice Ulusoy1,2
1Department of Food Hygiene and Technology, Faculty of Veterinary Medicine, Near East University, Nicosia, Cyprus;
2DESAM Research Institute, Near East University, Nicosia, Cyprus
Abstract
The present study aimed to inhibit the activity of some foodborne pathogens by probiotic lactic acid bacteria (LAB) in kefir. The antimicrobial effect of probiotic LAB was evaluated by using Artificial Intelligence (AI)-based models, Artificial Neural Network (ANN), and Adaptive Network-based Fuzzy Inference System (ANFIS). The experiment was performed on fermentation day 0, 1, and 2, and storage day 1, 3, 7, and 10 of kefir. The average inhibition results obtained for Listeria monocytogenes, Staphylococcus aureus, and Escherichia coli at training stage was 2.4 log10 CFU/g, 2.0 log10 CFU/g, and 2.4 log10 CFU/g in Artificial Neural Network, respectively, and 2.33 log10 CFU/g, 2.04 log10 CFU/g, and 2.03 log10 CFU/g in Adaptive Network-based Fuzzy Inference System, respectively. The average result obtained in the case of tested LAB was 4.9 log10 CFU/g, 4.8 log10 CFU/g, and 4.9 log10 CFU/g, respectively, in Artificial Neural Network in each organism; while similar result was observed in Adaptive Network-based Fuzzy Inference System. The results indicate that the activity of all targeted foodborne pathogens was reduced during fermentation and storage days by the potential probiotic LAB present in kefir. Based on the experiment, it was concluded that the activity of foodborne pathogens can be inhibited by probiotic LAB in kefir. In addition, it was suggested that probiotic bacteria in kefir are promising bio-controlling agents that can be used in the food industry.
Key words: Listeria monocytogenes, Staphylococcus aureus, Escherichia coli, kefir, probiotic, lactic acid bacteria
*Corresponding Author: Fatma Kaya Yıldırım, Department of Food Hygiene and Technology, Faculty of Veterinary Medicine, Near East University, Nicosia, Cyprus. Email: [email protected]
Academic Editor: Amin Mousavi Khaneghah – Faculty of Biotechnologies (BioTech), ITMO University, Saint Petersburg, Russia
Received: 22 January 2024; Accepted: 1 August 2024; Published: 22 November 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Foodborne pathogens are known to be the most common food contaminating microorganisms that cause foodborne illnesses. The occurrence of foodborne illnesses is mostly arising from contamination of fresh agricultural produce that intensify disease outbreaks globally (Yang et al., 2024a). The contamination of food by pathogens is mostly associated with inappropriate food handling during production, processing, storage, and transportation. Moreover, international trade fairs and diverse food supply chains also widely enhance the spread of foodborne diseases across borders, resulting in health and related risks to the consumers, especially people of vulnerable groups, such as pregnant women, elders, and infants. As a result, the issue of foodborne illnesses has reached the alarming stage of food safety because of global public health issue (Choi et al., 2020). The issue is not only of global health concern but also of its considerable economic burden, particularly in developing countries, because of the risk associated with the safety and quality of food products (Keba et al., 2020).
Among the common foodborne pathogens of public health and economic issue are Listeria monocytogenes, Staphylococus auerus, and coliform bacteria that cause serious foodborne infections and damage food products. L. monocytogenes is the most serious and deadly pathogen that causes foodborne infections (Rivas-Macho et al., 2024). The pathogen is ubiquitous in nature, and can live and persist in different conditions, including farm environments, food production environments, food products, food contact surfaces and utensils, as it is a biofilm-forming organism in monospecies or in conjunction with other microbes (Fagerlund et al., 2020; Gu et al., 2024; Kallipolitis et al., 2020; Kannan et al., 2020). It causes listeriosis in humans, particularly in venerable groups, such as pregnant women, elders, and people with debilitated or compromised immune system. It spreads following consumption of either contaminated or undercooked foods, pasteurized milk, soft cheese, semi-soft cheese, and cooked meat (Chen et al., 2020; Cufaoglu et al., 2021; Olaimat et al., 2021).
It has been suggested by some research findings that foods with low moisture content (foods with water activity < 0.85) are found to be a potential source of this pathogen (Ly et al., 2019; Taylor and Zhu, 2021); foods that are kept under cold conditions for more than the recommended period can enhance the chance of survival for the pathogen in the food products. Another finding has indicated that prevalence of the pathogen in dairy products is high and negatively affects the dairy industry (El Hag et al., 2021). For the most part, since the pathogen is capable to contaminate food products along the food chain from production stages to consumption, a large number of recalls are observed in the food processing industry (Duze et al., 2021). Besides the infections associated with consumption of food products, the pathogen is responsible for other illnesses, such as meningitis (meningoencephalitis), central nervous system infection, silent diabetes mellitus, and other complications in immune-compromised groups (Steinbrecher et al., 2023).
Another foodborne pathogen is Staphylococcus aureus. It is well characterized by its colonization on the skin and the upper respiratory tract in humans under normal conditions (Flora et al., 2019). The pathogen causes staphylococcal food poisoning by the consumption of staphylococcal enterotoxins produced in foods and subsequent exposure of consumers to various health problems, resulting in huge economic losses to the food industry (Farha et al., 2020). The level of poisoning depends on the secretion of multiple toxic proteins, especially if they are produced >105 CFU/g, representing pathogenic toxins (Zhao et al., 2020). The staphylococcal enterotoxins are often reported in dairy milk products, and are observed as the major cause of infections associated with food poisoning in humans and as mastitis in animals (Zhao et al., 2021). The pathogen is opportunistic as it can survive in the food processing environments because of its tolerance to desiccation and other stress conditions (Wang et al., 2023). Similar to L. monocytogenes, S. aureus is a ubiquitous foodborne pathogen and biofilm-forming bacteria that persistently attach to food contact surfaces during food production, thereby contaminating food products (Rubab et al., 2018; Titouche et al., 2019). In dairy farms, the pathogen is most commonly known to cause mastitis in dairy cattle, leading to potential risk to public health as well as product loss and economic crises to the dairy industry.
Coliforms are also among the foodborne bacteria that are abundantly present in the environment as well as in the fecal matter of humans and warm-blooded animals. However, the presence of total coliform bacteria in food products interfere the quality of products, leading to the loss of physicochemical characteristics, particularly that of dairy products (Selover et al., 2021). Among the group of coliforms, the presence of toxin-producing E. coli in dairy products (raw and processed milk) has become a public health concern due to its ability to produce infective shigatoxin (Rosario et al., 2021). Thus, food products, including dairy products, are among the various food matrices where pathogens are detected and lead to the majority of food borne illnesses in humans (Fallah et al., 2021; Tian et al., 2022).
Several mechanisms and approaches are established to mitigate the activity and spread of foodborne pathogens. Biological controlling approach is considered as an effective way of combating the potential risk of pathogens to ensure food safety and quality. Probiotic bacteria are one of the biological agents that a number of scientific researchers have addressed for their antimicrobial effects on foodborne pathogens (Abdelhamid and El-Dougdoug, 2020; Jara et al., 2020; Kaya and Simsek, 2019; Rajabi et al., 2020). These probiotic bacteria produce different metabolites, such as bacteriocins, organic acids, and other components that inhibit the growth of pathogenic microorganisms. Meanwhile, some research findings have reported that some probiotics have traits (genes) that might be transferred to their hosts. In light of this, some research reports identified that probiotic bacteria, such as Bacillus species, have virulence genes/factors, such as nheABC, entFM enterotoxin genes, hbIA, hbIC, hbID, and cytK (Anokyewaa et al., 2021) as well as nheABC, hblCDA, and cytotoxin cytK2 (Deng et al., 2021) that enable them constrain the activity of microbes and resist antibiotics. In another investigation, it was identified that isolates of some probiotic Enterococcus faecium from Tulum cheese traditionally produced in Turkey exhibited their antibacterial activity against L. monocytogenes ATCC 7644 and S. aureus ATCC 25923 because of the presence of virulence gene determinants, such as asal, gelE, CylA, cylLL,s, cylM, Hylemf, and gelE, detected in the probiotic bacteria (Özkan et al., 2021). The majority of these probiotics, particularly LAB, are naturally present in fermented dairy and other food products, and these are considered as desirable and beneficial microorganisms (Klimko et al., 2020). Kefir is a valuable probiotic dairy product; many researchers studied bacteriostatic and bactericidal effects against bacterial pathogens as well as preventive effect on spoilage fungi (González-Orozco et al., 2022; Gut et al., 2022; John and Deeseenthum, 2015; Rosa et al., 2017; Ulusoy et al., 2007).
Regarding technological requirements, these bacteria are suitable for addition to food products because of their viability and efficacy as well as their resistance to several processing conditions employed in the food industry (Soares et al., 2023). Thus, probiotics are considered as alternative antimicrobial agents when incorporated into food products to antagonize the activity of foodborne pathogens (Ağagündüz et al., 2022; Khaneghah et al., 2020; Vahdat et al., 2024). In the present study, kefir was used as a product that contains probiotic microorganisms. It is a traditionally fermented dairy product produced by the action of bacteria and yeast existing in symbiotic association with kefir grains, and characterized by probiotic properties and antimicrobial activity (Gökmen et al., 2022; González-Orozco et al., 2022; Yilmaz et al., 2022). Limited research findings exist regarding the inhibition of targeted foodborne pathogens by using probiotic LAB in kefir milk. Therefore, the aim of this research was to inhibit the activity and growth ability of L. monocytogenes, S. aureus, and coliform bacteria, more specifically E. coli bacteria, by using probiotic LAB, naturally present in kefir milk. Feasible benefits of inhibiting these foodborne pathogens from the perspective of food safety are also identified. Rapid development of Artificial Intelligence (AI) provides significant benefits for research studies, especially for finding the way and obtaining the hypothesis of the research by predicting. In this research supporting help of AI was used for our hypothesis.
Materials and Methods
Activation of pathogenic bacterial suspensions
Pure cultures of Listeria monocytogenes (ATCC 19111) and Staphylococcus aureus (ATCC 19212) were obtained from Ankara University, Turkey. Activation of pathogenic bacteria was performed on nutrient agar media through the streaking technique, or alternatively, about 1 mL of bacterial suspension from each pathogen was suspended in 9 mL of Maximum Recovery Dilution (MRD) (Lab M Limited, Lancashire, United Kingdom). The plates containing bacterial inoculum were incubated at 37°C for 24–48 h. A fecal sample was collected for the cultivation of coliform bacteria, which was swabbed on Violet Red Bile (VRB) agar (Merck kGaA, Darmstadt, Germany) and incubated at 37°C for 24–48 h. The suspension of both Listeria monocytogenes and Staphylococcus aureus pathogens was separately adjusted to 0.5 McFarland turbidity standards to approximate bacterial concentration to 106 CFU/mL as described by Prezzi et al. (2020).
Kefir grains and preparing kefir milk
Kefir grains were donated by the Department of Food Hygiene and Technology of Near East University, Nicosia, Cyprus. The grains were maintained and activated in full-fat ultra high temperature (UHT) milk at 25°C for 18-h fermentation period. The grains were strained through a sterile sieve and washed with sterile normal saline solution to remove curdle part of the milk as described by Angelidis et al. (2020) and fermented kefir milk was obtained by following the process of straining kefir grains.
Experimental contamination of kefir
Four sterile glass jars (beakers) were used to prepare experimental groups. For each group, 5 g of kefir grains was weighed aseptically and mixed with 50 mL of UHT milk. About 0.1 mL of bacterial suspension of L. monocytogenes, S. aureus, and coliforms was added to each jar. Inoculation was not done in one of the jars, which was considered as the control group. All the jars containing homogenized solutions were kept in an incubator at 30°C for 2 days of fermentation process as described by Dimitreli and Antoniou (2011) and Gökmen et al. (2022). After complete removal of grains, the kefir milk in each jar was refrigerated at 4°C for microbiological analysis.
