
Download
Proliferation rate phagocytosis rate = A 1 A 0 × 100 1 Inhibitation rate = 1 − A 1 A 0 h 2 



Original Article
Evaluation of active components and pharmacological activities by UPLC-Q/TOF-MS and network pharmacology in Lentinula edodes and Lyophyllum decastes
Yingnan Zhang, Gaoxing Ma, Fei Pei, Ning Ma, Anxiang Su, Qiuhui Hu, Meng Wang*
College of Food Science and Engineering, Nanjing University of Finance and Economics/Collaborative Innovation Center for Modern Grain Circulation and Safety, Nanjing, China
Abstract
Lentinula edodes (L. edodes) is a premium edible fungus cultivated extensively throughout China, and Lyophyllum decastes (L. decastes) is a wild edible fungus with a high degree of industrial-level cultivation in China. Regarding the edible value of the two fungi, research on their medicinal properties is relatively scarce. In this study, we utilized ultra-high performance liquid chromatography with quadrupole and time-of-flight mass spectrometry to analyze comprehensively the chemical composition of both fungi. In all, 120 and 144 chemical components were identified in L. edodes and L. decastes, respectively. Subsequently, we elaborated active components, potential biological activities, and therapeutic targets associated with both fungi by using network pharmacology. The results showed that seven active components in L. edodes and nine active components in L. decastes could act on common targets, such as AKT serine/threonine kinase1 and peroxisome proliferator-activated receptor gamma, thereby regulating signaling pathways, such as epidermal growth factor receptor and mitogen-activated protein kinase, to affect the body. In addition, these components affect immune regulation and inhibition of liver cancer. Finally, we used cell experiments to verify their activities. Our findings provided a reliable foundation for future research aimed at harnessing the bioactivities of L. edodes and L. decastes, thereby offering novel strategies for their utilization in the field of functional foods and natural medicine.
Key words: edible fungi, active components, pharmacological activities, UPLC-Q/TOF-MS, network pharmacology
*Corresponding Author: Meng Wang, College of Food Science and Engineering, Nanjing University of Finance and Economics/Collaborative Innovation Center for Modern Grain Circulation and Safety, Nanjing 210023, China. Email: [email protected]
Academic Editor: Teresa D’Amore, PhD, Chemistry Department, Experimental Zooprophylactic Institute of Puglia and Basilicata, Via Manfredonia 20, 71121 Foggia, Italy
Received: 28 March 2024; Accepted: 14 October 2024; Published: 21 December 2024
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Lentinula edodes (L. edodes) is a widely consumed fungus classified under the taxonomic hierarchy of fungi, Basidiomycota, Agaricomycetes, Agaricales, Omphalotaceae, and Lentinula. Its natural distribution includes warm and moist climates of Southeast Asia (Roszczyk et al., 2022). Presently, it is found all over the world because of its artificial planting. L. edodes is rich in polysaccharides, dietary fiber, protein, and other nutrients, and has been used as an edible source of food for thousands of years in Asian and European countries. L. edodes is also considered a medicinal fungus in traditional Chinese medicine (TCM); its pharmacological effects have long been mentioned in TCM classics.
Lyophyllum decastes (L. decastes) is a wild edible and medicinal fungus classified within the taxonomic hierarchy of fungi, Basidiomycota, Agaricomycetes, Agaricales, Lyophyllaceae, and Lyophyllum. China is the largest producer of L. decastes globally, and its 2020 yield in China was about 11,590 tons (Xu et al., 2023). L. decastes has delicate flesh, fragrant aroma, delicious taste, and is rich in proteins, minerals, and polysaccharides. It is recorded in Chinese Materia Medica that L. decastes can stop bleeding, detoxify, and treat breast cancer and hemangiomas (Lin, 2012).
Several studies have reported the biological activities of the crude extracts or related products of both fungi in vivo as well as in vitro. Hye-Lim et al. (2017) estimated antioxidant activities of different extracts of L. edodes. Among them, water extract exhibited the strongest 2,2-Diphenyl-1-picrylhydrazyl (DPPH), 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid positive (ABTS+) radicals, and nitrite scavenging activities. Hot water extracts of L. decastes can induce the production of interferon-gamma (IFN-γ) and interleukin-4 (IL-4) in mice and enhance T cells immune activity (Ike et al., 2012). Nevertheless, these studies assessing the biological activities of fungi were not exhaustive, as they merely provided a brief evaluation of their primary component and its specific active functions (Dai et al., 2023a).
Many studies demonstrated different effects of various bioactive components of edible fungi on the body, and these components are often multifunctional in biological activity (Li et al., 2022). They have multiple targets through which they regulate biological processes and signaling pathways of the body (Garcia et al., 2022). Phytochemical profile analysis is a key step in the development and utilization of plant resources and quality safety assurance. So far, research on the composition of the two fungi has mainly involved the nutritional composition (Gameli-Kwabla, 2020; Nayik et al., 2023), polysaccharides (Dai et al., 2023b; Zhang et al., 2023), and volatile components (Fujita et al., 2021; Wang et al., 2021) of the fruiting body, mycelium and fermentation broth. Literature reports also described the small-molecule chemical composition of both fungi. A number of techniques have been developed to analyze the composition of fungi, such as headspace solid-phase microextraction, gas chromatography–mass spectrometry, electronic nose for volatile substances (Lu et al., 2022), and high-performance liquid chromatography (HPLC), and nuclear magnetic resonance (NMR) for other substances (Ding et al., 2022).
Immune system diseases (ISD), also known as immunological disorders, arise from dysfunctions within the immune system, which is responsible for maintaining homeostasis and safeguarding the health. These ISDs result in the immune system erroneously attacking its own tissues and molecules, leading to a variety of debilitating conditions (Wen et al., 2024). Virtually, all pathological states in organ tissues are intricately linked to the immune system. Infections, autoimmunity, immune deficiencies, antigen responses, cancer development, and various other conditions are intricately connected to the functioning of the immune system. Alternatively, cancer disease (CD) is often referred to as the “king of all diseases” because of its ability to affect nearly every part and organ of the human body. Cancer is the first or second leading cause of premature deaths (at ages 30–69 years) globally (Xie et al., 2021). According to the World Health Organization (WHO), cancer is one of the main causes of death in humans, with 10 million (nearly one in six) deaths in 2020 attributed to cancer. There exists an interactive and complex relationship between ISDs and cancer. In addition to predisposing individuals to ISDs, an imbalance in the immune system may also facilitate the evasion of cancer cells from detection and foster their growth within the body (Li et al., 2021). In contrast, cancer cells possess diverse mechanisms to interfere the immune system, thus promoting immune evasion. Consequently, patients often exhibit signs of immunosuppression, rendering them more vulnerable to infections with other pathogens (Starzer et al., 2022).
Many studies have shown diverse health effects of fungi because of the presence of a variety of bioactive components (Cui et al., 2022; Yin et al., 2022). Ultra-high performance liquid chromatography with quadrupole and time-of-flight mass spectrometry (UPLC-Q–TOF-MS), with the advantages of high speed, high resolution, and high precision, is a mass spectrometry technology used to integrate separation and detection analysis. The technology is extensively employed in investigating the chemical components and active constituents of natural medicines (Liang et al., 2021). It also disintegrates the metabolic kinetics of pharmacodynamic components combined with network pharmacology (Qiu et al., 2024). Network pharmacology is a combined technology of computer application and systems biology, used to explore the complex network of interactions between drugs and biological systems. This approach helps to understand the multi-target mechanisms of drugs, their adverse effects, and potential new uses (Zhu et al., 2023). Du et al. (2024) confirmed that phosphoinositide 3-kinase–protein kinase B (PI3K/AKT) signaling pathway could probably be the mechanism by which pachymic acid could effectively treat gastric cancer by network pharmacology. Xu et al. (2024) used network pharmacology to confirm that the mechanism of Res against glycogenin (GN) could be the regulation of arachidonic acid metabolism by regulating prostaglandin-endoperoxide synthase 1 (PTGS1) and PTGS2.
