
Download
RESEARCH ARTICLE
Resveratrol distribution in peanuts and its resistance to aflatoxin accumulation
Boyi Yang1,2, Wei Liu3, Li Yu1, Xiaofeng Hu1,4, Kai Hu4, Zhaowei Zhang1,3,4*, Peiwu Li1,3,4*
1Key Laboratory of Biology and Genetic Improvement of Oil Crops, Laboratory of Risk Assessment for Oilseed Products (Wuhan), Quality Inspection and Test Center for Oilseed Products, National Reference Lab for Agricultural Testing (Biotoxin), Ministry of Agriculture and Rural Affairs; Oil Crops Research Institute, Chinese Academy of Agricultural Sciences, Wuhan, P.R. China;
2Department of Forensic Medicine of Tongji Medical College, Huazhong University of Science and Technology, Wuhan, P.R. China;
3Institute of Quality Standard and Testing Technology Research, Sichuan Academy of Agricultural Sciences, Chengdu, P.R. China;
4Hubei Hongshan Lab, Wuhan, P.R. China
Abstract
The presence of aflatoxin is a major risk factor in peanut processing and consumption. It has been found that resveratrol, which is known to have numerous health benefits, plays a crucial role in resisting aflatoxin. However, the distribution of resveratrol in peanuts is not clearly understood. To address this issue, a peanut recombinant inbred line (RIL) population consisting of 240 lines was used to investigate the distribution of resveratrol and examine its correlation with aflatoxin resistance. We found that resveratrol is controlled by multiple genes, with a significantly higher maximum content (867.0 µg/kg) in the recombinant inbred population compared to the parental population. Resveratrol content in peanuts showed a significant negative correlation with both aflatoxin B1 levels and total aflatoxin content when it exceeded 300 µg/kg (correlation coefficients: −0.61 and −0.62, respectively). Exceeding 300 µg/kg of resveratrol in peanuts effectively inhibited aflatoxin production, suggesting that peanut varieties enriched with resveratrol have resistance against aflatoxin pollution.
Key words: aflatoxin accumulation, peanut, resveratrol distribution
*Corresponding Author: Zhaowei Zhang and Peiwu Li, Chinese Academy of Agricultural Sciences, Wuhan, P.R. China. Emails: [email protected]; [email protected]
Academic Editor: Matteo Bordiga, PhD, Dipartimento di Scienze del Farmaco, Food Chemistry, Biotechnology and Nutrition Unit, Università del Piemonte Orientale “A. Avogadro”, Largo Donegani 2, 28100 Novara, Italy
Received: 2 March 2024; Accepted: 20 September 2024; Published: 1 July 2025
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Resveratrol (3,5,4’-trihydroxy-trans-stilbene) is a naturally occurring stilbene phytoalexin phenolic compound in grape skins, peanuts, and red wine (Frémont, 2000). Resveratrol is beneficial for human health due to its antioxidant, cardioprotective, chemopreventative, anti-inflammatory, and estrogenic properties, as well as its interaction with signal transduction pathways (Bian et al., 2022; King et al., 2006; Luo et al., 2022; Mingrou et al., 2022; Ros 2010; Toomer 2018; Wu et al., 2022). It is also involved in resistance or tolerance to various biotical and abiotic stresses (Chung et al., 2003; Galiniak et al., 2019; Guo et al., 2022; Jeandet et al., 2002; Mwakinyali et al., 2019; Qin-fei et al., 2015). Peanut (Arachis hypogaea L.) is a key source of resveratrol (Chung et al., 2003; Sales & Resurreccion, 2014), with significantly different content levels reported (Chen Hai-wen, 2021; Lee et al., 2004; Ogaki & Sagawa, 2003; Wang & Pittman, 2009). Nevertheless, peanuts are vulnerable to aflatoxin contamination. Aflatoxins are highly toxic, carcinogenic, immunogenic, and teratogenic (Marshall et al., 2020; Meneely et al., 2023).
Resveratrol is a potential candidate for resisting biological stresses such as Aspergillus flavus, thereby reducing the production of aflatoxins. A resveratrol concentration exceeding 3.0 μg/mL significantly reduced aflatoxin production and accumulation (Wang et al., 2013), suggesting the capability of resveratrol to inhibit aflatoxin production.
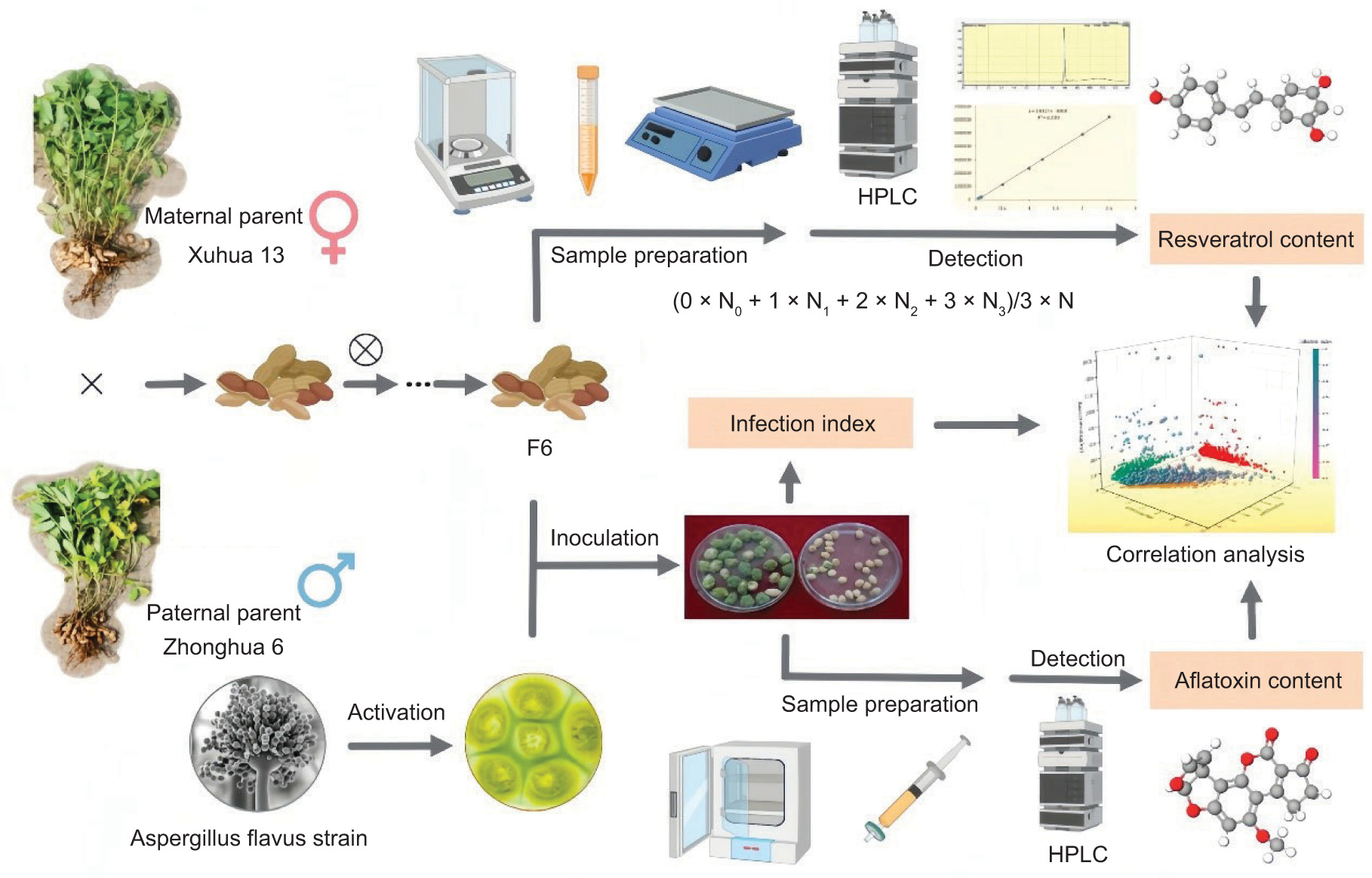