Microbiological analysis
The results of microbiological analysis of LAB in L. monocytogenes, S. aureus, coliforms/E. coli are described in Table 1.
Table 1. Medium, incubation conditions, and analytical references of methods used for microbiological analysis of LAB.
| Micro-organisms | Analytical reference method |
Media name | Incubation conditions | ||
|---|---|---|---|---|---|
| Incubation temperature | Incubation period | O2 requirement | |||
| Staphylococci Staphylococcus aureus | ISO 6888-1:1999 + A1:2003 | Baird Parker Medium Agar (LAB 085) + Egg Yolk Tellurite Emulsion (X 085) |
35–37ºC | 24 ± 2 h | Aerobic |
| Brain Heart Infusion Broth (LAB 049) | Confirmation for Staphylococcus aureus | ||||
| Coliform bacteria | ISO 4832:2006 | Violet Red Bile Glucose Agar (LAB 031) | 30–37ºC | 24 ± 2 h | Microaerophilic |
| Brilliant Green Bile Broth (LAB051) | Confirmation | ||||
| Listeria monocytogenes | ISO 11290-1: 1996 + A1:2004 | Half Fraser Broth Base (LAB 164) | 30ºC | 24 ± 2 h | Aerobic |
| Fraser Broth Base (LAB 164) | 37ºC | 24 h | |||
| Palcam Agar (LAB 148) | 37ºC | 24 ± 3 h | |||
| Tryptone Soya Yeast Extract Broth (LAB004) | Confirmation for Listeria spp. | ||||
| Sheep Blood Agar (LAB028) | Confirmation for L. monocytogenes | ||||
| Lactic acid bacteria | ISO 15214:1998 | MRS Agar (LAB223) | 30ºC | 2–3 days | Anaerobic |
Lactic acid bacteria
Serial dilutions were prepared with minimum required dilution (MRD) in the ratio of 1:9 in aseptic conditions for microbially contaminated solutions. About 0.1 mL of solution representing the targeted microbe was spread on selective agar plates. The microbiological analysis of LAB was carried out aseptically by pouring onto the pre-prepared De Man–Rogosa–Sharpe (MRS) agar medium (Merck kGaA) under anaerobic conditions. The plates were incubated at 37°C for 48 h. Microbial enumeration was conducted by using colony counting machine after completion of incubation. The enumerated LAB was analyzed and compared with the control group.
Pathogens
From the serial dilutions prepared for microbial contaminated solutions, about 0.1 mL of solution of each pathogen was added into the designated growth media and spread on the surfaces of agar plates. L. monocytogenes was grown on polymyxin acriflavin lithium-chloride ceftazidime esculin mannitol (PALCAM) agar (Lab M Limited, Lancashire, UK), while S. aureus and coliform/E. coli bacteria were allowed to grow on Baird–Parker agar supplemented with egg yolk tellurite emulsion (20%) (Merck kGaA) and VRB agar (Merck kGaA, 64271 Darmstadt, EMD Millipore Corporation, Germany), respectively. The procedure was carried out during fermentation and storage (post-fermentation) of dairy kefir. The plates were kept