For better utilization of L. edodes and L. decastes resources, it is crucial to conduct an exhaustive analysis of their chemical constituents and biological properties. The primary objective of this study was to conduct a comprehensive assessment of the chemical composition and bioactivity of both fungi. Owing to the lack of systematic analysis of the components and complex pharmacological effects of both edible fungi, this study aimed to elucidate through cell experiments the principal active constituents present in two fungi, along with identifying their pharmacodynamic targets. Such insights are pivotal for validating their clinical applications and fostering the continued development of both fungal resources.
Materials and Methods
Analysis through UPLC-Q–TOF-MS
Chemicals and reagents
Chromatography-grade methanol, acetonitrile, and formic acid were purchased from Sigma-Aldrich (St. Louis, MO, USA). Deionized water (18.3 MΩ) was generated by a Milli-Q water purification system obtained from Millipore Ltd. (Bedford, MA, USA). Macrophages RAW264.7 were purchased from Pricella Biotechnology Co. (Wuhan, China). The liver cancer cells HepG-2 were purchased from Beyotime Biotechnology (Nanjing, China). Dulbecco’s Modified Eagle Medium (DMEM), penicillin/streptomycin, and phosphate-buffered saline (PBS) solution were purchased from Thermo Fisher Scientific Inc. (Nanjing, China). CCK-8 kit and neutral red kit were purchased from Beyotime Biotechnology. L. edodes and L. decastes were collected from Yunnan Bacteria Horizon Biotechnology Co. (Yunnan, China).
Preparation of Samples
In all, 1.0 g of fungi freeze-dried material was weighed, placed in a 50-mL centrifuge tube, and extracted with 20 mL of methanol–water (1:1, v/v). The solution was vortexed for 1 min, ultrasonicated at room temperature for 30 min, centrifuged at 4°C with 12,000 r/min for 10 min at 4°C; 1 mL of the supernatant was filtered using a 0.22-µm nylon membrane.
Instrumentation
Sepax GP-C18 column (1.8 µm, 120 Å, 2.1 mm × 150 mm). Each component was gradient eluted for 21 min. The mobile phase was (a) 0.01% formic acid and (b) 100% acetonitrile. The flow rate was maintained at 0.3 mL/min, with column temperature of 40°C and an injection volume of 2 µL (Table 1). Electrospray ionization (ESI) with positive ion and negative ion modes was used for detection. ESI source conditions were as follows: ion spray voltage 5,500 V in positive ion mode, gas 50 psi, temperature 500°C; negative ion mode 4,400 V, gas 50 psi, temperature 450°C; curtain gas 25 psi; time-of-flight–mass spectrometry (TOF-MS) scanning range: 100–1,200 Da; ion scanning range: 50–1,000 Da; TOF/MS scan accumulation time 0.2 s; product ion scan accumulation time 0.01 s; secondary mass spectrometry using high sensitivity mode and information-dependent acquisition, declustering voltage ±60 V, and collision energy 35±15 eV.
Table 1. Gradient elution procedure conditions.
| Elution time(min) | Phase A (%) | Phase B (%) |
|---|---|---|
| 0 | 95 | 5 |
| 10 | 30 | 70 |
| 17 | 0 | 100 |
| 18 | 0 | 100 |
| 19 | 95 | 5 |
| 21 | 95 | 5 |
Database search and comparison of fungi components
The mass spectrometry acquisition .wiff file was preprocessed by the software MS-DIAL 4.70, including peak extraction, denoising, deconvolution, peak alignment, and export of a three-dimensional (3D) data matrix in comma-separated value (CSV) format. The extracted peak information was compared with the database GNPS (https://gnps.ucsd.edu/), Respect (http://spectra.psc.riken.jp/), and MassBank (https://massbank.eu/MassBank). The databases were searched in their entirety and matched according to the parameters shown in Table 2. This 3D matrix includes the following information: sample information, retention time, mass-to-nuclear ratio, and mass spectrometry response intensity.
Table 2. Database matching parameters.
| Classification | Parameters | Setting |
|---|---|---|
| Peak detection parameters | Minimum peak height | 1,000 amplitude |
| Mass slice width | 0.1 Da | |
| Alignment parameters | Retention time tolerance | 0.05 min |
| MS1 tolerance | 0.015 Da | |
| Identification setting | Accurate mass tolerance (MS1) | 0.01 Da |
| Accurate mass tolerance (MS2) | 0.05 Da | |
| Identification score cut off | 80 |
Analysis of network pharmacology
Target prediction of L. edodes and L. decastes
First, the active components of L. edodes and L. decastes were screened by the SwissADME (http://www.swissadme.ch/) platform and literature. At SwissADME platform, gastrointestinal absorption (GA), one of the pharmacokinetic parameters, was set at “HIGH” as a condition for drug absorption, and active compounds with good oral bioavailability were screened. Likewise, the drug-likeness (DL) was also considered for cosmeceutical parameters (Lipinski, Ghose, Veber, Egan, and Muegge); two or more of them with “YES” can be regarded as active components. Second, SwissTarget Prediction platform (http://www.swisstargetprediction.ch/) was applied to predict possible targets. SwissTarget Prediction selected the targets whose bioavailability score (BS) was more than 0.85 in the prediction results for further analysis. At the same time, experimentally verified targets information was downloaded from SwissTarget Prediction, and the entries related to the active components of L. edodes and L. decastes were extracted. Finally, target information was integrated and accumulated to obtain the possible targets of L. edodes and L. decastes active components.
Prediction o targets of two diseases
Data for all-associated disease targets were acquired from the GeneCards (https://www.genecards.org/) database and Biotechnology Information database (https://www.ncbi.nlm.nih.gov/) (National Center for Biotechnology Information [NCBI]) using “Immune system diseases,” “Cancer diseases” and their synonyms. The above targets were converted and queried into the UniProt ID format with “Homo sapiens” as the qualifying condition in the UniProt database. Finally, the gene library of all targets was established by eliminating repeated targets.
Intersection between active compounds and disease targets
The intersection targets between the disease genes and the predicted L. edodes and L. decastes targets were obtained. Jvenn website (https://jvenn.toulouse.inra.fr/app/example.html) was used to construct a Venn diagram for visualization.
Protein–protein interaction network construction
The above intersection targets were imported into the STRING database (http://string-db.org) for protein interaction network analysis. The screening condition of the species was set to “Homo sapiens” and the minimum required interaction score was “highest confidence (0.9).” Input protein–protein interaction (PPI) information into Cytoscape 3.10.1 for visualization and constructs network of potential key targets.
Topological and cluster analyses of the protein–protein interaction network
The CytoHubba plugin in Cytoscape (https://www.cytoscape.org/) was used to identify hub genes. Three critical topological parameters were chosen for screening core composite targets based on the PPI network: degree, betweenness, and closeness. Values for the three parameters indicated the significance as well as the impact of relevant nodes in the entire network. The MCODE plug-in in Cytoscape was used to screen PPI network modules using various cut-offs: betweenness centrality, betweenness centrality, and degree are greater than the median.
Gene ontology and Kyoto encyclopedia of genes and genomes enrichment analyses
The above-mentioned intersection targets were subjected to the gene ontology (GO) biological process analysis and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways enrichment analyses using the DAVID database (https://david.ncifcrf.gov/). R version 3.10.1 was used to visualize the results.
Construction of active compound-target network
For visualization, potential active components and matching intersection targets were imported into the Cytoscape 3.10.1 software and a network of compound-target network was built. Each component of targets is represented by nodes, and the relationship between the components, diseases, and targets is represented by connecting lines.
Culture of cells
Macrophages RAW264.7 and liver cancer cells HepG-2 were cultured. Macrophage RAW264.7 cells were cultured in a constant temperature incubator using a prepared complete medium (DMEM containing 10% fetal bovine serum [FBS] and 1% penicillin/streptomycin). After 48 h of cell culture, the cells adhered to the wall and grew into a monolayer of cells before passage. During passage, the culture medium was discarded, 1 mL of sterile PBS was added to wash twice and then the PBS was discarded. Complete medium, 1 mL, was used to blow until the cells no longer adhered to the wall, and then added to the culture flask containing 6 mL of complete medium prepared in advance. The difference between HepG-2 culture and RAW 264.7 is that the complete culture medium contains 15% FBS and 1% penicillin/streptomycin, and trypsin digestion is required during sub-culturing.