In this study, we investigated the genetic distribution of peanut resveratrol and its impact on resistance against aflatoxin contamination. The peanut variety Xuhua 13 was selected as the maternal parent (P1), while Zhonghua 6 was chosen as the paternal parent (P2) (Yu et al., 2023). A total of 240 lines from its F6-generation recombinant inbred line population were used as experimental materials to analyze the resveratrol content in P1, P2, and F6. Additionally, the distribution of peanut resveratrol content in the recombinant inbred line population was investigated. We examined aflatoxin production and infection index in peanuts inoculated with Aspergillus flavus at different levels of resveratrol content. This work could provide useful information on resistance to aflatoxin contamination (Figure 1).
Figure 1. Scheme of resveratrol resistance to aflatoxin accumulation.
Materials and Methods
Reagents and instruments
Resveratrol, aflatoxin B1 (AFB1), aflatoxin B2 (AFB2), aflatoxin G1 (AFG1), and aflatoxin G2 (AFG2), solid phase extraction column, and immunoaffinity column were procured from Sigma-Aldrich (USA). Acetonitrile, methanol, Tween-80, potato, agar, glucose, ethanol, alumina, and sodium chloride were sourced from Wuhan Aolaite Biotechnology Co., Ltd. (China). Ultrapure water was obtained from a Millipore Milli-Q system. All the chemicals were of analytical grade unless otherwise specified.
A high-performance liquid chromatography (HPLC) system with a fluorescence detector (Agilent 1200, USA) was employed to detect the content of AFB1, AFB2, AFG1, and AFG2, while resveratrol content was detected using an HPLC with a UV detector (Shimadzu LC-20A, Japan). The biosafety cabinet was purchased from Esco Micro Pte. Ltd. (Singapore).
Experimental material
The experimental materials consisted of a recombinant inbred line population of 240 lines derived from the maternal parent Xuhua 13 and the paternal parent Zhonghua 6. Both peanut varieties have distinct characteristics, Xuhua 13 was developed by Xuzhou Academy of Agricultural Sciences, China, while Zhonghua 6 was home-produced for its resistance to Aspergillus flavus. The F6 generation and the two parents were planted in Wuhan, China. Field management followed standard agricultural practices (Li et al., 2022).
A highly infective and toxigenic strain AF2202 (Wang et al., 2016) of Aspergillus flavus was isolated and purified from peanut pods, and stored at −80°C before use. The experimental materials were divided into two parts. One part was used to determine resveratrol content and study its genetic regulation in peanuts, while the other part was inoculated with the Aspergillus flavus strain to investigate the correlation between resveratrol content and aflatoxin production.
Resveratrol determination
Resveratrol was determined according to a previous study, with some modifications (Chen Hai-wen, 2021). A 5-gram crushed peanut sample was mixed with 40 mL of 85% ethanol solution and heated at 80°C for 45 min. An aliquot of 8 mL was cleaned using a solid phase extraction column. The filtrate was concentrated to 1 mL and filtered through a membrane with a pore size of 0.22 μm. To prevent the conversion of resveratrol isomers under light conditions, the entire experiment was conducted in darkness (Chen Hai-wen, 2021). Resveratrol content was determined by HPLC analysis with triplicate repeats. The HPLC conditions were as follows: C18 column (4.6 mm × 250 mm, particle size: 5 μm), gradient elution, flow rate: 0.8 mL/min, column temperature: 40°C, detection wavelength: 306 nm, and injection volume: 10 μL.
Activation of Aspergillus flavus strain and preparation of inoculated bacteria solution
The Aspergillus flavus strain was activated, and an inoculated bacterial solution was prepared using AF2202 conidia mixed with 0.05% Tween, resulting in a concentration of 4.0×106 CFU/mL. A 10 μL aliquot of AF2202 conidial liquid (4.0×106 CFU/mL) on a PDA plate was incubated at 29±1°C, 80%~90% humidity, in the dark for 7 days to obtain fresh AF2202 conidial liquid with the same level of activity and vigor. Fresh conidia were washed with 0.05% Tween in a biosafety cabinet to obtain a sufficient conidia solution. The conidial solution was then diluted with 0.05% Tween solution to prepare a conidial suspension at 4.0×106 CFU/mL (validated via the blood cell counting plate) for inoculating peanuts with Aspergillus flavus.