in an incubator at 37°C for 24–48 h as described by Angelidis et al. (2020) with some modifications. Following the removal of colony counting plates from the incubator, bacterial colonies were enumerated on the surface of plates. The enumeration of coliforms was done by pour plating method by cultivating bacteria onto VRB agar in anaerobic conditions. The identification of E. coli was done by incubating VRB agar plate containing coliform bacteria at a temperature of 44°C for 24–48 h.
Data analysis by AI
The experimental analysis of the present study was conducted in triplicate. AI data-based approaches (artificial neural network [ANN] and adaptive network-based fuzzy inference system [ANFIS] models) were applied to analyze inhibition of L. monocytogenes, S. aureus, and E. coli foodborne pathogenic bacteria by using probiotic LAB present in dairy kefir milk.
Results
Listeria monocytogenes
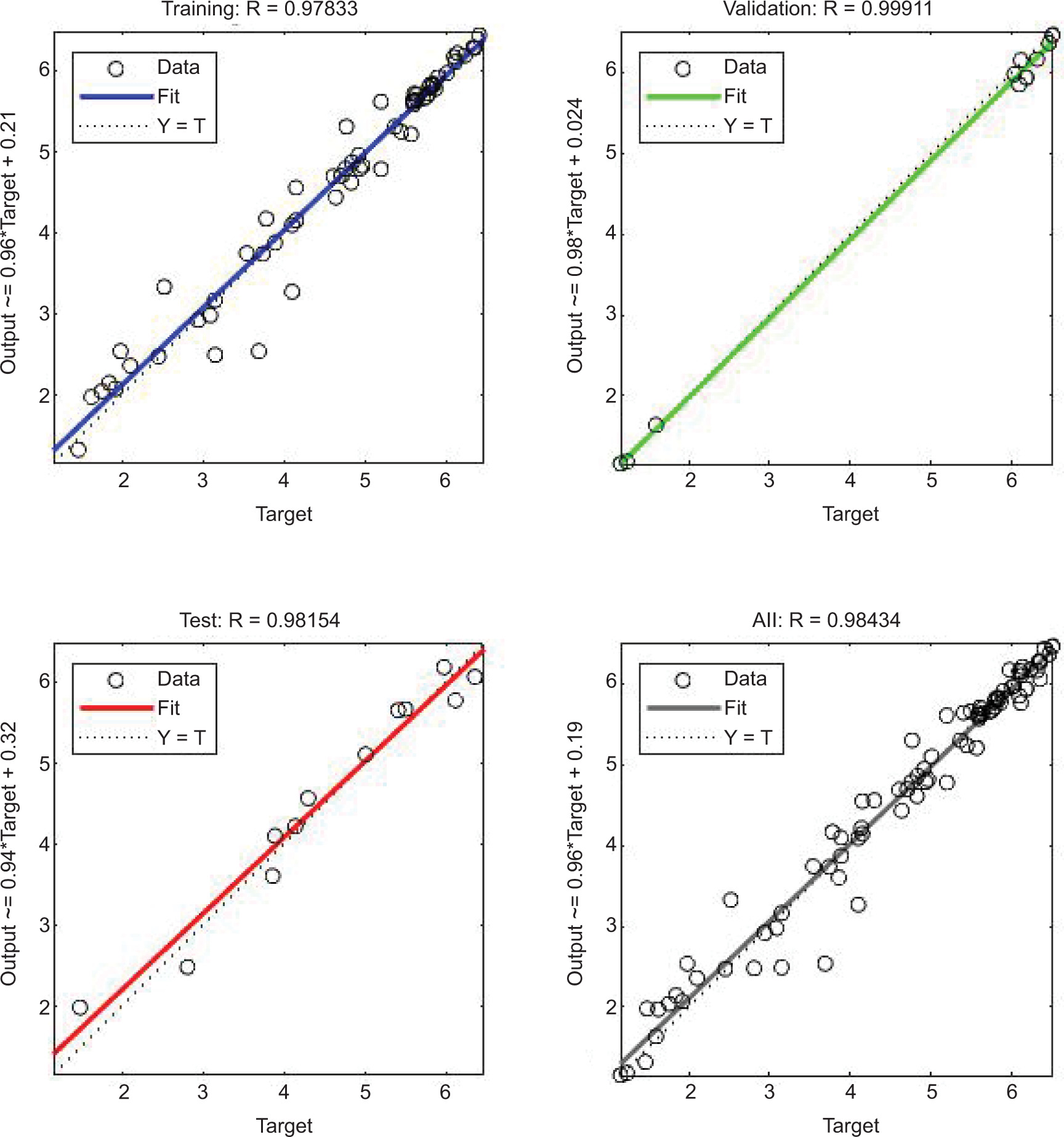
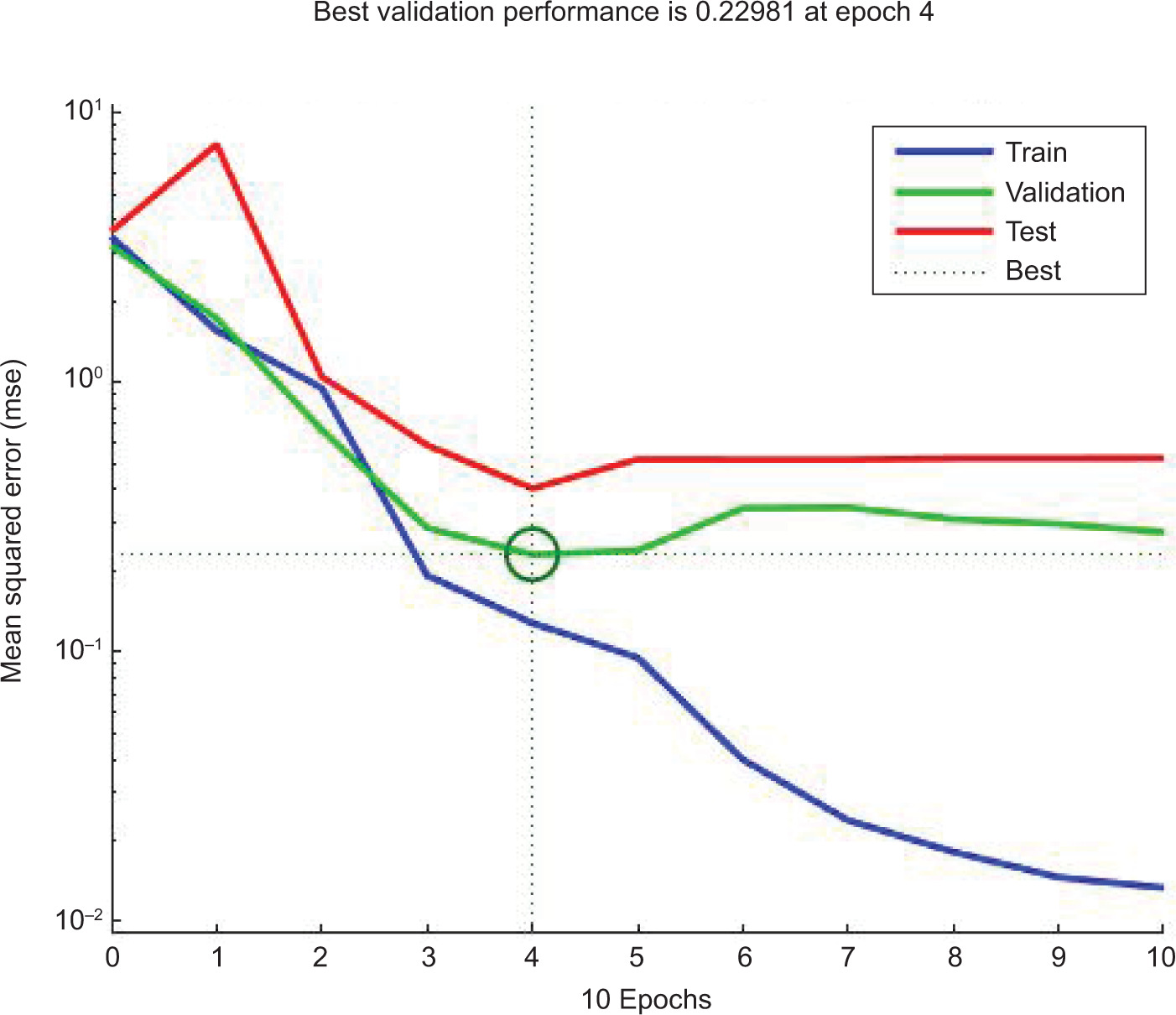
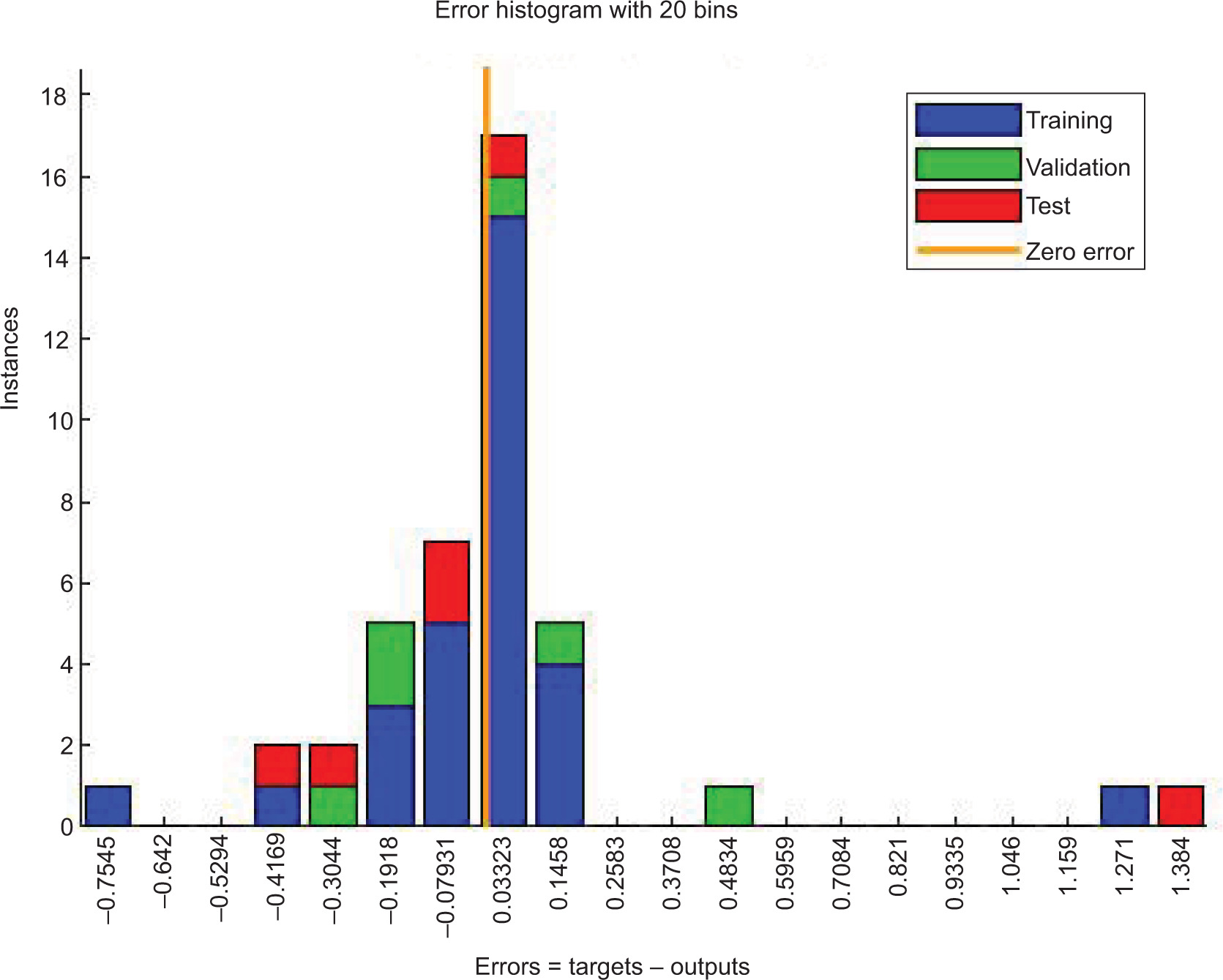
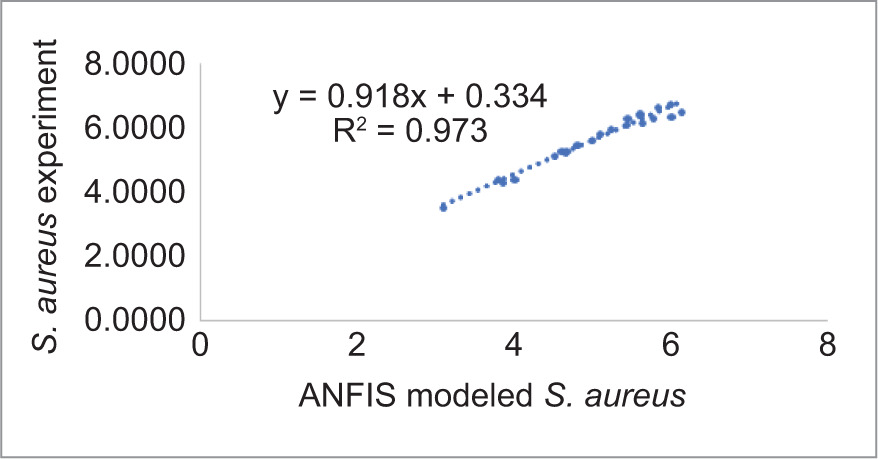
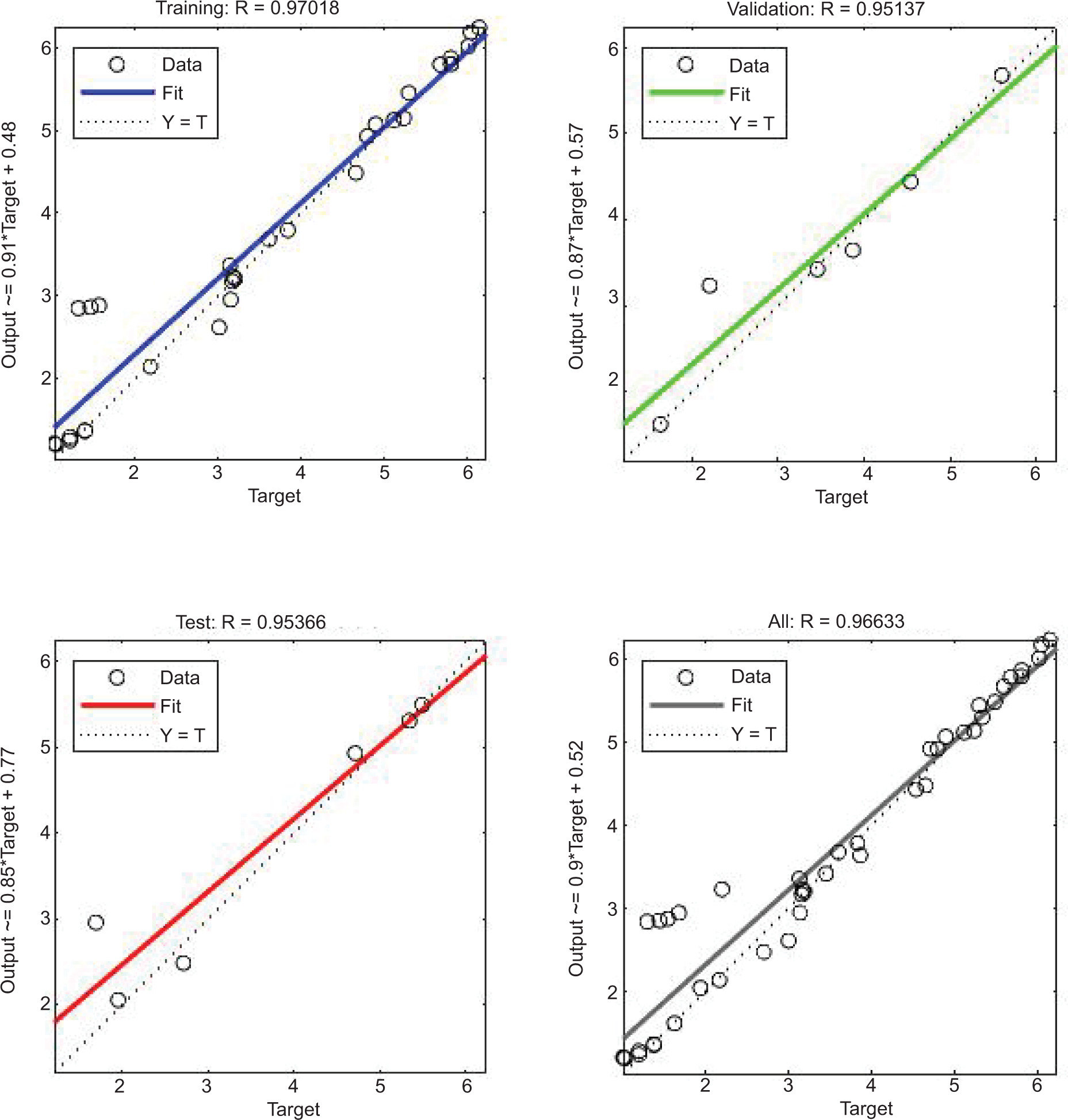
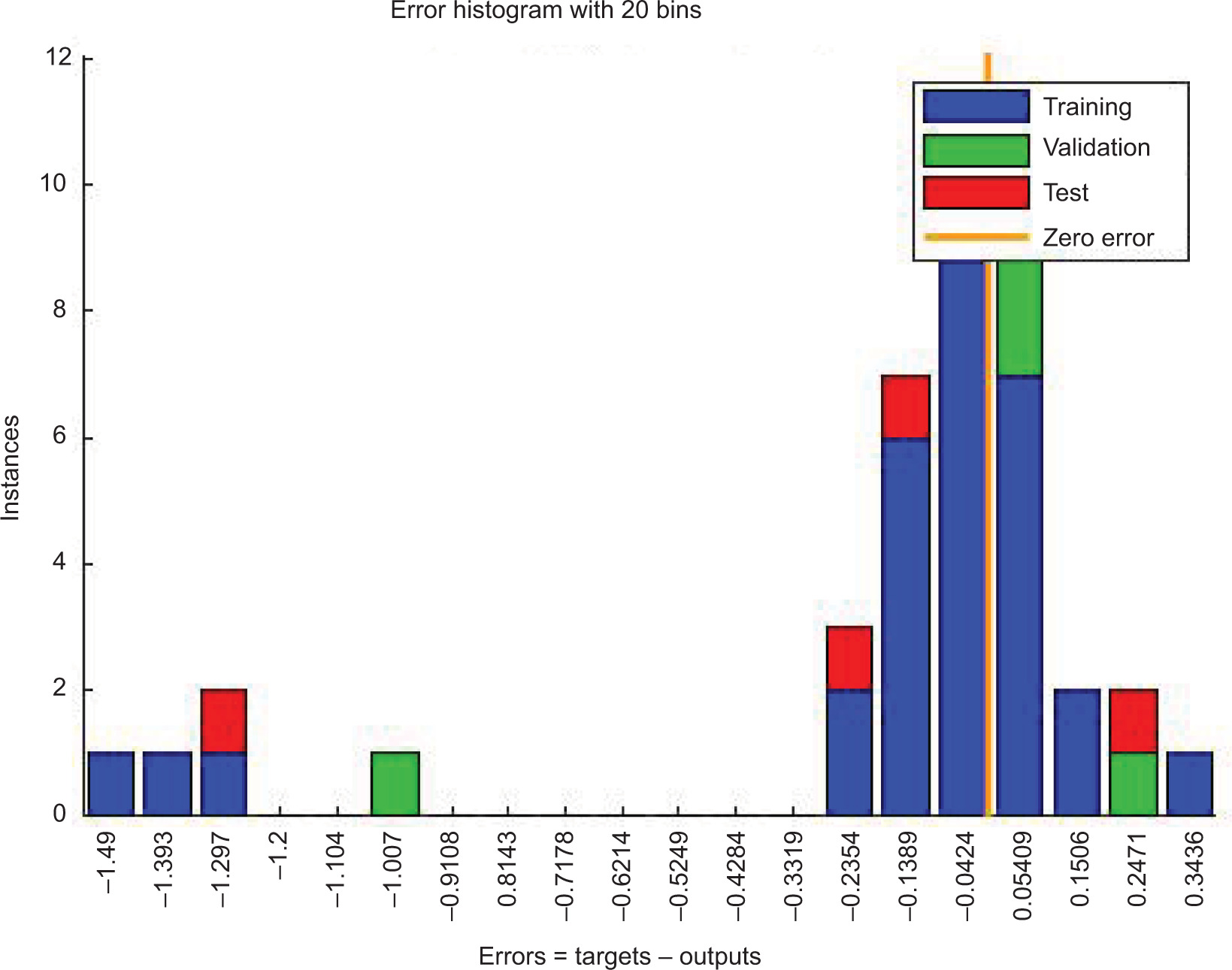
The ANN model analyzed the inhibition of L. monocytogenes by using probiotic dairy kefir, and the inhibition was evaluated at a specified time. The average of obtained results for LAB control, tested LAB, and L. monocytogenes were 5.23, 4.95, and 2.41 log10 CFU/g, respectively, at training stage whereas at testing stage, the recorded values were 5.80, 5.38, and 2.04 log10 CFU/g, respectively. As described in Table 2, reduction in the number of pathogens was observed on SD1, SD3, SD7, and SD10 whereas the number of tested LAB increased contrarily on stated days. In addition, the obtained result for the reduction of number of L. monocytogenes was also supported by regression analysis at training, validation, and testing stages, with regression (R) = 0.9783, 0.9991, 0.9815, respectively (Figure 1). Furthermore, the ANN model revealed the inhibitory activity of LAB against the pathogen with the best validation performance of 0.2298 at epoch 4 (Figure 2) and error of the model as 0.02395 as shown in Figure 3. Similarly, the ANFIS model also simulated the overall modelling of inhibition of L. monocytogenes using probiotic dairy kefir. In this model, the average number of the bacteria was 5.23, 4.93, and 2.33 log10 CFU/g for LAB control, tested LAB, and L. monocytogenes, respectively, for the training stage, and 5.65, 5.29, and 2.45 log10 CFU/g, respectively, at the testing stage (Table 3). The number of pathogens reduced starting from fermentation day 2 (FD2) to storage day 10 (SD10) whereas the number of LAB increased from FD0 to SD10, unlike the number of pathogens.