Determination of cell viability
The proliferation and phagocytic proportions of RAW264.7 and the inhibition proportion of HepG-2 were measured. The proliferation and inhibition proportions were determined as follows: (Formula 1), after freeze-drying, a certain amount of DMEM was added to prepare the mother solution, and after ultrasonication, the mother solution was filtered through a 0.22-μm microporous filter membrane for sterilization, and the DMEM was added continuously to dilute the mother solution to obtain different concentrations. RAW264.7 or HepG-2 in the logarithmic growth phase were inoculated on a 96-well plate at a density of 1×105/mL, 200 μL per well; the culture medium was discarded after 24 h of culture, and 200 μL of culture medium solution of different groups was added for 24 h. Using the CCK-8 kit, 20 μL of CCK-8 solution was added to each well and incubated for 1 h, and the absorbance value was measured at 450 nm using an enzyme-linked-immunosorbent serologic assay (ELISA) reader:
The difference in determining phagocytosis rate is that the above CCK-8 kit is replaced with a neutral red kit (Formula 2):
Statistical analysis
Data were mainly obtained by downloading from public databases. In the cell experiments, each sample with different concentrations was repeated for six times to avoid accidental errors. Data were considered statistically significant with P < 0.05.
Results
Targeted analysis of chemical components in L. edodes and L. decastes
The mass spectrometry results of both fungi were analyzed and compared with the database, which had 120 components of L. edodes and 144 components of L. decastes (Supplementary Tables S1 and S2 and Supplementary Figure S1). The components with higher contents in the two extracts were carbohydrates, proteins, and fatty acids. Among them, carbohydrates in L. edodes accounted for 22.1% and proteins for 16.1%. Carbohydrates in L. decastes accounted for 22.4% and proteins 17.3%. The contents of proteins and polysaccharide were the same as reported in edible fungi (Zhou et al., 2023). In addition, L. edodes had a higher content of purine and L. decastes had a higher content of some organic bases (Figure 1).
Figure 1. Component content identified by UPLC-Q–TOF-MS. (A) Components of L. edodes and (B) components of L. decastes.
Analysis of biological activities of L. edodes and L. decastes
As raw materials of functional foods, L. edodes and L. decastes are always consumed wholly. They have complex systems with many types of components; hence, the bioactivity evaluation and mechanism study of a single component cannot truly reflect the overall physiological effects of L. edodes and L. decastes (Zhang et al., 2022). Here, the potential bioactivity of both fungi was systematically predicted by enrichment analysis of their active components and their corresponding targets (Figure 2). SwissADME web tool database was used to screen and evaluate gastrointestinal absorption and their drug-like properties. This led to the identification of 60 types of medicinal components in L. edodes and 73 types in L. decastes that met the identification conditions. Further, medicinal components with a bioavailability > 0.85 were selected for further screening, resulting in seven key medicinal components in L. edodes and nine in L. decastes (Table 3). Many types of fatty acids among key medicinal components were screened. Fatty acids have beneficial health effects, particularly in the prevention of cardiovascular diseases, inflammation, and metabolic disorders, such as diabetes (Coudray et al., 2021). Fatty acids exert significant immune activity by modulating immune cells and regulating intestinal flora (Sganzerla et al., 2022).
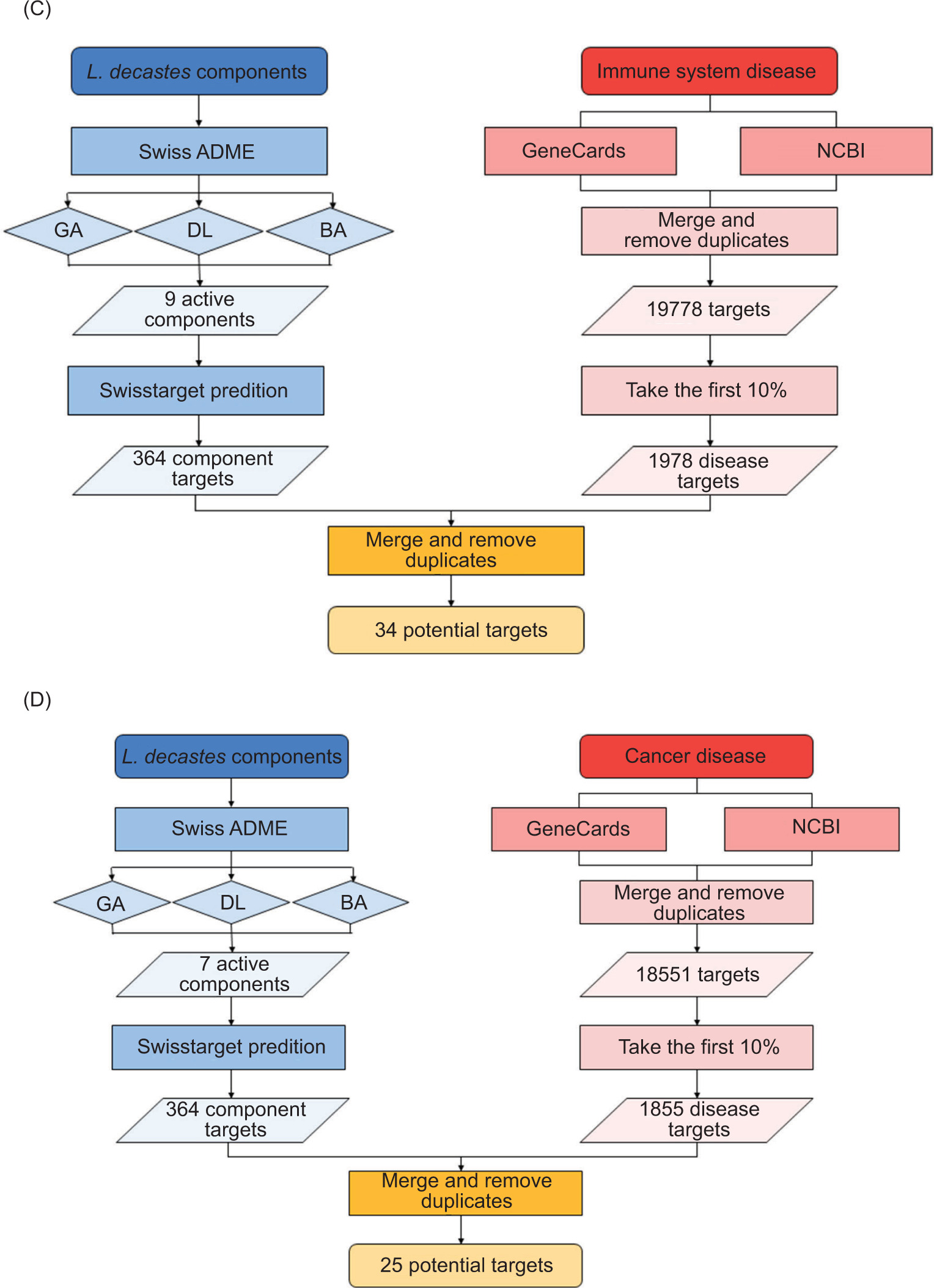
Figure 2. Flow chart for the prediction of targets of active components from fungi and diseases by network pharmacological analysis. (A) Active components of L. edodes and ISD; (B) active components of L. edodes and CD.
Table 3. Active components of L. edodes and L. decastes.
| L. edodes | L. decastes | ||
|---|---|---|---|
| No. | Active components | No. | Active components |
| 1. | FA 18:2+2O | 1. | FA 18:2+2O |
| 2. | L-5-Oxoproline | 2. | FA 18:3+2O |
| 3. | Propanoic acid | 3. | FA 18:1+1O |
| 4. | FA 18:4+2O | 4. | Succinic acid |
| 5. | Lapachol | 5. | 9Z,12Z-linoleic acid |
| 6. | Sebacic acid | 6. | Maleic acid |
| 7. | 10-Hydroxydecanoic acid | 7. | 16-Hydroxyhexadecanoic acid |
| 8. | 3-Methyladipic acid | ||
| 9. | Ketoisovaleric acid | ||
In addition, potential targets that satisfied the conditions were obtained. After merging and de-duplicating the potential targets predicted by SwissTarget Prediction, a total number of L. edodes and L. decastes targets were calculated, yielding 383 therapeutic targets for key medicinal components of L. edodes and 364 therapeutic targets for key medicinal components of L. decastes. After collating and removing duplicates, 1,978 ISD gene targets and 1,855 CD gene targets were identified. Through jvenn website, 31 types of intersection targets between L. edodes and ISD were discovered, while 27 types associated with CD were also identified. Similarly, 34 types of intersection targets between L. decastes and ISD were established, while 25 types associated with CD were also identified. The related targets of these two diseases were queried from GeneCards database and NCBI (Figure 2).