Inoculation and culture of Aspergillus flavus in peanut
Twenty mature, plump, healthy peanuts with normal color and uniform size were selected. The surface of peanut seeds was disinfected using a 70% ethanol solution for 1.0 min in a biosafety cabinet, followed by washing with sterile dd H2O 3 times. After cleaning, the peanut seeds were mixed with 0.5 mL Aspergillus flavus conidial suspension for the inoculation. Inoculated peanut seeds were incubated at 29±1°C and relative humidity of 80%~90% for 7 days in the dark before being sterilized at 121°C for 30 min. Finally, after cooling down, AFB1, AFB2, AFG1, and AFG2 content were determined through three repetitions per sample.
Peanut infection index
The infection index was categorized into four levels: level 0 (spore coverage rate of 0), level 1 (spore coverage rate of 0%–25%), level 2 (spore coverage rate of 25%–50%), and level 3 (spore coverage rate >50%). The peanut infection index = (0×N0 + 1×N1 + 2×N2 + 3×N3) / (3N), where N0, N1, N2, and N3 represented the number of peanut seeds at levels 0, 1, 2, and 3 in the population, respectively. “N” represents the sample size (Jing Dan, 2021).
Aflatoxin detection
The aflatoxin concentration was determined after inoculation with Aspergillus flavus. An amount of 5 g crushed peanuts was shaken with 1 g NaCl and 50 mL 70% methanol for 20 min, before being centrifuged at 6000 r/min for 10 min. A 1 mL initial extract was diluted with 49.0 mL 50% methanol solution. After cleanup using an immunoaffinity column, the dilution was injected into an HPLC with fluorescence detection on a C18 column (250 mm × 46 mm, particle size: 5.0 μm). Chromatographic conditions included an excitation wavelength of 360 nm and an emission wavelength of 440 nm with photochemical post-derivative treatment; the mobile phase consisted of methanol (55 mL) and water (45 mL), iso-degree elution, and a flow rate set at 0.8 mL/min. The column temperature was maintained at 40°C, while the sample size used was 10.0 µL.
Statistical analysis
We analyzed the resveratrol content distribution in F6-generation peanuts and its correlation with resistance to aflatoxin accumulation and Aspergillus flavus infection.
Results and Discussion
Resveratrol content
The resveratrol content in Xuhua 13 variety and the paternal Zhonghua 6 was 44 and 116 µg/kg, respectively. Among the 240 recombinant inbred populations, resveratrol content ranged from 37.3 to 867.0 µg/kg, with values at the 5 %, 50 %, and 95 % levels, as well as an average value of 52.9, 96.7, 270, and 127.31µg/kg (Table 1). Both Wang et al (2013) and Sanders et al (2000) reported that resveratrol in peanuts is trans-resveratrol, which is consistent with the results of this experimental study.
Table 1. Distribution of resveratrol and AFB1, AFB2, AFT and infection index in peanut.
| Resveratrol, µg/kg | AFB1, mg/kg | AFB2, mg/kg | AFT, mg/kg | Infection index | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P5 | P50 | P95 | AVE | P5 | P50 | P95 | AVE | P5 | P50 | P95 | AVE | P5 | P50 | P95 | AVE | P5 | P50 | P95 | AVE | |
| Zhonghua 6 | 116 | 30.68 | 2.00 | 32.68 | 0.89 | |||||||||||||||
| Xuhua 13 | 44 | 52.03 | 4.74 | 56.78 | 0.91 | |||||||||||||||
| F6 | 52.9 | 96.7 | 270.0 | 127.31 | 60.06 | 107.42 | 198.19 | 112.93 | 3.58 | 8.06 | 15.65 | 8.57 | 63.48 | 115.37 | 215.09 | 121.50 | 0.68 | 0.84 | 0.96 | 0.83 |
P5, P50, P95, and AVE represent the 5% value, 50% value, 95% value, and average value, respectively.
In the F6 recombinant inbreeding population, 3 individuals had lower resveratrol contents than the maternal Xuhua 13 variety (44 µg/kg). A total of 155 individuals showed intermediate contents between those of their parent varieties, and 82 individuals displayed higher resveratrol contents than that of the paternal Zhonghua 6 variety (116 µg/kg). The highest resveratrol level reached 867.0 µg/kg, which was 7.5 times greater than that in its father and 20 times more abundant than in its mother.
Aflatoxin concentration
There is a correlation between aflatoxins and peanut varieties, as different varieties can influence aflatoxin production and contamination levels. Varieties with good disease resistance tend to have a lower risk of aflatoxin contamination, whereas those with poor disease resistance are more susceptible. Our study focused on examining the relationship between aflatoxin levels and peanut species. AF2202 infection experiments resulted in predominant AFB1, accounting for 89.13% to 96.89% of total aflatoxins. AFB1, AFB2, and total aflatoxins (AFT) in the Xuhua 13 variety were 52.03, 4.74, and 56.78 mg/kg, while those in the Zhonghua 6 variety were 30.68, 2, and 32.68 mg/kg, respectively. Among the 240 recombinant inbred populations, AFB1, AFB2, and AFT concentrations were 17.04–325.40, 0.70–31.95, and 17.74–357.35 mg/kg, respectively. The 5%, 50%, and 95% quantile values of AFB1 were 60.06, 107.42, and 198.19 mg/kg, with an average value of 112.93 mg/kg. The 5%, 50%, and 95% quantile values of AFB2 were 3.58, 8.06, and 15.65 mg/kg, with an average value of 8.57 mg/kg. AFT’s 5%, 50%, and 95% quantile values were 63.48, 115.37, and 215.09 mg/kg, averaging 121.50 mg/kg (Table 1).