Table 2. ANN model inhibition of L. monocytogenes by probiotic dairy kefir (in log10 CFU/g).
| Days | LAB control | Tested LAB | L. monocytogenes |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.1554 | 2.92988 |
| FD1 | 4.1 | 3.81108 | 3.16429 |
| FD2 | 4.96 | 4.70434 | 3.9069 |
| SD1 | 5.1 | 4.95914 | 2.62325 |
| SD3 | 5.2 | 5.03216 | 2.25576 |
| SD7 | 5.29 | 5.04857 | 2.17457 |
| SD10 | 5.4 | 5.0539 | 2.15349 |
| FD0 | 4.2 | 4.09884 | 3.72184 |
| FD1 | 5.1 | 4.95914 | 2.62325 |
| FD2 | 5.86 | 5.25682 | 2.02536 |
| SD1 | 5.98 | 5.41074 | 1.91552 |
| SD3 | 6.11 | 5.52634 | 1.77089 |
| SD7 | 6.34 | 6.01235 | 1.29927 |
| SD10 | 6.43 | 6.25392 | 1.18377 |
| Average | 5.2350 | 4.9488 | 2.4106 |
| Testing stage | |||
| FD0 | 4.18 | 4.03903 | 3.6057 |
| FD1 | 4.97 | 4.72179 | 3.81903 |
| FD2 | 5.78 | 5.16275 | 2.08419 |
| SD1 | 5.88 | 5.28384 | 2.00786 |
| SD3 | 6.1 | 5.51774 | 1.78513 |
| SD7 | 6.23 | 5.69512 | 1.53622 |
| SD10 | 6.41 | 6.20991 | 1.20264 |
| Average | 5.8081 | 5.3876 | 2.0497 |
Table 3. ANFIS model inhibition of L. monocytogenes by probiotic dairy kefir (in log10 CFU/g).
| Days | LAB control | Tested LAB | L. monocytogenes |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.15 | 3.223 |
| FD1 | 4.1 | 3.9478 | 3.2745 |
| FD2 | 4.96 | 4.6715 | 2.7313 |
| SD1 | 5.1 | 4.816 | 2.608 |
| SD3 | 5.2 | 4.8571 | 2.6016 |
| SD7 | 5.29 | 4.9144 | 1.8703 |
| SD10 | 5.4 | 5.0067 | 1.3871 |
| FD0 | 4.2 | 4.0791 | 3.3261 |
| FD1 | 5.1 | 4.8143 | 2.7302 |
| FD2 | 5.86 | 5.5105 | 2.8393 |
| SD1 | 5.98 | 5.6223 | 1.6358 |
| SD3 | 6.11 | 5.7867 | 1.6372 |
| SD7 | 6.34 | 5.8936 | 1.2656 |
| SD10 | 6.43 | 5.9725 | 1.5433 |
| Average | 5.2350 | 4.9316 | 2.3338 |
| Testing stage | |||
| FD0 | 4.18 | 4.1123 | 3.1567 |
| FD1 | 4.97 | 4.611 | 2.8691 |
| FD2 | 5.78 | 5.4321 | 1.6415 |
| SD1 | 5.88 | 5.5083 | 1.8067 |
| SD3 | 6.1 | 5.6885 | 1.9563 |
| SD7 | 6.23 | 5.804 | 2.7218 |
| SD10 | 6.41 | 5.8989 | 3.0079 |
| Average | 5.6500 | 5.2936 | 2.4514 |
Figure 1. Regression graph of ANN model prediction for the inhibition of L. monocytogenes (in log10 CFU/g) using probiotic dairy product kefir.
Figure 2. Plot performance graph of ANN model prediction for the inhibition of L. monocytogenes (in log10 CFU/g) using probiotic dairy product kefir.
Figure 3. Histogram of ANN model prediction for the inhibition of L. monocytogenes (in log10 CFU/g) using probiotic dairy product kefir.
Staphylococcus aureus
Similar to L. monocytogenes, the ANN and ANFIS models also analyzed the inhibition of S. aureus by using probiotic dairy kefir during fermentation and storage days. The models evaluated at the specified time interval as displayed in Tables 4 and 5, respectively. On average, S. aureus was simulated by ANN model in relation to LAB control (5.23 log10 CFU/g) and tested LAB (4.89 log10 CFU/g), arriving at an average reduction of 2.04 log10 CFU/g. The inhibition of S. aureus by probiotic LAB in dairy kefir was also supported by regression analysis at training, validation, and testing stages, with R = 0.9842, 0.9905, 0.8873, respectively, as shown in Figure 5, and with the best validation performance of 0.071812 at epoch 21 (Figure 6). The inhibition of S. aureus by probiotic LAB was also analyzed by ANFIS model. As shown in Table 5, reduction in number of S. aureus was observed from FD2 to SD10 at the training stage, while the number of LAB increased during the above-stated days.
Figure 4. Scatter plots of ANFIS model prediction for the inhibition of L. monocytogenes using probiotic dairy product kefir.
Figure 5. The regression graph of ANN model prediction for the inhibition of S. aureus (in log10 CFU/g) using probiotic dairy product kefir.
Figure 6. Plot performance graph of ANN model prediction for the inhibition of S. aureus (in log10 CFU/g) using probiotic dairy product kefir.
Table 4. ANN model inhibition of S. aureus using probiotic dairy product kefir (in log10 CFU/g).
| Days | LAB control | Tested LAB | S. aureus |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.099997 | 2.729985 |
| FD1 | 4.1 | 3.860021 | 2.890063 |
| FD2 | 4.96 | 4.574421 | 2.371017 |
| SD1 | 5.1 | 4.601372 | 2.29078 |
| SD3 | 5.2 | 4.870955 | 1.455232 |
| SD7 | 5.29 | 5.011625 | 1.042168 |
| SD10 | 5.4 | 5.051125 | 1.099865 |
| FD0 | 4.2 | 4.051899 | 3.792911 |
| FD1 | 5.1 | 4.601372 | 2.29078 |
| FD2 | 5.86 | 5.495737 | 2.373361 |
| SD1 | 5.98 | 5.599042 | 2.122545 |
| SD3 | 6.11 | 5.728212 | 1.802943 |
| SD7 | 6.34 | 5.947762 | 1.258809 |
| SD10 | 6.43 | 6.015005 | 1.092137 |
| Average | 5.2350 | 4.8935 | 2.0438 |
| Testing stage | |||
| FD0 | 4.18 | 4.010105 | 3.69915 |
| FD1 | 4.97 | 4.574777 | 2.370089 |
| FD2 | 5.78 | 5.437892 | 2.496509 |
| SD1 | 5.88 | 5.511534 | 2.336036 |
| SD3 | 6.1 | 5.717981 | 1.828292 |
| SD7 | 6.23 | 5.84888 | 1.503898 |
| SD10 | 6.41 | 6.001234 | 1.12627 |
| Average | 5.8081 | 5.4447 | 2.0041 |
Table 5. ANFIS model inhibition of S. aureus by probiotic dairy kefir (in log10 CFU/g).
| Days | LAB control | Test LAB | S. aureus |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.1 | 2.73 |
| FD1 | 4.1 | 3.7966 | 3.1944 |
| FD2 | 4.96 | 4.5281 | 2.3997 |
| SD1 | 5.1 | 4.6312 | 2.2107 |
| SD3 | 5.2 | 4.8051 | 1.1803 |
| SD7 | 5.29 | 4.951 | 1.1873 |
| SD10 | 5.4 | 5.1009 | 1.2163 |
| FD0 | 4.2 | 3.873 | 3.081 |
| FD1 | 5.1 | 4.6364 | 2.6065 |
| FD2 | 5.86 | 5.2431 | 2.9766 |
| SD1 | 5.98 | 5.5245 | 1.3546 |
| SD3 | 6.11 | 5.6451 | 1.6865 |
| SD7 | 6.34 | 5.854 | 1.3672 |
| SD10 | 6.43 | 5.9209 | 1.469 |
| Average | 5.2350 | 4.8293 | 2.0472 |
| Testing stage | |||
| FD0 | 4.18 | 3.8731 | 3.7969 |
| FD1 | 4.97 | 4.6098 | 3.9406 |
| FD2 | 5.78 | 5.3633 | 2.1166 |
| SD1 | 5.88 | 5.4137 | 1.6937 |
| SD3 | 6.1 | 5.5426 | 1.1974 |
| SD7 | 6.23 | 5.5781 | 1.1467 |
| SD10 | 6.41 | 5.7215 | 1.0504 |
| Average | 5.6500 | 5.1574 | 2.134 |
Escherichia coli
The inhibition of E. coli by probiotic LAB present in dairy kefir was analyzed using ANN model. The results showed reduction in the number of E. coli from FD2 to SD10 whereas an increment in the number of LAB was observed from FD0 to SD10 (Table 6) at the training stage. Likewise, reduction in the number of E. coli was observed on days FD0–SD10 at the testing stage. The average result obtained for LAB control, tested LAB, and E. coli was 5.23, 4.96, and 2.46 log10 CFU/g, respectively, at the training stage whereas it was 5.81, 5.46, and 1.93 log10 CFU/g, respectively, at the testing stage as displayed in Table 6. In addition, the inhibition of the targeted pathogen was also braced by regression analysis, with R = 0.9702, 0.9514 and 0.9537 at training, validation, and testing stages, respectively (Figure 9). The best validation performance for the inhibition of the pathogen was obtained at 0.18637 as shown in Figure 10.