Figure 2. Flow chart for the prediction of targets of active components from fungi and diseases by network pharmacological analysis. (C) active components of L. decastes and ISD; and (D) active components of L. decastes and CD.
Components act on multiple targets to treat diseases. In general, the components of L. edodes and L. decastes act on 12 targets, such as glycogen synthase kinase 3 beta (GSK3B), telomerase reverse transcriptase (TERT), and AKT serine/threonine kinase (AKT1), for immune regulation, and 13 targets, such as kinase insert domain receptor (KDR), peroxisome proliferator-activated receptor alpha (PPARA), androgen receptor (AR), etc., to affect liver tumors. For example, tryptophan 2,3-dioxygenase 2 can upregulate IL-8 by phosphorylating AKT–GSK3B and promote the polarization of M2 macrophages. The activation of peroxisome proliferator-activated receptor gamma (PPARG) can regulate the cell cycle distribution of colon cancer cells and promote cell apoptosis (Selenz et al., 2022). In addition, the two edible fungi components can simultaneously act on 10 targets, such as GSK3B, AKT1, and PTGS2, to affect immune function and liver tumor development. This shows that some targets play a wide range of roles when participating in biological activities, which could be a key point in disease regulation.
Gene set enrichment analysis of GO and KEGG
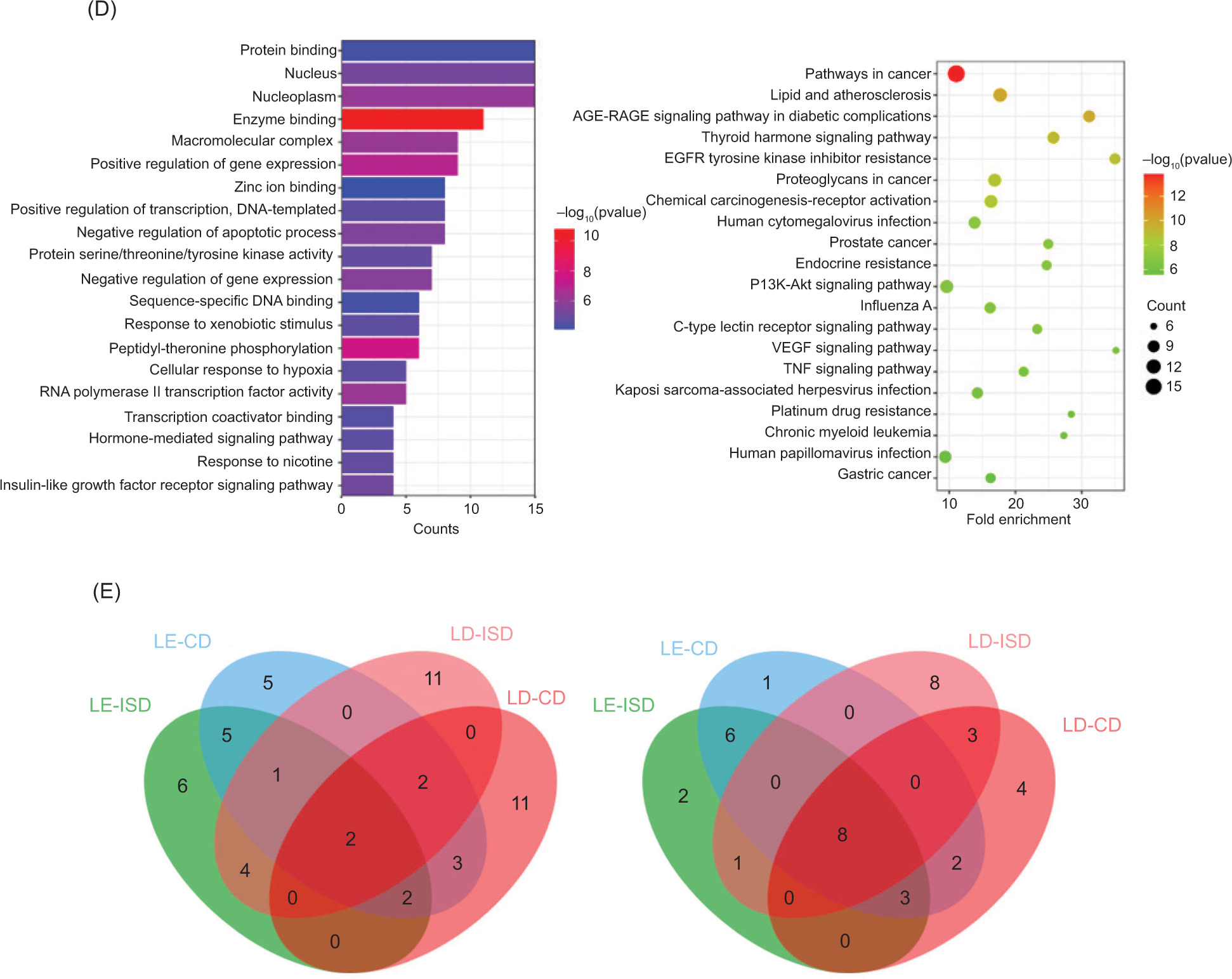
Finally, further enrichment analysis was conducted to analyze overlapping targets and the detailed results. Similar to the results obtained for the targets, several identical GO biological processes and KEGG pathways were observed in the first four panels (Figure 4), indicating that these represent key biological processes and pathways for treatment of diseases by fungi. Enrichment analysis of seven active components of L. edodes yielded 31 core targets related to immunity. In all, 309 GO biological processes, 99 KEGG signaling pathways and 27 core targets related to cancer were obtained. Enrichment analysis of 305 GO biological processes and 96 KEGG signaling pathways were obtained. Correspondingly, enrichment analysis of nine types of active components of L. decastes yielded 34 core targets related to immunity, and 300 GO biological processes, 135 KEGG signaling pathways, and 25 core targets related to cancer. Enrichment analysis resulted in 218 GO biological processes and 137 KEGG signaling pathways.