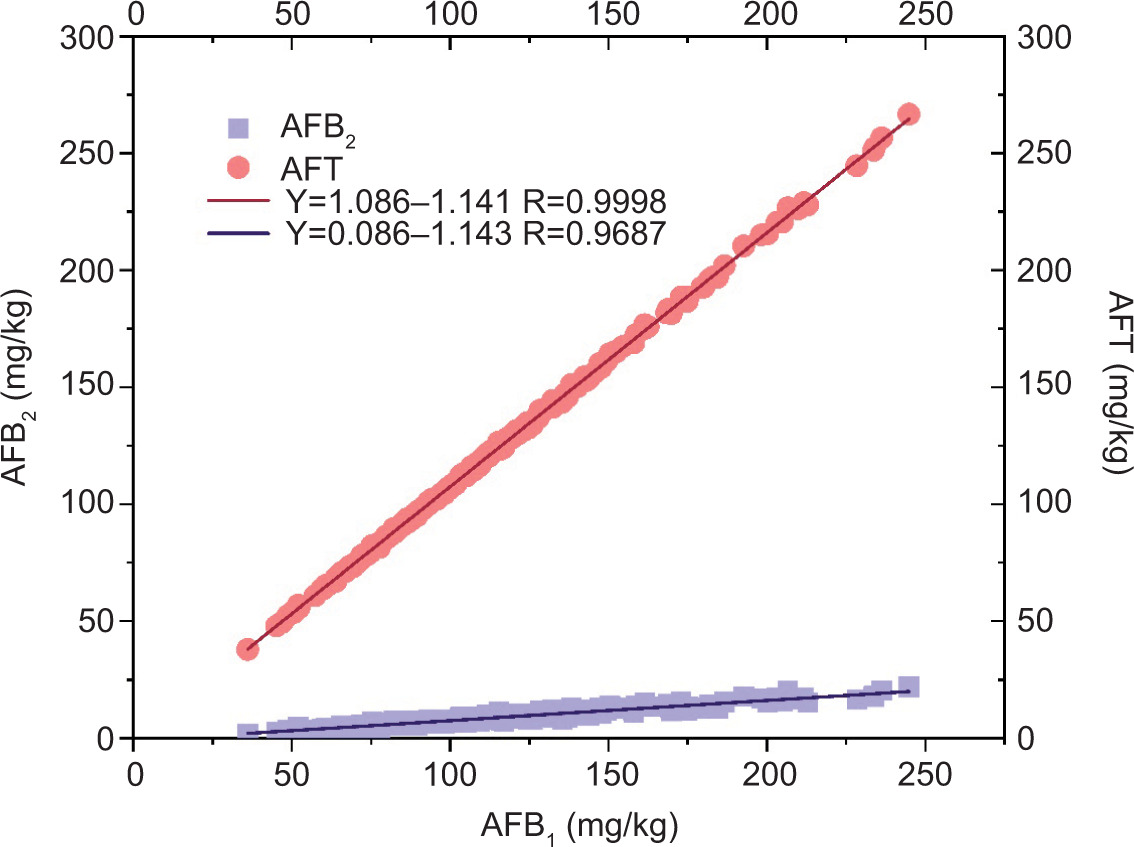
AFB1 exhibited a positive correlation with AFT (correlation coefficient = 0.9998). A significant positive correlation was also found between AFB1 and AFB2 (correlation coefficient = 0.9687) (Figure 2).
Figure 2. Correlation analysis of AFT with AFB1 and AFB2 of Xuhua 13 and Zhonghua 6.
Infection index
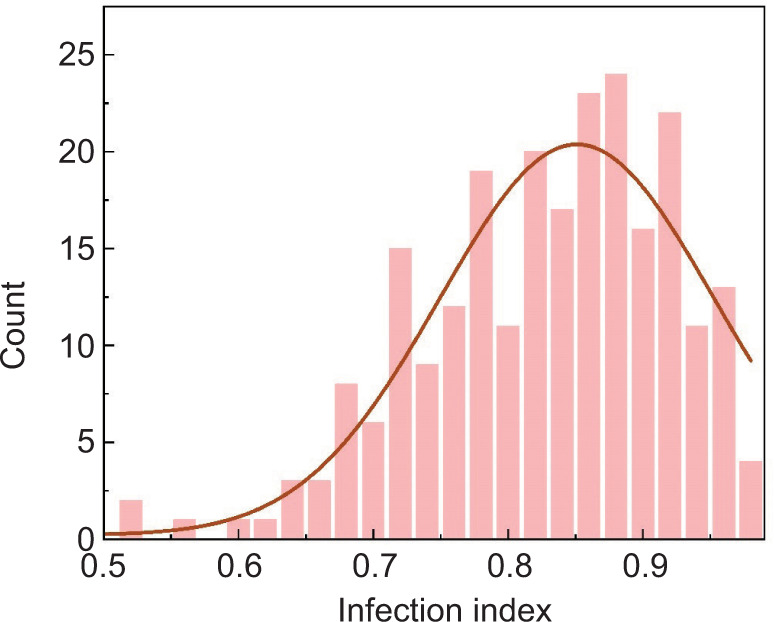
The infection index is a metric utilized to assess the severity of plant diseases, indicating the level of pathogen infection in plants. Certain peanut varieties exhibit strong disease resistance, which helps reduce the infection index by preventing pathogen attacks. Conversely, peanut varieties with low disease resistance are more vulnerable to disease infections, leading to a higher infection index. The infection index of the Xuhua 13 variety inoculated with AF2202 (0.91) was higher than that of the Zhonghua 6 variety (0.89). Among the 240 recombinant inbred populations, the infection index varied from 0.52 to 0.98, with significance values (5%, 50%, and 95%) and average values at 0.68, 0.84, 0.96, and 0.83, respectively (Table 1). The distribution of the infection index exhibited a left-skewed pattern (Figure 3).
Figure 3. The distribution frequency diagram of Xuhua 13 and Zhonghua 6 infection index.
Resveratrol distribution in peanuts RIL population
Analyzing the distribution of resveratrol in RIL peanut populations holds significant importance for disease resistance breeding, product development, and the efficient utilization of planting resources. Such analysis can enhance the peanut’s disease resistance, yield, and quality, offering substantial support for peanut production. The distribution of resveratrol, AFB1, AFB2, and the infection index in 240 family materials of the Xuhua 13/Zhonghua 6 recombinant inbred line F6 population is shown in Figure 4.