Figure 7. Histogram of ANN model prediction for the inhibition of S. aureus (in log10 CFU/g) using probiotic dairy product kefir.
Figure 8. Scatter plots of ANFIS model prediction for the inhibition of S. aureus (in log10 CFU/g) using probiotic dairy product kefir.
Figure 9. The regression graph of ANN model prediction for the inhibition of E. coli (in log10 CFU/g) using probiotic dairy product kefir.
Figure 10. Plot performance graph of ANN model prediction for the inhibition of E. coli (in log10 CFU/g) using probiotic dairy product kefir.
Figure 11. Histogram of ANN model prediction for the inhibition of E. coli (in log10 CFU/g) using probiotic dairy product kefir.
Figure 12. Scatter plots of ANFIS model prediction for the inhibition of E. coli (in log10 CFU/g) using probiotic dairy product kefir.
Table 6. ANN model inhibition of E. coli using probiotic dairy product kefir (in log10 CFU/g).
| Days | LAB control | Tested LAB | E. coli |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.22781 | 2.61819 |
| FD1 | 4.1 | 3.79129 | 3.17393 |
| FD2 | 4.96 | 4.43822 | 3.23498 |
| SD1 | 5.1 | 4.92613 | 2.95347 |
| SD3 | 5.2 | 5.07217 | 2.88304 |
| SD7 | 5.29 | 5.12174 | 2.85996 |
| SD10 | 5.4 | 5.14246 | 2.84803 |
| FD0 | 4.2 | 3.64604 | 3.42336 |
| FD1 | 5.1 | 4.92613 | 2.95347 |
| FD2 | 5.86 | 5.44944 | 2.14581 |
| SD1 | 5.98 | 5.67452 | 1.62552 |
| SD3 | 6.11 | 5.79773 | 1.36348 |
| SD7 | 6.34 | 6.01353 | 1.25022 |
| SD10 | 6.43 | 6.23813 | 1.20511 |
| Average | 5.2350 | 4.9618 | 2.4670 |
| Testing stage | |||
| FD0 | 4.18 | 3.68151 | 3.36319 |
| FD1 | 4.97 | 4.48445 | 3.20556 |
| FD2 | 5.78 | 5.30451 | 2.48313 |
| SD1 | 5.88 | 5.49074 | 2.04982 |
| SD3 | 6.1 | 5.79159 | 1.37418 |
| SD7 | 6.23 | 5.87139 | 1.29047 |
| SD10 | 6.41 | 6.18071 | 1.21626 |
| Average | 5.8081 | 5.4569 | 1.9261 |
Similarly, the inhibition of E. coli by biological means, namely probiotic LAB naturally present in dairy kefir milk and used in the present study, was analyzed using ANFIS model. The model simulated with a complete inhibition of E. coli at the training stage with an average number of LAB in control (5.23 log10 CFU/g), tested LAB (4.93 log10 CFU/g), and E. coli (2.03 log10 CFU/g) as shown in Table 7. Likewise, the average value of LAB control, tested LAB, and E. coli obtained at the testing stage were 5.65, 5.25, and 1.34 log10 CFU/g, respectively. For this model, the inhibition of the activity of E. coli was observed during fermentation and storage days of kefir milk, which was confirmed by the reduced number of the pathogen from FD2 to SD10.
Table 7. ANFIS model inhibition of E. coli using probiotic dairy kefir (in log10 CFU/g).
| Days | LAB control | Tested LAB | E. coli |
|---|---|---|---|
| Training stage | |||
| FD0 | 3.22 | 3.1800 | 3.0100 |
| FD1 | 4.1 | 3.8091 | 3.3488 |
| FD2 | 4.96 | 4.6054 | 2.5160 |
| SD1 | 5.1 | 4.7106 | 1.9043 |
| SD3 | 5.2 | 4.9028 | 1.4241 |
| SD7 | 5.29 | 5.0724 | 1.4142 |
| SD10 | 5.4 | 5.2434 | 1.4378 |
| FD0 | 4.2 | 3.9059 | 3.2552 |
| FD1 | 5.1 | 4.7187 | 2.7691 |
| FD2 | 5.86 | 5.3117 | 2.2918 |
| SD1 | 5.98 | 5.6904 | 1.2978 |
| SD3 | 6.11 | 5.8206 | 1.3768 |
| SD7 | 6.34 | 6.0181 | 1.2115 |
| SD10 | 6.43 | 6.0909 | 1.2226 |
| Average | 5.2350 | 4.9343 | 2.0343 |
| Testing stage | |||
| FD0 | 4.18 | 3.9910 | 2.1385 |
| FD1 | 4.97 | 4.6777 | 1.4653 |
| FD2 | 5.78 | 5.5236 | 1.3392 |
| SD1 | 5.88 | 5.5680 | 1.2989 |
| SD3 | 6.1 | 5.6617 | 1.1773 |
| SD7 | 6.23 | 5.6334 | 1.0062 |
| SD10 | 6.41 | 5.7466 | 1.0029 |
| Average | 5.6500 | 5.2574 | 1.3469 |
Discussion
Biological approach for controlling the activity of foodborne pathogens has attracted the attention researchers. The present study was conducted to investigatr the activity of targeted foodborne pathogens, namely L. monocytogens, S. aureus, and coliform/E. coli by using biological controlling approach, which include probiotic LAB present naturally in dairy kefir milk. Evidence showed that various probiotic products were used as alternative controlling means to overcome the risks associated with the effect of pathogens present the foods (Abdelhamid and El-Dougdoug, 2020; Hossain et al., 2020; Kouhi et al., 2022; Martín et al., 2022; Muñoz et al., 2019). More specifically, the report of Prezzi et al. (2020), who evaluated the effect of Lactobacillus rhamnosus GG on the growth of two foodborne pathogens (S. aureus and L. monocytogenes) by inoculating the surface of Minas Frescal cheese, indicated that the addition of Lactobacillus rhamnosus in the cheese was found to be responsible for the inhibition of L. monocytogenes.
Probiotic LAB is naturally present in different types of fermented dairy products and other food matrices, providing health benefits and microbial balance in the gastrointestinal tract of consumers (Colombo et al., 2018; Kefyalew et al., 2021). Because of the presence of different metabolites, such as organic acids, carbon dioxide, hydrogen peroxide, and bacteriocin (nisin), these beneficial microorganisms are able to inhibit the activity of foodborne pathogens. Therefore, the agricultural sector and food processing plants use beneficial microorganisms as alternative bio-control agents to mitigate the potential risk of foodborne pathogenic bacteria and their biofilms (Abdelhamid and El-Dougdoug, 2020; Kaya and Simsek, 2019).
In the present study, the inhibition of these targeted pathogens was evaluated by using ANN and ANFIS models during fermentation and storage days of dairy kefir. The activity of all selected foodborne pathogens was found to be inhibited by probiotic LAB present in dairy kefir, particularly during storage periods of kefir milk. Different research findings suggested that potential probiotic LAB isolated from various dairy products possess antimicrobial effects on foodborne pathogens, including those discussed in the present study (Mkadem et al., 2023; Wu et al., 2022; Yang et al., 2024a). For instance, the inhibitory activity of certain strains of LAB, such as Lactobacillus rhamnosus (Kamal et al., 2018), Lactobacillus paracasei (Shahverdi et al., 2023), Pediococcus pentosaceus, Pediococcus acidilactici, Enterococcus faecium, Enterococcus faecalis, and Limosilactobacillus fermentum (Roldán-Pérez et al., 2023), on different foodborne pathogens, such as E. coli O157:H7, S. aureus, Yersinia enterocolitica, Salmonella enterica serovar Typhimurium, and other strains identified as Lactobacillus plantarum strains 4–10 (Yang et al., 2024b), is taken as supportive findings for the present study.
Similarly, the findings of Mulaw et al. (2019) on the inhibitory activity of probiotic LAB against the growth/impact of S. aureus ATCC 25923, L. monocytogenes, and E. coli ATCC 25922 are evidences regarding the antimicrobial activity of LAB. More such types of probiotic bacteria are required to incorporate into foods as well as for developing functional foods. In vitro investigations revealed that LAB that survived in different bile concentrations (0.3% and 0.8%) (Paongphan et al., 2023) and was tolerant to acid (Klimko et al., 2020) exhibited antagonizing effect on the growth of foodborne pathogens. In addition to their inhibitory activity, probiotics are supplemented into foods to contribute to their organoleptic characteristics as well as to extend their shelf life during fermentation processes. Probiotics implement this function by maintaining their viability and efficacy. In the present study, the growth and activity of L. monocytogenes were tested by LAB present in dairy kefir from FD0 to FD2 and SD1 to SD10 by analyzing using both ANN and ANFIS models. The count of pathogens reduced from SD1 to SD10 at the training stage and from FD2 to SD10 at the testing stage of ANN model, while count of the same pathogens reduced for FD2–SD10 at the training stage and for FD1–SD10 at the testing stage of ANFIS model. Contrarily, the number of LAB in experimentally contaminated kefir with pathogens increased from day zero (0) of fermentation to day 10 of storage in refrigerator (Tables 2 and 3). The reduced number of pathogens indicated that the selected probiotic LAB in kefir milk antagonized the activity and growth of pathogens. Hence, the findings of the present study were in close agreement to the findings of Lim et al. (2020), who identified two strains of probiotic bacteria, Leuconostoc mesenteroides and Lactobacillus curvatus, isolated from kimchi, and were responsible to antagonize the growth of L. monocytogenes.
In the present study, the average count obtained in LAB control, tested LAB, and L. monocytogenes was 5.23, 4.94, and 2.41 log10 CFU/g, respectively, at the training stage, while at the testing stage, the respective values were 5.80, 5.38, and 2.04 log10 CFU/g (Table 2). However, in a study conducted by Gökmen et al. (2022), the obtained counts of strains of LAB, namely lactobacilli and lactic streptococci, were 9.64–7.91 log CFU/mL and 9.64–8.69 log CFU/mL, respectively. This result indicates that there is observable variation in the number of probiotic LAB, compared to the findings of the present study. However, results of the present study closely agreed with the findings of Jara et al. (2020), who identified the potential of probiotic Lactobacillus fermentum MP26 and Lactobacillus salivarius MP14, both inhibiting the activity and growth of Listeria monocytogenes. In addition, in the investigation conducted by Morandi et al. (2020), the inhibitory activity of LAB against L. monocytogenes in Gorgonzola cheese indicated counts of the pathogen as <2.0 log10 CFU/g, and this result was close to the result obtained during storage period at both training and testing stages of the present study.
In a previous study, the inhibitory effect of Lactobacillus rhamnosus in probiotic Minas Frescal cheese against L. monocytogenes was reported as 1.1–1.6 log CFU/g (Prezzi et al., 2020); this result was similar to the result obtained in the present study at SD1 (1.6 log10 CFU/g) and SD3 (1.6 log10 CFU/g) with ANFIS model, and at SD10 (1.18 log10 CFU/g) with ANN model. The inhibitory potential of probiotic LAB was further confirmed on biofilm-forming pathogenic L. monocytogenes present in vegetables and in the food industry without any risk to consumers (Hossain et al., 2020). Moreover, Martín et al. (2022) suggested that the strain of Lactiplantibacillus plantarum B2 alone or combined with Lactiplantibacillus spp. B4 are good candidates against growth of L. monocytogenes in traditional soft cheese obtained from dairy milk during their refrigerated storage. The survival of some foodborne pathogens, including both Gram-positive and Gram-negative bacteria, in kefir produced by microbial level and pullulan was determined by Gökmen et al. (2022), of which L. monocytogenes was the most susceptible bacterium to the metabolites of LAB during storage, with maximum reduction of pathogen after 24-h fermentation at 30°C.