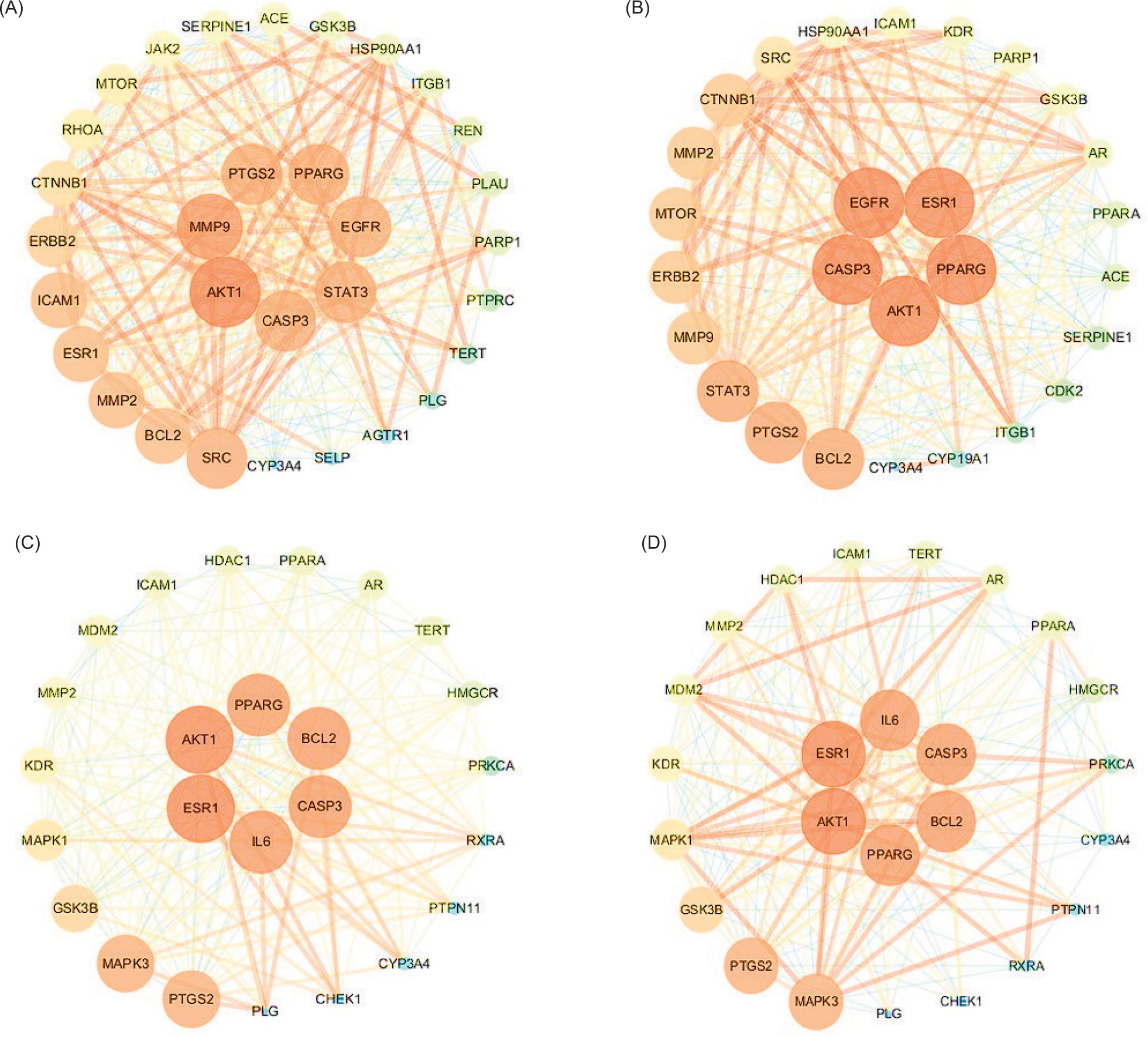
Figure 3. Interaction network diagram of core pharmacodynamic targets of fungi components for treating diseases. (A) L. edodes components and ISD; (B) L. edodes components and CD; (C) L. decastes components and ISD; and (D) L. decastes components and CD.
Figure 4. The GO and KEGG enrichment analysis of core pharmacodynamic targets. (A) L. edodes components and ISD; (B) L. edodes components and CD; (C) L. decastes components and ISD.
For two diseases, the GO enrichment analysis revealed that the targets of L. edodes were always closely related to 10 biological processes, which were nitric oxide (NO) synthase regulator activity, protein phosphatase binding, protein autophosphorylation, positive regulation of protein kinase B signaling, estrogen receptor binding, positive regulation of apoptotic process, enzyme binding, response to xenobiotic stimulus, negative regulation of gene expression, and negative regulation of apoptotic process, The KEGG pathway enrichment analysis indicated that the targets of L. edodes were always significantly enriched in 17 pathways, which were prostate cancer, colorectal cancer, fluid shear stress and atherosclerosis, chemical carcinogenesis–receptor activation, diabetic cardiomyopathy, focal adhesion, estrogen signaling pathway, gastric cancer, HIF-1 signaling pathway, pathways in cancer, proteoglycans in cancer, lipid and atherosclerosis, epidermal growth factor receptor (EGFR) tyrosine kinase inhibitor resistance, endocrine resistance, advanced glycation endproducts–receptor for advanced glycation endproducts (AGE-RAGE) signaling pathway in diabetic complications, Kaposi sarcoma-associated herpesvirus infection, and human cytomegalovirus infection. Likewise, for two diseases, the GO enrichment analysis showed that the targets of L. decastes were always closely related to four biological processes, which were enzyme binding, response to xenobiotic stimulus, macromolecular complex and positive regulation of gene expression. The KEGG pathway enrichment analysis indicated that the targets of L. decastes were always significantly enriched in nine pathways, which were pathways in cancer, proteoglycans in cancer, lipid and atherosclerosis, EGFR tyrosine kinase inhibitor resistance, endocrine resistance, AGE-RAGE signaling pathway in diabetic complications, Kaposi sarcoma-associated herpesvirus infection, human cytomegalovirus infection, and prolactin signaling pathway.
Components–targets–pathways–diseases network analysis
The “components–targets–pathways–diseases” network analysis diagram illustrates the interplay between various elements (Figure 4). Yellow rectangles denote biological processes or signaling pathways, blue rectangles represent key targets associated with these pathways, red diamonds depict potential active components, and green ovals indicate diseases. This figure serves to elucidate the intricate relations among these components and their relevance to disease pathogenesis. It can be seen that the potential active components, such as octadecadienoic acid, octadecatrienoic acid, and lapachol in L. edodes, can act on 22 targets, such as PARP1, CTNNB1, and steroid receptor coactivator (SRC), to regulate pathways in cancer. EGFR tyrosine kinase inhibitor and other signaling pathways produce immune and tumor suppressive effects (Figure 5A). Similarly, potential active ingredients, such as octadecadienoic acid, octadecenoic acid, and linoleic acid in L. decastes, can act on 13 targets, such as GSK3B, AKT1, and PTGS2 points, and then regulate signaling pathways, such as pathways in cancer, EGFR tyrosine kinase inhibitor, AGE-RAGE signaling pathway, etc., to produce immune and tumor suppressive effects (Figure 5B).
Figure 4. The GO and KEGG enrichment analysis of core pharmacodynamic targets. (D) L. decastes components and CD; and (E) Venn diagram of the results of enrichment analysis of the intersection of components and diseases. The results of GO analysis are on the left and that of KEGG analysis on the right.
Figure 5. Components–targets–pathways–diseases network analysis. (A) L. edodes components and two diseases and (B) L. decastes components and two diseases.
Cell experiment verification
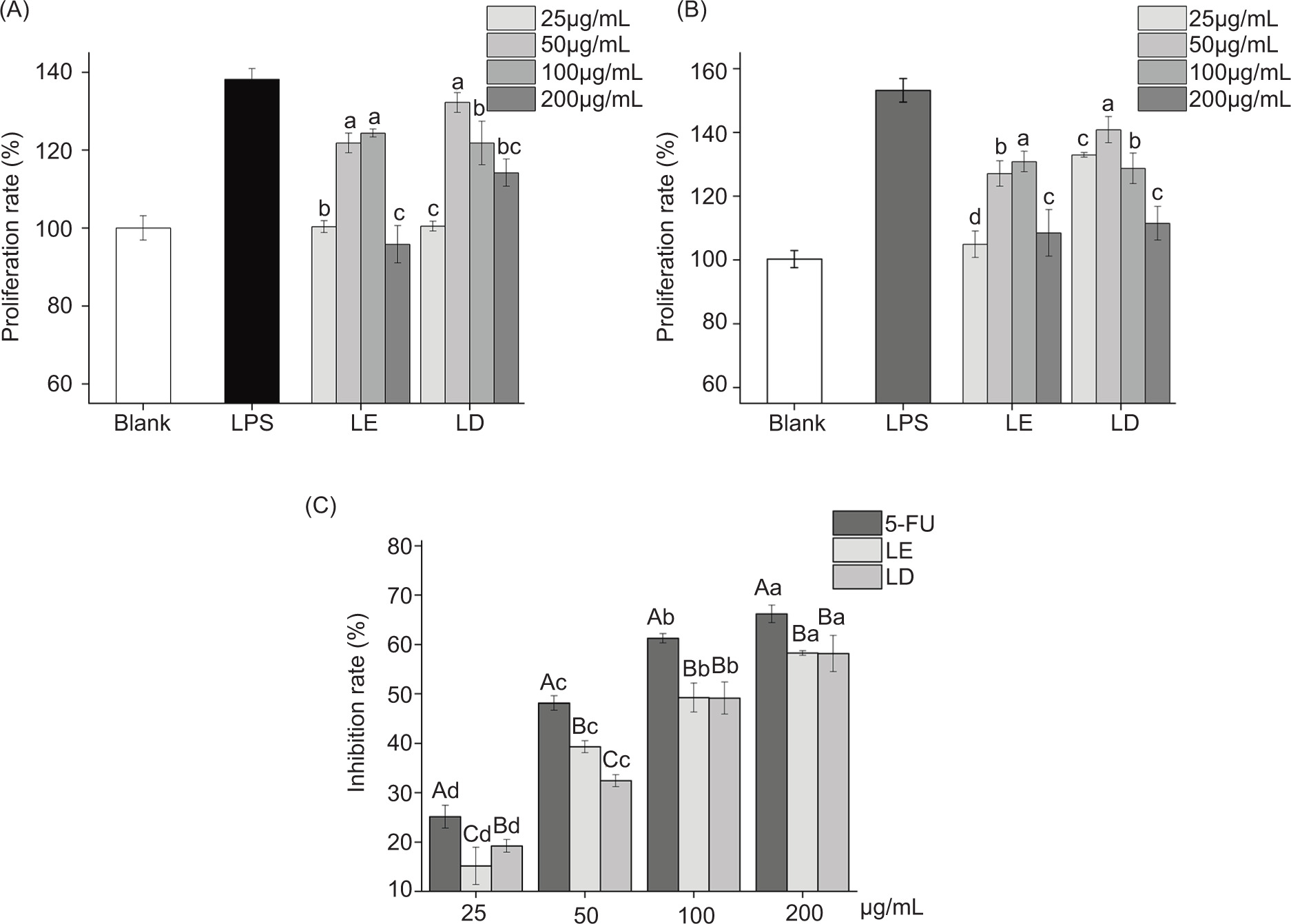
The cell proliferation rate or phagocytosis rate of blank control group was defined as 100%. Changes in the proliferation and phagocytosis of macrophages of RAW264.7 were studied. The results showed significant differences in cell proliferation and phagocytosis between the experimental group and the control group. In addition, dosage differences were observed between different concentrations. At appropriate concentrations, edible fungi have a significant stimulating effect on macrophages, significantly promoting their growth and activity (Figures 6A and 6B). Similarly, as concentration increased, the toxicity of edible fungi to HepG-2 became stronger, with a clear effect of inhibiting cell growth (Figure 6C).