Figure 4. The distribution map of resveratrol content, AFB1, AFB2, and infection index in inbred lines of Xuhua 13 and Zhonghua 6.
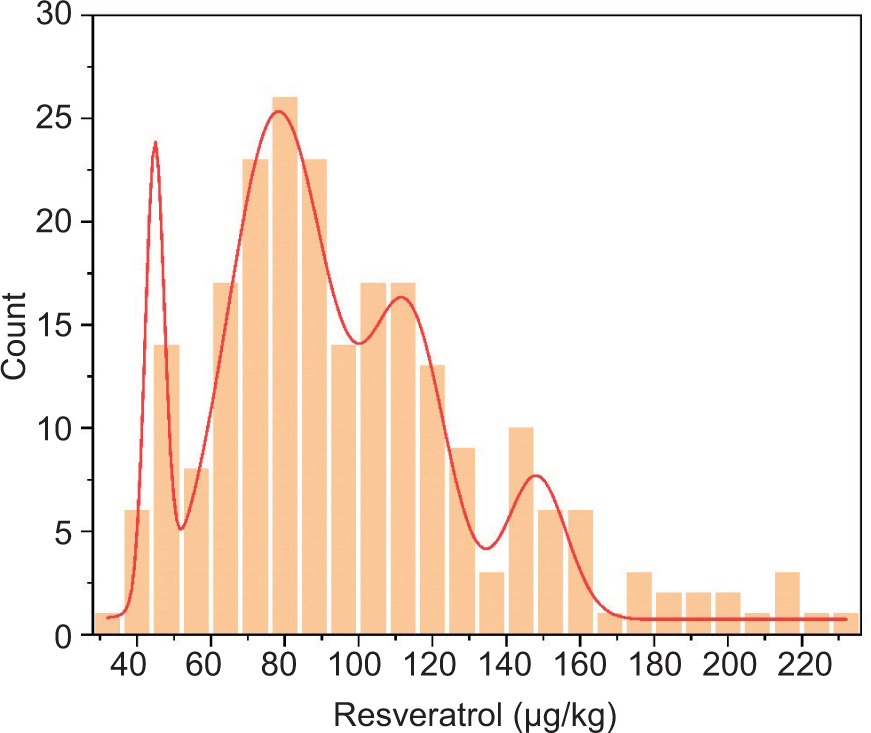
Resveratrol content exhibited a skewed distribution (Figure 5). Gaussian fitting results suggested that it was controlled by multiple genes. A total of 34% (82/240) of RIL individuals had higher resveratrol content than their male and female parents. The maximum resveratrol content in the RIL population was 867 μg/kg, 7.5 times higher than the male parent’s resveratrol content of 116 μg/kg and 19.7 times higher than the female parent’s resveratrol content of 44 μg/kg. Increased resveratrol content in peanuts can be achieved through hybrid breeding or molecular design breeding.
Figure 5. The distribution frequency diagram of Xuhua 13 and Zhonghua 6 resveratrol content.
Peanut resveratrol and its resistance to aflatoxin accumulation
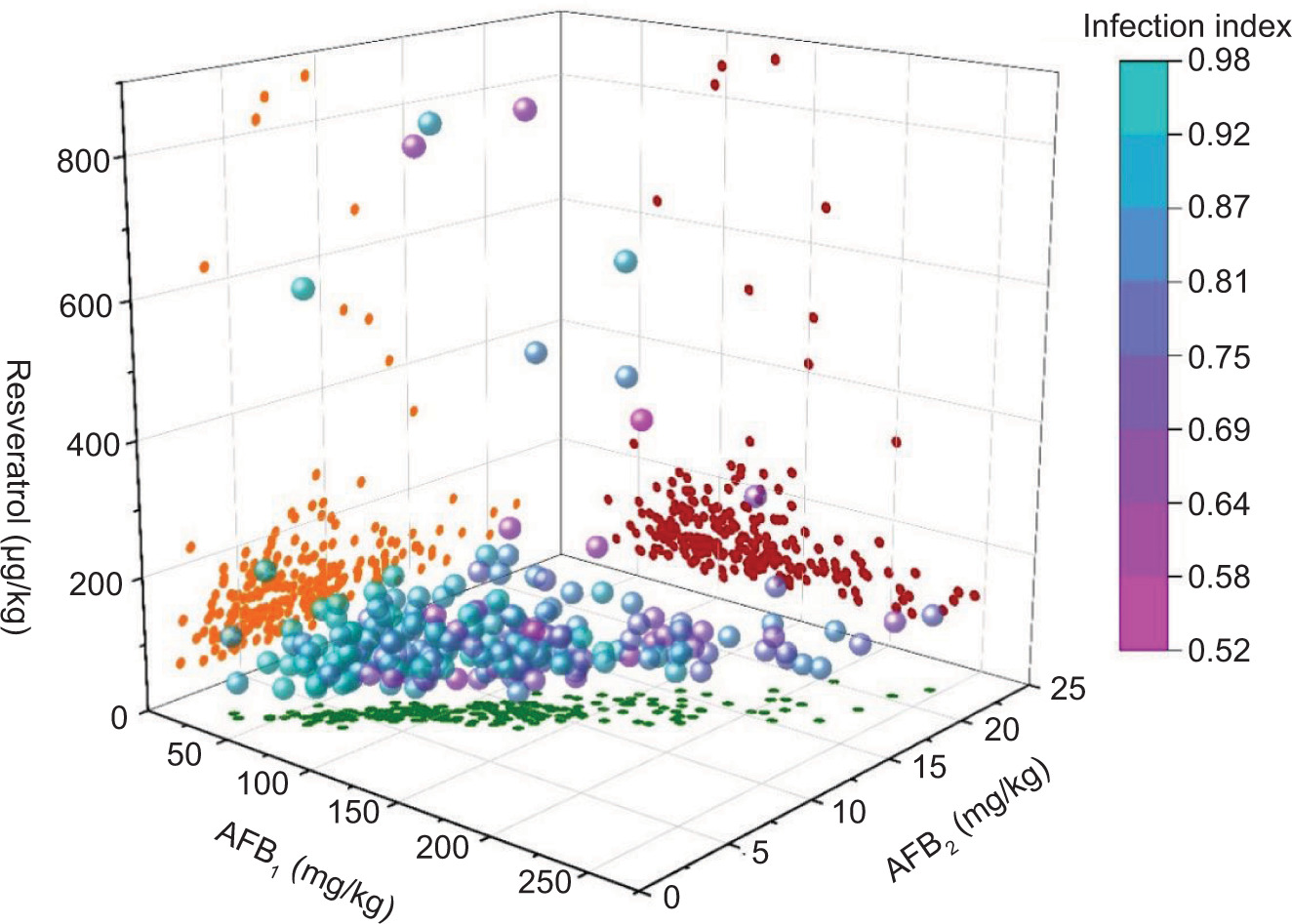
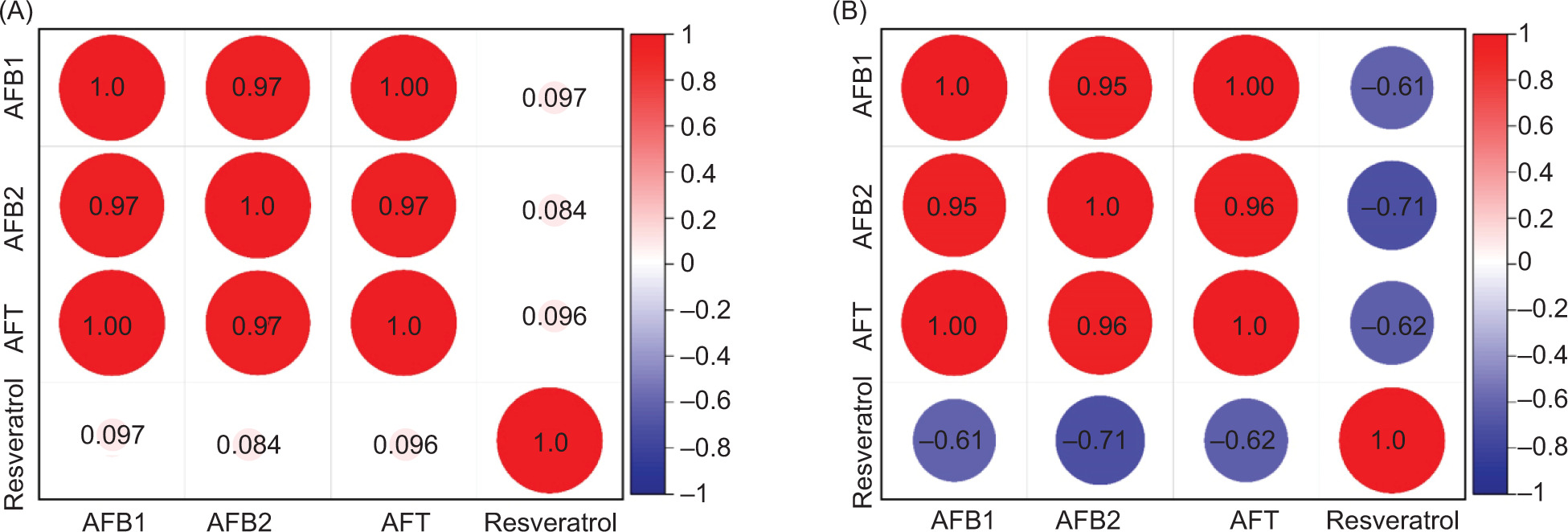
Exploring the correlation between resveratrol and aflatoxin accumulation in peanuts has the potential to enhance the development of peanut varieties with increased resistance to aflatoxin accumulation, decrease aflatoxin levels in peanut products, and improve both the safety and yield of peanuts. When the resveratrol content in peanuts fell below 300 μg/kg, no significant correlation was observed between the resveratrol and AFB1 content (Figure 6A). However, when the resveratrol content exceeded 300 μg/kg (Figure 6B), a substantial negative correlation with AFB1 and total aflatoxin contents was observed. This negative correlation is evident in the correlation coefficients of −0.61 and −0.62, respectively. In other words, higher levels of resveratrol in peanuts (>300 μg/kg) led to lower levels of AFB1 produced by peanut mildew. Some research suggests that resveratrol was efficacious in reducing and/or reversing the ZEA-induced toxicity (Virk et al., 2020).
Figure 6. The correlation between the content of resveratrol and AFB1, AFB2, and AFT [(A) Fell below 300 μg/kg; (B) Exceeded 300 μg/kg].