The other targeted foodborne pathogen involved in the present study was S. aureus. The microbiological profile of this pathogen was evaluated and analyzed by both ANN and ANFIS models at specified time intervals as displayed in Tables 5 and 6. The analysis was also carried out for the evaluation of LAB control and test LAB for both fermentation and storage periods. Based on the analysis with ANN model, the average number of S. aureus was found as 2.04 log10 CFU/g, while the number of LAB control and test LAB was 5.23 log10 CFU/g and 4.89 log10 CFU/g, respectively, at the testing stage whereas with ANFIS model, the average number of S. aureus was recorded as 2.04 log10 CFU/g, while the number of LAB control and test LAB was 5.23 log10 CFU/g and 4.82 log10 CFU/g, respectively.
Recently published articles have indicated that the potential of LAB to inhibit the growth of S. aureus has been confirmed in various food products, such as cheese, yoghurt, kefir, and milk. Jiang et al. (2022) reported that strains of LAB isolated from traditional fermented yoghurt were found to have antibacterial and antibiofilm activity against S. aureus, indicating that the obtained result was similar to the finding of the present study. Likewise, the application of probiotic LAB to decrease the growth of S. aureus in the co-culture of sheep’s milk reported by Rodríguez-Sánchez et al. (2022) was nearly similar to the findings of the present study. In contrary to the present study, Prezzi et al. (2020) indicated that the efficacy of probiotic Lactobacillus rhamnosus isolated from Minas Frescal cheese during the storage period of 21 days at 7oC was not observed against pathogenic S. aureus
In the present study, the growth and activity of S. aureus was evaluated by testing in LAB of dairy kefir during fermentation days (FD0–FD2) and storage days (SD1–SD10) with both ANN and ANFIS models. As a result, count of the pathogen was found to decrease from FD2 to SD10 at the training stage and from FD1 to SD10 at the testing stage with ANN model (Table 5). On the other hand, the number of S. aureus reduced from FD2 to SD10 at both training and testing stages with ANFIS model (Table 6). Contrarily, the number of test LAB and LAB control increased from FD0 to SD10 during the refrigerated storage of experimental kefir. The inhibition of S. aureus could involve different metabolites of LAB, which are the most important compounds that inhibit the growth of undesirable microorganisms, particularly S. aureus, in food and pharmaceutical industries (Nataraj et al., 2021). Thus, the inhibition of S. aureus enhances food safety and hygiene to ensure the health of consumers.
Different studies have revealed that strains of probiotic LAB isolated from dairy food products possess antagonizing activity against S. aureus, particularly in food production environments (Folliero et al., 2022; Jiang et al., 2021; Nataraj et al., 2021; Tarique et al., 2022). For example, the efficacy of probiotic L. brevis gp104, which was isolated from Iranian traditional cheese, had a promising potential against the growth of S. aureus, and has potential health benefits for its application as a novel biotherapeutic and biopreserving agent (Hojjati et al., 2020). Similarly, the antagonizing activity of some probiotic LAB isolated from traditional high acid and low moisture yogurt-like products, including Streptococcus thermophilus, Lactobacillus delbrueckii, Enterococcus faecium, and Lacticaseibacillus rhamnosus, was reported against S. aureus in a study conducted by Tarique et al. (2022).
Moreover, in the present study, the inhibition of S. auerus was also confirmed by regression analysis at training, validation, and testing stages, with respective R = 0.9842, 0.9905, 0.8873, as indicated in Figure 6. In addition, the model reflected the evaluation of S. auerus with the best validation performance of 0.071812 at epoch 21 (Figure 7). Evaluation of the inhibitory activity of potential probiotic LAB of dairy kefir against S. aureus was also analyzed with ANFIS model as described in Table 6. The findings showed that reduction in the count of the targeted pathogen was observed from FD2 to SD10 at both training and testing stages whereas the number of LAB increased during the above-stated days. In this analysis, reduction in the count of S. auerus was high during storage days than during fermentation days of the experimental kefir. A study conducted by (Yan et al. (2019) confirmed that several strains of LAB exhibited antagonizing potential against foodborne pathogens; among of these strains were Pediococcus acidilactic and Lactococcus plantarum, both being promising probiotics against S. aureus CMCC 26003. Thus, the findings of the cited investigation were in close agreement to the results of the present study.
The inhibition of E. coli by probiotic LAB present in dairy kefir was analyzed using ANN model, with reduced number of pathogen from FD2 to SD10 whereas an increment in the number of LAB was observed from FD0 to SD10 (Table 7) at the training stage. Likewise, decrease in the number of E. coli was observed from FD0 to SD10 at the testing stage. However, the average result obtained for LAB control, tested LAB, and E. coli was 5.23, 4.96, and 2.46 log10 CFU/g, respectively, at the training stage whereas for the testing stage, the respective values were 5.80, 5.45, and 1.92 log10 CFU/g, as displayed in Table 7. Findings of the study conducted by Esfandiari et al. (2024) regarding the inhibitory activity of probiotic LAB against E. coli were in close agreement with the findings of the present study. Similarly, investigation done by de Amorim Trindade et al. (2022) and Darvishi et al. (2021) indicated that the strains of LAB exhibited probiotic potential against the growth of E. coli and other foodborne pathogens. Thus, it is obvious that the effects of various probiotic bacterial strains against the growth and activity of foodborne pathogenic bacteria, usually presenting in food products and the environment, are documented in different food processing plants. The strains of Lactobacillus, Bifidobacterium, Lactococcus, and Enterococcus are also responsible for inhibiting the growth of biofilm-forming E. coli and are applied as bio-control agents against spoilage and pathogenic bacteria in meat and meat products (Abouloifa et al., 2023; Darvishi et al., 2021).
In addition, inhibition of E. coli was also braced by regression analysis, with R = 0.9702, 0.9514 and 0.9537 at training, validation and testing stages, respectively. The best validation performance for the inhibition of E. coli was obtained at 0.18637. Similarly, the inhibition of pathogenic E. coli by biological means, namely probiotic LAB, which are naturally present in dairy kefir milk and used in the present study, was analyzed with ANFIS model. The model simulated with inhibition of E. coli at training stages with the average number of LAB in control (5.23 log10 CFU/g), tested LAB (4.93 log10 CFU/g), and E. coli (2.03 log10 CFU/g). Likewise, the average value of LAB control, tested LAB, and E. coli obtained at the testing stage were 5.65, 5.25, and 1.34 log10 CFU/g, respectively. In this model, inhibition of the activity of E. coli was observed along with the fermentation and storage days of kefir milk, which was confirmed by the reduced number of the pathogen from FD2 to SD10.
E. coli is among the well-known and most serious foodborne bacteria, causing severe health problems to consumers through adherence to the mucosal membrane of host’s intestines. To combat the activity of this pathogen, the application of probiotic food products, such as dairy products, is more reliable, as demonstrated in various studies. As evidenced, the strains of some probiotic LAB, reported in previous studies, including Lactobacillus sakei, Lactobacillus plantarum, Lactobacillus reuteri, Lactobacillus fermentum, and Lactobacillus casei, have an antagonizing activity toward pathogenic E. coli (Darvishi et al., 2021; de Amorim Trindade et al., 2022; Hansen et al., 2021). These strains of LAB possess desirable potential for passing through the low pH of the stomach and entering the intestines to inhibit the adherence activity of infectious E. coli (Behbahani et al., 2019). In the present study, the inhibition of E. coli was more observable during the storage period of dairy kefir stored at 4°C. This finding was in close agreement with the investigations of Choi et al. (2021), who confirmed the antibacterial influence of probiotic Leuconostoc mesenteroides (KCTC 13374) and Lactobacillus plantarum (KCTC 33133) isolated from commercially manufactured kimichi during fermentation at respective temperatures of 10°C and 25°C.
Conclusions and Recommendations
The present study was conducted on the inhibition of pathogenic foodborne pathogens, viz. L. monocytogenes, S. aureus, and E. coli, by probiotic LAB present naturally in dairy kefir using artificial intelligence models (ANN and ANFIS). The activity and growth of these foodborne pathogens was repressed by LAB present in dairy kefir. Thus, probiotic dairy kefir products are biological controlling means that can be applied to both food industry and agricultural sector. Based on the present study, the antibacterial activity of probiotic LAB in kefir was more observable during storage period than fermentation period. Therefore, based on the results, the following steps are recommended: first, more research work must be emphasized on the investigation of kefir as a potential probiotic antagonizing the activity of serious foodborne pathogens. Second, Artificial Intelligence-based approaches for the inhibition of targeted pathogens could be a baseline for more attention to the research.
Author Contributions
BC Kefyalew and BH Ulusoy designed the study. BC Kefyalew and F Kaya Yıldırım conducted the data analysis. BC Kefyalew drafted the manuscript and F Kaya Yıldırım revised the same. All authors read and approved the final manuscript.
Data Availability
The datasets used in this study are available from corresponding authors upon reasonable request.
Conflicts of Interest
The authors reported no conflict of interest.
Funding Statement
Not applicable.