Figure 6. Changes in the activity of different cells after edible fungi treatment. (A) Changes in RAW264.7 proliferation rate; (B) changes in RAW264.7 phagocytosis rate and (C) changes in HepG-2 inhibition rate. Lowercase letters represent significant differences between different concentrations in the same group, and uppercase letters represent significant differences between different groups at the same concentration (n = 6, P < 0.05).
Discussion
Modern studies on the composition and pharmacological activity of L. edodes and L. decastes mostly focused on single component and single activity, which seriously interfered the development and utilization of L. edodes and L. decastes. Traditional Chinese medicine emphasizes the holistic concept, and its drug use has the characteristics of multi-target and multi-function. Furthermore, various pieces of evidence suggest that targets, biological processes, and metabolic pathways play important roles in diseases. In our study, we used the PPI network to construct network targets for fungi and diseases from a holistic and systemic perspective for further screening core genes. Subsequently, we explored their potential mechanisms of action through GO and KEGG enrichment analysis. Finally, we utilized the component–target–pathway-disease network to unveil the mechanism through which its components influenced targets, regulated target pathways or biological processes, and consequently treated diseases.
First, we discussed several targets of interest. AKT1, also known as protein kinase B, is an important member of mitogen-activated protein kinase that exerts a key role in cell signaling. It serves as one of the central components of the PI3K/AKT signaling pathway and is involved in regulating a variety of biological processes, such as cell growth, proliferation, survival, and metabolism (Guerau-Arellano et al., 2022). Within the immune system, AKT1 is involved in cell proliferation, differentiation, and survival of T and B cells, and its activation may promote immune cell functions. AKT1 modulated adaptive immune response of Nile tilapia by promoting lymphocyte activation and proliferation via mammalian target of rapamycin complex 1 (mTORC1) signaling (Ai et al., 2021). In cancer, aberrant activation of AKT1 occurs, and its activation may promote tumor cell proliferation, survival, and invasion and inhibits apoptosis, rendering it a potential therapeutic target. For example, Yang et al. (2022) demonstrated that inhibition of AKT1 suppresses the growth of prostate cancer cells.
PPARG is a nuclear receptor that acts as a transcription factor and is widely distributed in a variety of tissues, such as adipose tissue, liver, intestine, and immune cells, in the human body (Rudko et al., 2020). Within the immune system, PPARG is involved in regulating inflammatory response, cell migration, and phagocytosis in the monocyte–macrophage system. Studies demonstrated that PPARG could be employed to temper allergic inflammation by suppressing pro-inflammatory gene expression programs in epithelial cells (Stark et al., 2021). In addition, many synthetic agonists of PPARG are shown to suppress hepatocellular carcinoma (HCC); these synthetic agonists prevent HCC invasion and metastasis by inducing cell cycle arrest and apoptosis in HCC cells (Katoch et al., 2022).
In addition to several shared pathways, there are also specific targets directed by components for treating diseases. For instance, protein tyrosine phosphatase receptor type C (PTPRC) is identified as the target modulated by L. edodes in the treatment of ISD. The PTPRC gene, also known as the CD45 gene, encodes a phosphatase located on the cell membrane. Immune-related genes, such as CD45 and CD69, exhibit activation in hyperimmune subtype group of chronic kidney disease (CKD), leading to a higher proportion of immune cells. This finding holds promising implications for the treatment of CKD (Fang et al., 2024). Similarly, the protein kinase C alpha (PRKCA) target gene is affected by L. decastes in the treatment of CKD. PRKCA encodes the protein kinase C alpha. The circular(circ)-PRKCA promotes the tumorigenesis of non-small cell lung cancer (NSCLC). Xu et al. (2021) demonstrated that curcumin inhibits the growth of NSCLC by downregulating circ-PRKCA.
Second, we discussed various important pathways of interest. EGFR tyrosine kinase inhibitor resistance is the most significant pathway of fold enrichment. EGFR, the receptor for members of the epidermal growth factor family, regulates cell proliferation and signal transduction. Moreover, EGFR is associated with inhibiting cell proliferation, angiogenesis, invasion, metastasis, and apoptosis. Therefore, EGFR has emerged as an important target for the treatment of cancer, including NSCLC, head and neck cancers, breast cancer, glioma, cervical cancer, and bladder cancer (Shi et al., 2022). The EGFR signaling pathway regulates proliferation, migration, and cytokine secretion of immune cells, thereby impacting the activity and functioning of immune cells. The inhibition of EGFR mediates the activation of immune cells and enhances the local proliferation of T cells in the environment (Zou et al., 2020). EGFR inhibitors block cell proliferation, metastasis, and other signaling pathways by inhibiting EGFR tyrosine kinase activity, thereby inhibiting the growth of cancer cells. Upon activation, EGFR triggers multiple downstream signaling pathways, such as PI3K/AKT and mitogen-activated protein kinase (MAPK) pathways, to promote cell growth. Proliferation, survival, and metastasis thereby promote the increased activity of immune cells (Selenz et al., 2022).
The MAPK pathway is an important signaling pathway. It plays a key regulatory role in cells and is involved in the regulation of various biological processes, such as cell proliferation, apoptosis, cell differentiation, cell migration, and inflammation. MAPKs are one of the most important enzymes in various cellular activities and involve a series of phosphorylation reactions. They are activated through receptors on cell membranes, respond to external signals, and regulate cell growth, differentiation, apoptosis, stress, and other processes (Kim et al., 2018). The MAPK family involves most of the pathways and mediates the activation process of immune cells, such as macrophages, T cells, and B cells. When cells are stimulated by external stimuli, such as bacterial and viral infection or cytokine stimulation, the MAPK pathway is activated, thereby triggering the corresponding response of the cell (Vilela et al., 2010). Both MAPK1 and MAPK3 belong to the mitogen-activated protein kinase family and promote the proliferation as well as inhibition of the apoptosis of cancer cells. Lee et al. (2022) used the inRas37 antibody, a KRAS-targeting antibody, to increase significantly the drug response of dual inhibitor BEZ-235 (also known as NVP-BEZ235 or RTB101) against pancreatic cancer cells by inhibiting MAPK.