Peanut resveratrol and its resistance to Aspergillus flavus infection
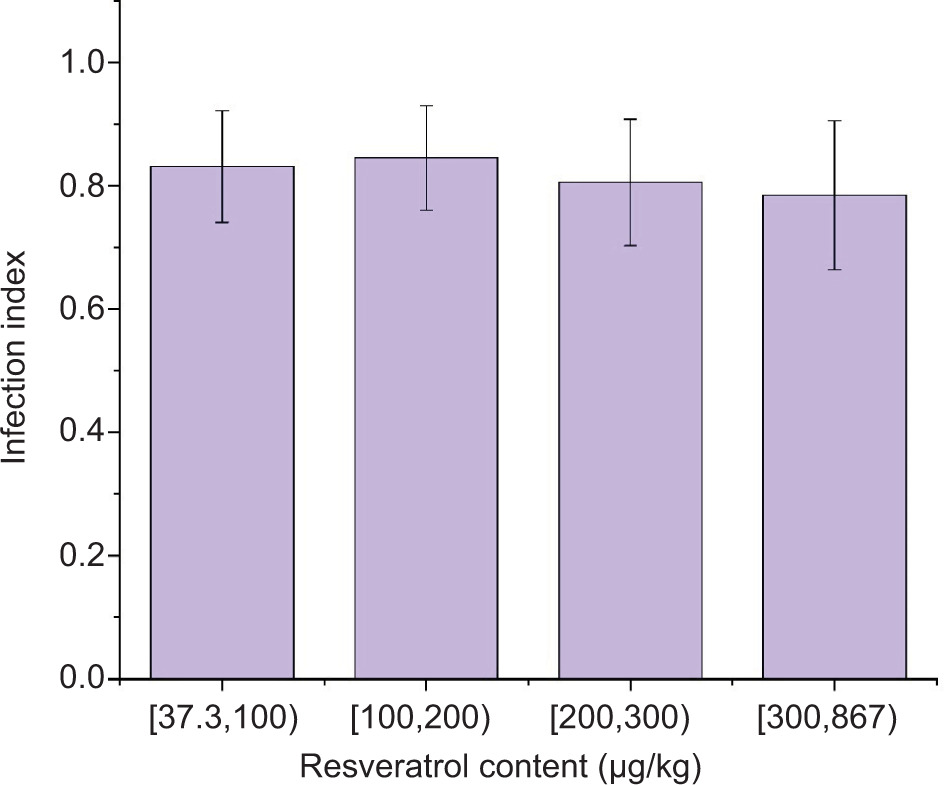
Little difference in the infection index of Aspergillus flavus was found among the 240 RIL populations with different levels of resveratrol (Figure 7), indicating that resveratrol did not inhibit the infection of Aspergillus flavus on peanuts (Wang et al., 2013). However, the high concentration of resveratrol exhibited an inhibitory effect on aflatoxin production. The CYP450 enzyme system is crucial in activating AFB1, which can induce and enhance the genotoxic effects of AFB1. Studies have shown that the mRNA expression of CYP genes was significantly reduced by resveratrol (Yang et al., 2022).
Figure 7. Analysis of the difference in infection index under different content of resveratrol.
Conclusions
This study addresses the issue of aflatoxin contamination in peanuts, aiming to enhance the quality and safety of this popular legume. The research utilized a peanut RIL population comprising 240 recombinant inbred lines to investigate the distribution of resveratrol in peanuts and its correlation with aflatoxin resistance. Findings indicate that resveratrol levels are influenced by multiple genes, with the highest content (867.0 µg/kg) observed in the recombinant inbred line population, surpassing that of the parent population. Notably, resveratrol content exceeding 300 µg/kg exhibited a significant negative association with aflatoxin B1 levels and total aflatoxin content, effectively inhibiting production. This suggests that peanut varieties rich in resveratrol possess resistance to aflatoxin contamination. The study advocates for the proactive development of aflatoxin-resistant peanut varieties, leveraging genetic enhancement techniques to bolster peanut disease resistance and mitigate aflatoxin risks. Furthermore, measures are recommended to suppress aflatoxin production during peanut harvesting, storage, and processing. These measures include improving storage conditions and optimizing processing techniques to uphold the quality and safety of peanut products.
Acknowledgments
Dr. Boshou Liao, Dr. Huifang Jiang and Dr Yong Lei provided experimental materials and technical guidance for this study.
Author Contributions
Conceptualization, Z.Z.; methodology, B.Y.; validation, X.H. and K.H.; formal analysis, W.L.; investigation, B.Y.; data curation, L.Y.; writing—original draft preparation, B.Y.; writing—review and editing, Z.Z.; supervision, Z.Z.; project administration and funding acquisition, P.L. All authors have read and agreed to the published version of the manuscript.
Conflict of Interest
The authors declare no conflict of interest.
Funding
This work was supported by the National Key Research and Development Program of China (2023YFD1400905), National Natural Science Foundation of China (32202172).
REFERENCES
Bian, Y., Wang, X., Zheng, Z., Ren, G., Zhu, H., Qiao, M., & Li, G. (2022). Resveratrol drives cancer cell senescence via enhancing p38MAPK and DLC1 expressions. Food & Function, 13(6), 3283–3293. 10.1039/D1FO02365A
Haiwen, C., Siliang, X., Jianbin, G., Weigang, C., Huaiyong, L., Nian, L., Li, H., Xiaojing, Z. et al. (2021). Identification and evaluation of resveratrol content in different peanut varieties. Chinese Journal of Oil Crop Sciences, 43(5), 942. 10.19802/j.issn.1007-9084.2020237
Chung, I.-M., Park, M. R., Chun, J. C., & Yun, S. J. (2003). Resveratrol accumulation and resveratrol synthase gene expression in response to abiotic stresses and hormones in peanut plants. Plant Science, 164(1), 103–109. 10.1016/S0168-9452(02)00341-2
Frémont, L. (2000). Biological effects of resveratrol. Life Sciences, 66(8), 663–673. 10.1016/S0024-3205(99)00410-5
Galiniak, S., Aebisher, D., & Bartusik-Aebisher, D. (2019). Health benefits of resveratrol administration. Acta Biochimica Polonica, 66(1), 13–21. 10.18388/abp.2018_2749
Guo, J., Chen, H., Liu, N., Chen, W., Zhou, X., Luo, H., Huang, L., Li, W., Wu, B., Huai, D., Lei, Y., Liao, B., & Jiang, H. (2022). Identification and validation of a major locus with linked marker for resveratrol content in culitivated peanut. Euphytica, 218(2), 15. 10.1007/s10681-022-02969-2
Jeandet, P., Douillet-Breuil, A. C., Bessis, R., Debord, S., Sbaghi, M., & Adrian, M. (2002). Phytoalexins from the Vitaceae: biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. Journal of Agricultural and Food Chemistry, 50(10), 2731–2741. 10.1021/jf011429s
Jing Dan, Y. X., Bai YiZhen, Guo Can, Ding XiaoXia, LI PeiWu, Zhang Qi. (2021). The Infectivity of Aspergillus flavus in Peanut. Scientia Agricultura Sinica, 54(23), 5008–5020. 10.3864/j.issn.0578-1752.2021.23.007
King, R. E., Bomser, J. A., & Min, D. B. (2006). Bioactivity of Resveratrol. Comprehensive Reviews in Food Science and Food Safety, 5(3), 65–70. 10.1111/j.1541-4337.2006.00001.x