REFERENCES
Abdelhamid, A.G., and El-Dougdoug, N.K., 2020. Controlling food borne pathogens with natural antimicrobials by biological control and anti-virulence strategies. Heliyon 6(9): e05020. 10.1016/j.heliyon.2020.e05020
Abouloifa, H., Gaamouche, S., Idrissi Yahyaoui, M., Moumnassi, S., Hasnaoui, I., Bellaouchi, R., and Asehraou, A., 2023. The efficiency of Lactiplantibacillus plantarum S61 strain as protective cultures in ground beef against foodborne pathogen Escherichia coli. World Journal of Microbiology and Biotechnology 39(12): 327. 10.1007/s11274-023-03763-5
Ağagündüz, D., Şahin, T.Ö., Ayten, Ş., Yılmaz, B., Güneşliol, B.E., Russo, P., and Özogul, F., 2022. Lactic acid bacteria as pro-technological, bioprotective and health-promoting cultures in the dairy food industry. Food Bioscience 47: 101617. 10.1016/j.fbio.2022.101617
Angelidis, A.S., Komodromos, D., Giannakou, R., Arsenos, G., Gelasakis, A.I., Kyritsi, M., and Sergelidis, D., 2020. Isolation and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus (MRSA) from milk of dairy goats under low-input farm management in Greece. Veterinary Microbiology 247: 108749. 10.1016/j.vetmic.2020.108749
Anokyewaa, M.A., Amoah, K., Li, Y., Lu, Y., Kuebutornye, F.K., Asiedu, B., and Seidu, I., 2021. Prevalence of virulence genes and antibiotic susceptibility of Bacillus used in commercial aquaculture probiotics in China. Aquaculture Reports 21: 100784. 10.1016/j.aqrep.2021.100784
Behbahani, B.A., Noshad, M., and Falah, F., 2019. Inhibition of Escherichia coli adhesion to human intestinal Caco-2 cells by probiotic candidate Lactobacillus plantarum strain L15. Microbial Pathogenesis 136: 103677. 10.1016/j.micpath.2019.103677
Chen, Y., Chen, M., Wang, J., Wu, Q., Cheng, J., Zhang, J., and Kou, X., 2020. Heterogeneity, characteristics, and public health implications of Listeria monocytogenes in ready-to-eat foods and pasteurized milk in China. Frontiers in Microbiology 11: 642. 10.3389/fmicb.2020.00642
Choi, H., Hwang, B.K., Kim, B.S., and Choi, S.H., 2020. Influence of pathogen contamination on beef microbiota under different storage temperatures. Food Research International 132: 109118. 10.1016/j.foodres.2020.109118
Choi, S.J., Yang, S.Y., and Yoon, K.S., 2021. Lactic acid bacteria starter in combination with sodium chloride controls pathogenic Escherichia coli (EPEC, ETEC, and EHEC) in kimchi. Food Microbiology 100: 103868. 10.1016/j.fm.2021.103868
Colombo, M., Castilho, N.P., Todorov, S.D., and Nero, L.A., 2018. Beneficial properties of lactic acid bacteria naturally present in dairy production. BMC Microbiology 18(1): Article No. 219. 10.1186/s12866-018-1356-8
Cufaoglu, G., Ambarcioglu, P., and Ayaz, N.D., 2021. Meta-analysis of the prevalence of Listeria spp., and antibiotic resistant L. monocytogenes isolates from foods in Turkey. Lebensmittel-Wissenschaft & Technologie (LWT) 144: 111210. 10.1016/j.lwt.2021.111210
Darvishi, N., Fard, N.A., and Sadrnia, M., 2021. Genomic and proteomic comparisons of bacteriocins in probiotic species Lactobacillus and Bifidobacterium and inhibitory ability of Escherichia coli MG 1655. Biotechnology Reports 31: e00654. 10.1016/j.btre.2021.e00654
de Amorim Trindade, D.P., Barbosa, J.P., Martins, E.M.F., and Tette, P.A.S., 2022. Isolation and identification of lactic acid bacteria in fruit processing residues Fromm te Brazilian Cerrado and its probiotic potential. Food Bioscience 48: 101739. 10.1016/j.fbio.2022.101739
Deng, F., Chen, Y., Sun, T., Wu, Y., Su, Y., Liu, C., and Wen, J., 2021. Antimicrobial resistance, virulence characteristics and genotypes of Bacillus spp. from probiotic products of diverse origins. Food Research International 139: 109949. 10.1016/j.foodres.2020.109949
Dimitreli, G., and Antoniou, K.D., 2011. Effect of incubation temperature and caseinates on the rheological behaviour of Kefir. Procedia Food Science 1: 583–588. 10.1016/j.profoo.2011.09.088
Duze, S.T., Marimani, M., and Patel, M., 2021. Tolerance of Listeria monocytogenes to biocides used in food processing environments. Food Microbiology 97: 103758. 10.1016/j.fm.2021.103758
El Hag, M.M., El Zubeir, I.E.M., and Mustafa, N.E., 2021. Prevalence of Listeria species in dairy farms in Khartoum State (Sudan). Food Control 123: 107699. 10.1016/j.foodcont.2020.107699
Esfandiari, Z., Vakili, B., Ahangarzadeh, S., Esfahani, S.N., and Shoaei, P., 2024. Impact of selenium nanoparticle-enriched Lactobacilli feeding against Escherichia coli O157: H7 infection of BALB/c mice. Probiotics and Antimicrobial Proteins 16: 784–795. 10.1007/s12602-023-10081-7
Fagerlund, A., Langsrud, S., and Møretrø, T., 2020. Microbial diversity and ecology of biofilms in food industry environments associated with Listeria monocytogenes persistence. Current Opinion in Food Science 38: 171–178. 10.1016/j.cofs.2020.10.015
Farha, A.K., Yang, Q.Q., Kim, G., Zhang, D., Mavumengwana, V., Habimana, O., and Gan, R.Y., 2020. Inhibition of multidrug-resistant foodborne Staphylococcus aureus biofilms by a natural terpenoid (+)-nootkatone and related molecular mechanism. Food Control 112: 107154. 10.1016/j.foodcont.2020.107154
Flora, M., Perrotta, F., Nicolai, A., Maffucci, R., Pratillo, A., Mollica, M., and Calabrese, C., 2019. Staphylococcus aureus in chronic airway diseases: an overview. Respiratory Medicine 155: 66–71. 10.1016/j.rmed.2019.07.008
Folliero, V., Lama, S., Franci, G., Giugliano, R., D’Auria, G., Ferranti, P., and Stiuso, P., 2022. Casein-derived peptides Fromm te dairy product kashk exhibit wound healing properties and antibacterial activity against Staphylococcus aureus: structural and functional characterization. Food Research International 153: 110949. 10.1016/j.foodres.2022.110949
Gökmen, G.G., Kowalik, J., and Kışla, D., 2022. Survival of some food-borne bacteria in kefir produced by microbial levan and pullulan. Food Bioscience 47: 101675. 10.1016/j.fbio.2022.101675
González-Orozco, B.D., García-Cano, I., Jiménez-Flores, R., and Alvárez, V.B., 2022. Invited review: milk kefir microbiota-direct and indirect antimicrobial effects. Journal of Dairy Science 105: 2021–21382. 10.3168/jds.2021-21382
Gu, T., Luo, Y., Jia, Z., Meesrison, A., Lin, S., Ventresca, I.J., and Zhang, B., 2024. Surface topography and chemistry of food contact substances, and microbial nutrition affect pathogen persistence and symbiosis in cocktail Listeria monocytogenes biofilms. Food Control 161: 110391. 10.1016/j.foodcont.2024.110391
Gut, A.M., Vasiljevic, T., Yeager, T., and Donkor, O.N., 2022. Anti-salmonella properties of kefir yeast isolates: an in vitro screening for potential infection control. Saudi Journal of Biological Sciences 29(1): 550–563.
Hansen, L.H.B., Nielsen, B., Boll, E.J., Skjøt-Rasmussen, L., Wellejus, A., Jørgensen, L., and Canibe, N., 2021. Functional in vitro screening of probiotic strains for inoculation of piglets as a prophylactic measure towards enterotoxigenic Escherichia coli infection. Journal of Microbiological Methods 180: 106126. 10.1016/j.mimet.2020.106126
Hojjati, M., Behabahani, B.A., and Falah, F., 2020. Aggregation, adherence, anti-adhesion and antagonistic activity properties relating to surface charge of probiotic Lactobacillus brevis gp104 against Staphylococcus aureus. Microbial Pathogenesis 147: 104420. 10.1016/j.micpath.2020.104420
Hossain, M.I., Mizan, M.F.R., Ashrafudoulla, M., Nahar, S., Joo, H.J., Jahid, I.K., and Ha, S.D., 2020. Inhibitory effects of probiotic potential lactic acid bacteria isolated from kimchi against Listeria monocytogenes biofilm on lettuce, stainless steel surfaces, and MBEC™ biofilm device. Lebensmittel-Wissenschaft & Technologie (LWT) 118: 108864. 10.1016/j.lwt.2019.108864
Jara, J., Pérez-Ramos, A., Del Solar, G., Rodríguez, J.M., Fernández, L., and Orgaz, B., 2020. Role of lactobacillus biofilms in Listeria monocytogenes adhesion to glass surfaces. International Journal of Food Microbiology 334: 108804. 10.1016/j.ijfoodmicro.2020.108804
Jiang, X., Ren, S., Geng, Y., Jiang, C., Liu, G., Wang, H., and Liang, Y., 2021. Role of te VirSR-VirAB system in biofilm formation of Listeria monocytogenes EGD-e. Food Research International 145: 110394. 10.1016/j.foodres.2021.110394
Jiang, Y.H., Xin, W.G., Yang, L.Y., Ying, J.P., Zhao, Z.S., Lin, L.B., et al. 2022. A novel bacteriocin against Staphylococcus aureus Fromm Lactobacillus paracasei isolated Fromm Yunnan traditional fermented yogurt: purification, antibacterial characterization, and antibiofilm activity. Journal of Dairy Science 105(3): 2094–2107. 10.3168/jds.2021-21126
John, S.M., and S. Deeseenthum., 2015. Properties and benefits of kefir—a review. Songklanakarin Journal of Science and Technology (SJST) 37: 275–282.
Kallipolitis, B., Gahan, C.G., and Piveteau, P., 2020. Factors contributing to Listeria monocytogenes transmission and impact on food safety. Current Opinion in Food Science 36: 9–17. 10.1016/j.cofs.2020.09.009
Kamal, R.M., Alnakip, M.E., Abd El Aal, S.F., and Bayoumi, M.A., 2018. Bio-controlling capability of probiotic strain Lactobacillus rhamnosus against some common foodborne pathogens in yoghurt. International Dairy Journal 85: 1–7. 10.1016/j.idairyj.2018.04.007
Kannan, S., Balakrishnan, J., and Govindasamy, A., 2020. Listeria monocytogens-amended understanding of its pathogenesis with a complete picture of its membrane vesicles, quorum sensing, biofilm and invasion. Microbial Pathogenesis 149: 104575. 10.1016/j.micpath.2020.104575
Kaya, H.I., and Simsek, O., 2019. Characterization of pathogen-specific bacteriocins from lactic acid bacteria and their application within cocktail against pathogens in milk. Lebensmittel-Wissenschaft & Technologie (LWT) 115: 108464. 10.1016/j.lwt.2019.108464
Keba, A., Rolon, M.L., Tamene, A., Dessie, K., Vipham, J., Kovac, J., and Zewdu, A., 2020. Review of the prevalence of foodborne pathogens in milk and dairy products in Ethiopia. International Dairy Journal 109: 104762. 10.1016/j.idairyj.2020.104762
Kefyalew, B.C., Ulusoy, B.H., Metekia, W.A., and Kaya Yıldırım, F., 2021. In vitro probiotic and industrial properties of bacteria isolated from fermented food products. International Food Research Journal, 28(4): 638–653. 10.47836/ifrj.28.4.01
Khaneghah, A.M., Abhari, K., Eş, I., Soares, M.B., Oliveira, R.B., Hosseini, H., and Sant’Ana, A.S., 2020. Interactions between probiotics and pathogenic microorganisms in hosts and foods: a review. Trends in Food Science and Technology 95: 205–218. 10.1016/j.tifs.2019.11.022
Kim, J.A., Bayo, J., Cha, J., Choi, Y.J., Jung, M.Y., Kim, D.H., and Kim, Y., 2019. Investigating the probiotic characteristics of four microbial strains with potential application in the feed industry. PLoS ONE, 14(6): e0218922. 10.1371/journal.pone.0218922
Klimko, A.I., Cherdyntseva, T.A., Brioukhanov, A.L., and Netrusov, A.I., 2020. In vitro evaluation of probiotic potential of selected lactic acid bacteria strains. Probiotics and Antimicrobial Proteins 12(3): 1139–1148. 10.1007/s12602-019-09599-6
Kouhi, F., Mirzaei, H., Nami, Y., Khandaghi, J., and Javadi, A., 2022. Potential probiotic and safety characterisation of Enterococcus bacteria isolated from indigenous fermented Motal cheese. International Dairy Journal 126: 105247. 10.1016/j.idairyj.2021.105247