In addition to several shared pathways, specific pathways targeted by components for treating diseases are also studied. The biological process involving most of the genes in the treatment of CD by L. edodes is identical protein binding, which refers to the process in which two or more identical proteins bind to each other to form a complex, and the binding of certain identical proteins may alter the transcription, translation, or stability of oncogenes, thus affecting the functioning of relevant genes. Wei et al. (2022) found that cancer cells often show resistance to drugs, and that the binding of the same proteins regulates relevant signaling pathways, affecting the sensitivity of cancer cells to therapeutic drugs, and thus affecting the treatment of cancer. The AGE-RAGE signaling pathway is the signaling pathway with the smallest P-value and the largest enrichment index in the treatment of ISD by L. decastes. The activation of AGE-RAGE signaling pathway induces the production of cytokines, such as TNF-α and IL-6, which are able to activate immune cells. The pathway also plays a key role in the maintenance of immune tolerance and suppression of immune response by influencing the functioning and number of immune cells, including regulatory T-cells (Sukjamnong et al., 2022).
Finally, we discussed the “components–targets–pathways–diseases” network. It was indicated that L. edodes and L. decastes had potential effect on regulating overlapping targets and multiple pathways, thus serving as a latent multi-target and multi-pathway treatment for two mentioned diseases. It is observed that seven active components of L. edodes and nine active components of L. decastes act on common targets, such as AKT1 and PPARG, thereby regulating signal pathways, such as EGFR and MAPK, to affect the body, and have immunomodulatory and liver tumor inhibition effects. The results showed that the mechanism of action of two fungi includes maintaining autoimmune tolerance and preventing normal tissues from being attacked by the immune system. External signal molecules can bind to receptors or signal molecules on cell membrane, triggering the activation and functional regulation of immune cells (Zhang et al., 2021). SRC regulates the activity of immune cells by affecting signal transduction, such as regulating the phosphorylation state of cytoskeleton (Chen et al., 2023) as well as inhibiting signaling pathways to enhance the immune response ability of immune cells and enhancing the immune system’s ability to attack tumor cells. EGFR signaling pathways inhibition mediates immune cell activation and increases local proliferation of T cells in the tumor environment (Halder et al., 2023). Therefore, the combined application of tumor suppressor drugs and immunotherapy drugs has a synergistic effect on cancer treatment. Shin et al. (2016) used an EGFR tyrosine kinase inhibitor and a PD-1 inhibitor, in combined application, blocking the combination of PD-1 and PD-L1 to inhibit the signaling pathway and enhance the killing ability of immune cells against tumor cells.
Conclusions
This study identified key bioactive components in L. edodes and L. decastes with potential immuno-enhancing and anticancer properties. It also explored the mechanisms of how active components exert their effects. The therapeutic mechanisms of L. edodes and L. decastes for immune system diseases include maintaining autoimmune tolerance, preventing normal tissues from being attacked by the immune system, inhibiting signal pathways, and enhancing the immune response of immune cells. On the other hand, the therapeutic mechanisms for liver cancer include inhibiting the activation of mitogen-activated proteins, blocking the EGFR pathway, and inhibiting the growth and metastasis of cancer cells. In addition, edible fungi also play both immunomodulatory and liver tumor inhibition roles. It was observed that enhancing body’s immune response is an important way to treat cancer. These findings serve as a crucial reference for elucidating the biological activities of L. edodes and L. decastes, and predict mechanisms of drug action. Activity verification in cell experiments also confirmed our analysis. It is recommended to explore the clinical applications of these bioactive ingredients and their mechanisms of action in more detail.
Data Availability Statement
Data are available upon request.
Author Contributions
Yingnan Zhang: conceptualization, data curation, investigation, formal analysis, writing of original draft. Gaoxing Ma: writing, review and editing as well as supervision. Fei Pei: writing, review and editing, and software application. Ning Ma: writing, review and editing as well as visualization. Anxiang Su: writing, review and editing as well as validation. Qiuhui Hu: conceptualization, methodology, fund acquisition, and project administration. Meng Wang: conceptualization, methodology, formal analysis, writing of original draft, and review and editing.
Conflicts of Interest
The authors declared that they had no known competing financial interests or personal relationships that could have influenced this paper.
Funding
This research was funded by Jiangsu Province Agricultural Science and Technology Innovation Fund (CX(21)2005).
REFERENCES
Ai, K.T., Yan, J., Li, K., et al., 2021. Akt1/mTORC1 signaling modulates adaptive immune response of Nile tilapia by promoting lymphocyte activation and proliferation. Developmental and Comparative Immunology 119: 10. 10.1016/j.dci.2021.104042
Chen, K., Dai, M.X., Luo, Q.N., et al., 2023. PARP1 controls the transcription of CD24 by ADP-ribosylating the RNA helicase DDX5 in pancreatic cancer. International Journal of Biochemistry–Cell Biology 155: 11. 10.1016/j.biocel.2022.106358
Coudray, C., Durand, E., Balas, L., et al., 2021. Potential favourable health effects of some dietary uncommon fatty acids. Ocl-Oilseeds and Fats Crops and Lipids 28(41): 9. 10.1051/ocl/2021028
Cui, L., Liu, Y., Liu, M., et al., 2022. Identification of phytochemicals from Lentinus edodes and Auricularia auricula with UPLC-Q-Exactive Orbitrap MS. Journal of Future Foods 2(3): 253–260. 10.1016/j.jfutfo.2022.06.006
Dai, J.N., Liu, B.L., Ji, D., et al., 2023a. Extraction, isolation, identification, and bioactivity of polysaccharides from Antrodia cinnamomea. Quality Assurance and Safety of Crops–Foods 15(4): 60–76. 10.15586/qas.v15i4.1341
Dai, Y., Wang, L., Chen, X.Y., et al., 2023b. Lentinula edodessing polysaccharide: extraction, characterization, bioactivities, and emulsifying applications. Foods 12(17): 17. 10.3390/foods12173289
Ding, X., Liu, Y., and Hou, Y. L., 2022. Structure identification and biological activities of a new polysaccharide isolated from Lyophyllum decastes (Fr.) Sing. Pharmacognosy Magazine 18(77): 112–120. 10.4103/pm.pm_185_21
Du, Y.H., Zhao, J.J., Li, X., et al., 2024. Mechanism of pachymic acid in the treatment of gastric cancer based on network pharmacology and experimental verification. World Journal of Gastrointestinal Oncology 16(1): 22. 10.4251/wjgo.v16.i1.30
Fang, X.D., Chen, Y.X., Chen, Y., et al., 2024. Identification and characterization of two immune-related subtypes in human chronic kidney disease. Transplant Immunology 82: 10. 10.1016/j.trim.2023.101983
Fujita, R., Yokono, M., Ube, N., et al., 2021. Suppression of alternaria brassicicola infection by volatile compounds from spent mushroom substrates. Journal of Bioscience and Bioengineering 132(1): 25–32. 10.1016/j.jbiosc.2021.03.003
Gameli-Kwabla, K., 2020. Examining the nutritional composition, value and health benefit of mushrooms. International Journal of Reseach and Scholarly Communication 3(1): 17.
Garcia, J., Rodrigues, F., Saavedra, M.J., et al., 2022. Bioactive polysaccharides from medicinal mushrooms: a review on their isolation, structural characteristics and antitumor activity. Food Bioscience 49: 11. 10.1016/j.fbio.2022.101955
Guerau-Arellano, M., Piedra-Quintero, Z.L.L., and Tsichlis, P.N., 2022. Akt isoforms in the immune system. Frontiers in Immunology 13: 990874. 10.3389/fimmu.2022.990874
Halder, S., Basu, S., Lall, S.P., et al., 2023. Targeting the EGFR signaling pathway in cancer therapy: what’s new in 2023? Expert Opinion on Therapeutic Targets 27(4–5): 305–324. 10.1080/14728222.2023.2218613
Hye-Lim, J., Park, S.-Y., and Nam, J., 2017. Effect of extraction solvent on the antioxidant activity of Lentinula edodes GNA01 extract. Korean Journal of Food and Nutrition 30(1): 51–58. 10.9799/ksfan.2017.30.1.051
Ike, K., Kameyama, N., Ito, A., et al., 2012. Induction of a T-Helper 1 (Th1) immune response in mice by an extract from the pleurotus eryngii (Eringi) mushroom. Journal of Medicinal Food 15(12): 1124–1128. 10.1089/jmf.2012.2239
Katoch, S., Sharma, V., and Patial, V., 2022. Peroxisome proliferator-activated receptor gamma as a therapeutic target for hepatocellular carcinoma: experimental and clinical scenarios. World Journal of Gastroenterology 28(28): 21. 10.3748/wjg.v28.i28.3535
Kim, M., and Ryu, S.E., 2018. Specific binding and catalytic activation of the MAPK-MKP complex. Biodesign 6(4): 79–83.