Lee, S. S., Lee, S. M., Kim, M., Chun, J., Cheong, Y. K., & Lee, J. (2004). Analysis of trans-resveratrol in peanuts and peanut butters consumed in Korea. Food Research International, 37(3), 247–251. 10.1016/j.foodres.2003.11.007
Li, W., Liu, N., Huang, L., Chen, Y., Guo, J., Yu, B., Luo, H., Zhou, X., Huai, D., Chen, W., Yan, L., Wang, X., Lei, Y., Liao, B., & Jiang, H. (2022). Stable major QTL on chromosomes A07 and A08 increase shelling percentage in peanut (Arachis hypogaea L.). The Crop Journal, 10(3), 820–829. 10.1016/j.cj.2021.09.003
Luo, X., Wu, S., Jia, H., Si, X., Song, Z., Zhai, Z., Bai, J., Li, J., Yang, Y., & Wu, Z. (2022). Resveratrol alleviates enterotoxigenic Escherichia coli K88-induced damage by regulating SIRT-1 signaling in intestinal porcine epithelial cells. Food & Function, 13, 7346–7360. 10.1039/d1fo03854k
Marshall, H., Meneely, J. P., Quinn, B., Zhao, Y., Bourke, P., Gilmore, B. F., Zhang, G., & Elliott, C. T. (2020). Novel decontamination approaches and their potential application for post-harvest aflatoxin control. Trends in Food Science & Technology, 106, 489–496. 10.1016/j.tifs.2020.11.001
Meneely, J. P., Kolawole, O., Haughey, S. A., Miller, S. J., Krska, R., & Elliott, C. T. (2023). The Challenge of Global Aflatoxins Legislation with a Focus on Peanuts and Peanut Products: A Systematic Review. Exposure and Health, 15(2), 467–487. 10.1007/s12403-022-00499-9
Mingrou, L., Guo, S., Ho, C.-T., & Bai, N. (2022). Review on chemical compositions and biological activities of peanut (Arachis hypogeae L.). Journal of Food Biochemistry, 46(7), e14119. 10.1111/jfbc.14119
Mwakinyali, S. E., Ding, X., Ming, Z., Tong, W., Zhang, Q., & Li, P. (2019). Recent development of aflatoxin contamination biocontrol in agricultural products. Biological Control, 128, 31–39. 10.1016/j.biocontrol.2018.09.012
Ogaki, Y., & Sagawa, I. (2003). Trans-resveratrol content in seeds and seed coats of Japanese peanut cultivars and that in peanut products. Nippon Shokuhin Kagaku Kogaku Kaishi, 50(12), 570–573. 10.3136/nskkk.50.570
Qin-fei, W., Qiang, X. U., Ru-lian*, Z., Di-fa, L. I. U., Dong-shan, Y., Ming, W., & Li-ping, L. I. (2015). Resveratrol and its metabolites accumulation responding to abiotic stresses and hormones in peanut seedlings. Chinese Journal of Oil Crop Sciences, 37(3), 301. 10.7505/j.issn.1007-9084.2015.03.007
Ros, E. (2010). Health benefits of nut consumption. Nutrients, 2(7), 652–682. 10.3390/nu2070652
Sales, J. M., & Resurreccion, A. V. (2014). Resveratrol in peanuts. Critical Reviews in Food Science and Nutrition, 54(6), 734–770. 10.1080/10408398.2011.606928
Sanders, T. H., McMichael, R. W., Jr., & Hendrix, K. W. (2000). Occurrence of resveratrol in edible peanuts. Journal of Agricultural and Food Chemistry, 48(4), 1243–1246. 10.1021/jf990737b
Toomer, O. T. (2018). Nutritional chemistry of the peanut (Arachis hypogaea). Critical Reviews in Food Science and Nutrition, 58(17), 3042–3053. 10.1080/10408398.2017.1339015
Virk, P., Al-mukhaizeem, N. A. R., Bin Morebah, S. H., Fouad, D., & Elobeid, M. (2020). Protective effect of resveratrol against toxicity induced by the mycotoxin, zearalenone in a rat model. Food and Chemical Toxicology, 146. 10.1016/j.fct.2020.111840
Wang, H., Huang, J., Lei, Y.-j., Yan, L., Wang, S.-Y., Jiang, H., Ren, X., Lou, Q., & Liao, B. (2013). Relationship of Resveratrol Content and Resistance to Aflatoxin Accumulation Caused by Aspergillus flavus in Peanut Seeds. Acta Agronomica Sinica, 38, 1875–1883.
Wang, H., Lei, Y., Yan, L., Wan, L., Ren, X., Chen, S., Dai, X., Guo, W., Jiang, H., & Liao, B. (2016). Functional Genomic Analysis of Aspergillus flavus Interacting with Resistant and Susceptible Peanut. Toxins, 8(2), 46. c10.3390/toxins8020046
Wang, M. L., & Pittman, R. N. (2009). Resveratrol content in seeds of peanut germplasm quantified by HPLC. Plant Genetic Resources, 7(1), 80–83. 10.1017/S1479262108048247
Wu, W., Li, K., Ran, X., Wang, W., Xu, X., Zhang, Y., Wei, X., & Zhang, T. (2022). Combination of resveratrol and luteolin ameliorates α-naphthylisothiocyanate-induced cholestasis by regulating the bile acid homeostasis and suppressing oxidative stress. Food & Function, 13(13), 7098–7111. 10.1039/D2FO00521B
Yang, H., Wang, Y. J., Yu, C. T., Jiao, Y. H., Zhang, R. S., Jin, S. J., & Feng, X. J. (2022). Dietary Resveratrol Alleviates AFB1-Induced Ileum Damage in Ducks via the Nrf2 and NF-κB/NLRP3 Signaling Pathways and CYP1A1/2 Expressions. Agriculture, 12(1). 10.3390/agriculture12010054
Yu, B., Liu, N., Huang, L., Luo, H., Zhou, X., Lei, Y., et al. (2023). Identification and application of a candidate gene AhAftr1 for aflatoxin production resistance in peanut seed (Arachis hypogaea L.). Journal of Advanced Research. 10.1016/j.jare.2023.09.014