Lim, J.Y., Lee, C.L., Kim, G.H., Bang, Y.J., Rhim, J.W., and Yoon, K.S., 2020. Using lactic acid bacteria and packaging with grapefruit seed extract for controlling Listeria monocytogenes growth in fresh soft cheese. Journal of Dairy Science 103(10): 8761–8770. 10.3168/jds.2020-18349
Ly, V., Parreira, V.R., and Farber, J.M., 2019. Current understanding and perspectives on Listeria monocytogenes in low-moisture foods. Current Opinion in Food Science 26: 18–24. 10.1016/j.cofs.2019.02.012
Martín, I., Rodríguez, A., Alía, A., Martínez-Blanco, M., Lozano-Ojalvo, D., and Córdoba, J.J., 2022. Control of Listeria monocytogenes growth and virulence in a traditional soft cheese model system based on lactic acid bacteria and a whey protein hydrolysate with antimicrobial activity. International Journal of Food Microbiology 361: 109444. 10.1016/j.ijfoodmicro.2021.109444
Mkadem, W., Belguith, K., Oussaief, O., ElHatmi, H., Indio, V., Savini, F., and Boudhrioua, N., 2023. Systematic approach to select lactic acid bacteria from spontaneously fermented milk able to fight Listeria monocytogenes and Staphylococcus aureus. Food Bioscience 51: 102275. 10.1016/j.fbio.2022.102275
Morandi, S., Silvetti, T., Vezzini, V., Morozzo, E., and Brasca, M., 2020. How we can improve the antimicrobial performances of lactic acid bacteria? A new strategy to control Listeria monocytogenes in Gorgonzola cheese. Food Microbiology 90: 103488. 10.1016/j.fm.2020.103488
Mulaw, G., Tessema, T.S., Muleta, D., and Tesfaye, A., 2019. In vitro evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented Ethiopian food products. International Journal of Microbiology 2019: 7179514. 10.1155/2019/7179514
Muñoz, N., Sonar, C.R., Bhunia, K., Tang, J., Barbosa-Cánovas, G.V., and Sablani, S.S., 2019. Use of protective culture to control the growth of Listeria monocytogenes and Salmonella typhimurium in ready-to-eat cook-chill products. Food Control 102: 81–86. 10.1016/j.foodcont.2019.03.009
Nataraj, B.H., Ramesh, C., and Mallappa, R.H., 2021. Characterization of biosurfactants derived from probiotic lactic acid bacteria against methicillin-resistant and sensitive Staphylococcus aureus isolates. Lebensmittel-Wissenschaft & Technologie (LWT) 151: 112195. 10.1016/j.lwt.2021.112195
Niaz, T., Shabbir, S., Noor, T., and Imran, M., 2019. Antimicrobial and antibiofilm potential of bacteriocin loaded nano-vesicles functionalized with rhamnolipids against foodborne pathogens. Lebensmittel-Wissenschaft & Technologie (LWT) 116: 108583. 10.1016/j.lwt.2019.108583
Olaimat, A.N., Ghoush, M.A., Al-Holy, M., Hilal, H.A., Al-Nabulsi, A.A., Osaili, T.M., and Holley, R.A., 2021. Survival and growth of Listeria monocytogenes and Staphylococcus aureus in ready-to-eat Mediterranean vegetable salads: impact of storage temperature and food matrix. International Journal of Food Microbiology 346: 109149. 10.1016/j.ijfoodmicro.2021.109149
Özkan, E.R., Demirci, T., and Akın, N., 2021. In vitro assessment of probiotic and virulence potential of Enterococcus faecium strains derived from artisanal goatskin casing Tulum cheeses produced in central Taurus Mountains of Turkey. Lebensmittel-Wissenschaft & Technologie (LWT) 141: 110908. 10.1016/j.lwt.2021.110908
Paongphan, P., Ditudompo, S., Vitheejongjaroen, P., Pachekrepapol, U., and Taweechotipatr, M. 2023. Selected lactobacilli isolated from Thai foods for production of fermented dairy products with cholesterol lowering potential. NFS Journal 33: 100151. 10.1016/j.nfs.2023.100151
Prezzi, L.E., Lee, S.H., Nunes, V.M., Corassin, C.H., Pimentel, T.C., Rocha, R.S., and Oliveira, C.A., 2020. Effect of Lactobacillus rhamnosus on growth of Listeria monocytogenes and Staphylococcus aureus in a probiotic Minas Frescal cheese. Food Microbiology 92: 103557. 10.1016/j.fm.2020.103557
Rajabi, S., Darban, D., Tabatabaei, R.R., and Hosseini, F., 2020. Antimicrobial effect of spore-forming probiotics Bacillus laterosporus and Bacillus megaterium against Listeria monocytogenes. Archives of Microbiology 202(10): 2791–2797. 10.1007/s00203-020-02004-9
Ranjbar, R., and Halaji, M., 2018. Epidemiology of Listeria monocytogenes prevalence in foods, animals and human origin from Iran: a systematic review and meta-analysis. BMC Public Health 18(1): 1–12. 10.1186/s12889-018-5966-8
Rivas-Macho, A., Eletxigerra, U., Diez-Ahedo, R., Merino, S., Goñi-de-Cerio, F., and Olabarria, G., 2024. LAMP-based electrochemical sensor for extraction-free detection of Listeria monocytogenes in food samples. Food Control 163(11): 110546. 10.1016/j.foodcont.2024.110546
Rodríguez-Sánchez, S., Ramos, I.M., Rodríguez-Pérez, M., Poveda, J.M., Seseña, S., and Palop, M.L., 2022. Lactic acid bacteria as biocontrol agents to reduce Staphylococcus aureus growth, enterotoxin production and virulence gene expression. Lebensmittel-Wissenschaft & Technologie (LWT) 170: 114025. 10.1016/j.lwt.2022.114025
Roldán-Pérez, S., Rodríguez, S.L.G., Sepúlveda-Valencia, J.U., Villadiego, O.S.R., Fernández, M.E.M., Campuzano, O.I.M., and Durango-Zuleta, M.M., 2023. Assessment of probiotic properties of lactic acid bacteria isolated from an artisanal Colombian cheese. Heliyon 9(11): e21558. 10.1016/j.heliyon.2023.e21558
Rosario, A.I., Castro, V.S., Santos, L.F., Lisboa, R.C., Vallim, D.C., Silva, M.C., and Costa, M.P., 2021. Shiga toxin-producing Escherichia coli isolated from pasteurized dairy products from Bahia, Brazil. Journal of Dairy Science 104(6): 6535–6547. 10.3168/jds.2020-19511
Rosengren, Å., Fabricius, A., Guss, B., Sylvén, S., and Lindqvist, R., 2010. Occurrence of foodborne pathogens and characterization of Staphylococcus aureus in cheese produced on farm dairies. International Journal of Food Microbiology 144(2): 263–269. 10.1016/j.ijfoodmicro.2010.10.004
Rosa, D.D., Dias, M.M.S., Grześkowiak, Ł.M., Reis, S.A., Conceição, L.L., and Peluzio, M.D.C.G., 2017. Milk kefir: nutritional, microbiological and health benefits. Nutrition Research Reviews 30: 82–96. 10.1017/S0954422416000275
Rubab, M., Shahbaz, H.M., Olaimat, A.N., and Oh, D.H., 2018. Biosensors for rapid and sensitive detection of Staphylococcus aureus in food. Biosensors and Bioelectronics 105: 49–57. 10.1016/j.bios.2018.01.023
Selover, B., Johnson, J., and Waite-Cusic, J.G., 2021. Population dynamics of coliforms in a commercial Cheddar cheese production facility. Journal of Dairy Science 104(7): 7480–7488. 10.3168/jds.2020-19808
Shahverdi, S., Barzegari, A.A., Bakhshayesh, R.V., and Nami, Y., 2023. In vitro and in vivo antibacterial activity of potential probiotic Lactobacillus paracasei against Staphylococcus aureus and Escherichia coli. Heliyon 9(4): e14641. 10.1016/j.heliyon.2023.e14641
Soares, M.B., Almada, C.N., Pereira, E.P., Ferreira, B.M., Balthazar, C.F., Khorshidian, N., and Sant’Ana, A.S., 2023. Sporeforming probiotic bacteria: characteristics, health benefits, and technological aspects for their applications in foods and beverages. Trends in Food Science & Technology 138: 453–469. 10.1016/j.tifs.2023.06.029
Steinbrecher, M., Wolfert, C., Maurer, C., Messmann, H., Shiban, E., Sommer, B., and Fuchs, A., 2023. Cerebral abscess due to Listeria monocytogenes infection in silent diabetes mellitus: case presentation, treatment and patient outcome. ID Cases 33: e01864. 10.1016/j.idcr.2023.e01864
Tarique, M., Abdalla, A., Masad, R., Al-Sbiei, A., Kizhakkayil, J., Osaili, T., ..., and Ayyash, M., 2022. Potential probiotics and postbiotic characteristics including immunomodulatory effects of lactic acid bacteria isolated from traditional yogurt-like products. Lebensmittel-Wissenschaft & Technologie (LWT) 159: 113207. 10.1016/j.lwt.2022.113207
Taylor, M.H., and Zhu, M.J., 2021. Control of Listeria monocytogenes in low-moisture foods. Trends in Food Science & Technology, 116: 802–814. 10.1016/j.tifs.2021.07.019
Titouche, Y., Hakem, A., Houali, K., Meheut, T., Vingadassalon, N., Ruiz-Ripa, L., and Auvray, F., 2019. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) ST8 in raw milk and traditional dairy products in te Tizi Ouzou area of Algeria. Journal of Dairy Science 102(8): 6876–6884. 10.3168/jds.2018-16208
Ulusoy, B.H., Çolak, H., Hampikyan, H., and Erkan, M.E., 2007. An in vitro study on the antibacterial effect of kefir against some foodborne pathogens. Turkish Society of Microbiology 37:103–107.
Vahdat, F., Mehdizadeh, T., Kazemeini, H., Reale, A., and Kaboudari, A., 2024. Physicochemical, microbial, and sensory characteristics of yogurt with Persian shallot (Allium B Boiss) and probiotic bacteria. Food Science & Nutrition Corpus ID: 267693677. 10.1002/fsn3.4036
Wang, Y., Li, A., Zhang, L., Waqas, M., Mehmood, K., Iqbal, M., and Li, J., 2019. Probiotic potential of Lactobacillus on the intestinal microflora against Escherichia coli induced mice model through high-throughput sequencing. Microbial Pathogenesis 137: 103760. 10.1016/j.micpath.2019.103760
Wang, H., Ma, K., Shen, J., Fang, M., Pei, H., Li, Y., and Xue, T., 2023. Genes associated with desiccation stress in foodborne Staphylococcus aureus as revealed by transposon insertion mutagenesis. Food Research International 163: 112271. 10.1016/j.foodres.2022.112271
Wu, M., Dong, Q., Ma, Y., Yang, S., Aslam, M.Z., Liu, Y., and Li, Z., 2022. Potential antimicrobial activities of probiotics and their derivatives against Listeria monocytogenes in food field: a review. Food Research International 160: 111733. 10.1016/j.foodres.2022.111733
Yan, X., Gu, S., Cui, X., Shi, Y., Wen, S., Chen, H., and Ge, J., 2019. Antimicrobial, anti-adhesive and anti-biofilm potential of biosurfactants isolated Fromm Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus aureus CMCC26003. Microbial Pathogenesis 127: 12–20. 10.1016/j.micpath.2018.11.039
Yang, Q., Chen, J., Dai, J., He, Y., Wei, K., Gong, M., and Yang, B., 2024a. Total coliforms, microbial diversity and multiple characteristics of Salmonella in soil-irrigation water-fresh vegetable system in Shaanxi, China. Science of te Total Environment 924: 171657. 10.1016/j.scitotenv.2024.171657
Yang, X., Peng, Z., He, M., Li, Z., Fu, G., Li, S., and Zhang, J., 2024b. Screening, probiotic properties, and inhibition mechanism of a Lactobacillus antagonistic to Listeria monocytogenes. Science of The Total Environment 906: 167587. 10.1016/j.scitotenv.2023.167587
Yilmaz, B., Sharma, H., Melekoglu, E., and Ozogul, F., 2022. Recent developments in dairy kefir-derived lactic acid bacteria and their health benefits. Food Bioscience 46: 101592. 10.1016/j.fbio.2022.101592
Zhao, Y., Xia, D., Ma, P., Gao, X., Kang, W., and Wei, J., 2020. Advances in the detection of virulence genes of Staphylococcus aureus originate from food. Food Science and Human Wellness 9(1): 40–44. 10.1016/j.fshw.2019.12.004
Zhao, X., Yuan, X., Hu, M., Zhang, Y., Li, L., Zhang, Q., and Liu, Y., 2021. Prevalence and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus isolated Fromm bulk tank milk in Shandong dairy farms. Food Control 125: 107836. 10.1016/j.foodcont.2020.107836