Lee, J.E., Woo, M.G., Jung, K.H., et al., 2022. Combination therapy of the active KRAS-targeting antibody inRas37 and a PI3K inhibitor in pancreatic cancer. Biomolecules and Therapeutics 30(3): 274–283. 10.4062/biomolther.2021.145
Li, H.Y., Guo, L., Ding, X.Y., et al., 2022. Molecular networking, network pharmacology, and molecular docking approaches employed to investigate the changes in ephedrae herba before and after honey-processing. Molecules 27(13): 18. 10.3390/molecules27134057
Li, Y.H., Liu, X.H., Zhang, X., et al., 2021. Immune cycle-based strategies for cancer immunotherapy. Advanced Functional Materials 31(50): 27. 10.1002/adfm.202107540
Liang, Z., Li, C., Wang, J., et al., 2021. Immunomodulatory effects of polysaccharides from edible fungus: a review. Food Science and Human Wellness 10(4): 393–400. 10.1016/j.fshw.2021.04.001
Lin, M., 2012. Fungi medicine in Chinese materia medica. Chinese Traditional and Herbal Drugs.
Lu, X.S., Hou, H., Fang, D.L., et al., 2022. Identification and characterization of volatile compounds in Lentinula edodes during vacuum freeze-drying. Journal of Food Biochemistry 46(6): 13. 10.1111/jfbc.13814
Sharma, A., Bhardwaj, G., Nayik, G.A. 2023. Phytochemistry and nutritional composition of significant wild medicinal and edible mushrooms. eBook collection. Royal Society of Chemistry, London. 10.1039/9781837672097
Qiu, M.Y., Zhang, J.Q., Wei, W.L., et al., 2024. Integrated UPLC/Q-TOF-MS/MS analysis and network pharmacology to reveal the neuroprotective mechanisms and potential pharmacological ingredients of aurantii fructus immaturus and aurantii fructus. Pharmaceuticals 17(2): 14. 10.3390/ph17020239
Roszczyk, A., Turlo, J., Zagozdzon, R., et al., 2022. Immunomodulatory properties of polysaccharides from Lentinula edodes. International Journal of Molecular Sciences 23(16): 13. 10.3390/ijms23168980
Rudko, O.I., Tretiakov, A.V., Naumova, E.A., et al., 2020. Role of PPARs in progression of anxiety: literature analysis and signaling pathways reconstruction. PPAR Research 2020: 8859017. 10.1155/2020/8859017
Selenz, C., Compes, A., Nill, M., et al., 2022. EGFR inhibition strongly modulates the tumour immune microenvironment in EGFR-Driven non-Small-Cell lung cancer. Cancers 14(16): 15. 10.3390/cancers14163943
Sganzerla, W.G., Todorov, S.D., and da Silva, A.P.G., 2022. Research trends in the study of edible mushroom: nutritional properties and health benefits. International Journal of Medicinal Mushrooms 24(5): 1–18.
Shi, K.Y., Wang, G., Pei, J.P., et al., 2022. Emerging strategies to overcome resistance to third-generation EGFR inhibitors. Journal of Hematology and Oncology 15(1): 44. 10.1186/s13045-022-01311-6
Shin, J.H., and Park, H.-B., 2016. Enhanced anti-tumor reactivity of cytotoxic T lymphocytes expressing PD-1 decoy. Immune Network 16(2): 134–139. 10.4110/in.2016.16.2.134
Stark, J.M., Coquet, J.M., and Tibbitt, C.A., 2021. The role of PPAR-γ in allergic disease. Current Allergy and Asthma Reports 21(11): 8. 10.1007/s11882-021-01022-x
Starzer, A.M., Preusser, M., and Berghoff, A.S., 2022. Immune escape mechanisms and therapeutic approaches in cancer: the cancer-immunity cycle. Therapeutic Advances in Medical Oncology 14: 14. 10.1177/17588359221096219
Sukjamnong, S., Chen, H., Saad, S., et al., 2022. Fimbristylisovata and artemisia vulgaris extracts inhibited AGE-mediated RAGE expression, ROS generation, and inflammation in THP-1 cells. Toxicological Research 38(3): 331–343. 10.1007/s43188-021-00114-0
Vilela, B., Pagès, M., and Lumbreras, V., 2010. Regulation of MAPK signaling and cell death by MAPK phosphatase MKP2. Plant Signaling and Behavior 5(11): 1497–1500. 10.4161/psb.5.11.13645
Wang, Y.H., Yang, Z.Y., Chen, X., et al., 2021. Lenthionine, a key flavor substance in Lentinula edodes, is regulated by cysteine under drought stress. Journal of Agricultural and Food Chemistry, 69(43): 12645–12653. 10.1021/acs.jafc.1c04829
Wei, A.A.J., Iacobucci, C., Schultze, W., et al., 2022. Different oligomeric states of the tumor suppressor p53 show identical binding behavior towards the S100β homodimer. Chembiochem 23(11): 6. 10.1002/cbic.202100665
Wen, S.D., Mo, S., Zhou, J., et al., 2024. Single-cell and spatial-omics in delineating immune-related diseases. Frontiers in Cell and Developmental Biology 12: 3. 10.3389/fcell.2024.1365242
Xie, Y.M., Shi, L.S., He, X.S., et al., 2021. Gastrointestinal cancers in China, the USA, and Europe. Gastroenterology Report 9(2): 91–104. 10.1093/gastro/goab010
Xu, L.L., Yang, W.J., Qiu, T.M., et al., 2023. Complete genome sequences and comparative secretomic analysis for the industrially cultivated edible mushroom Lyophyllum decastes reveals insights on evolution and lignocellulose degradation potential. Frontiers in Microbiology 14: 14. 10.3389/fmicb.2023.1137162
Xu, X.M., Yu, D.H., Wang, Y., et al., 2024. Integrating network pharmacology and renal metabonomics to reveal the protective mechanism of resveratrol on gouty nephropathy. Biomedical Chromatography 38(5): e5839. 10.1002/bmc.5839
Xu, X.Q., Zhang, X.Y., Zhang, Y., et al., 2021. Curcumin suppresses the malignancy of non-small cell lung cancer by modulating the circ-PRKCA/miR-384/ITGB1 pathway. Biomedicine & Pharmacotherapy 138: 111439. 10.1016/j.biopha.2021.111439
Yang, M., Liu, H., Qiu, G.P., et al., 2022. Silencing Akt1 enhances the resistance of prostate cancer cells to starvation and inhibits starvation-induced lung metastasis through epithelial-mesenchymal transition in prostate cancer. Medical Oncology 39(1): 12. 10.1007/s12032-021-01600-z
Yin, Z., Sun-Waterhouse, D., Wang, J., et al., 2022. Polysaccharides from edible fungi Pleurotus spp.: advances and perspectives. Journal of Future Foods 1(2): 128–140. 10.1016/j.jfutfo.2022.01.002
Zhang, P., He, S., Wu, S., et al., 2022. Discovering a multi-component combination against vascular dementia from danshen-honghua herbal pair by spectrum-effect relationship analysis. Pharmaceuticals (Basel) 15(9): 1073. https://www.mdpi.com/1424-8247/15/9/1073
Zhang, G.P., Wang, Y.N., Qin, C.Q., et al., 2023. Structural characterization of an antioxidant polysaccharide isolated from the fruiting bodies of Lyophyllum decastes. Journal of Molecular Structure 1285: 8. 10.1016/j.molstruc.2023.135507
Zhang, T.T., Hu, W., Chen, W., et al., 2021. Plasma membrane integrates biophysical and biochemical regulation to trigger immune receptor functions. Frontiers in Immunology 12: 8. 10.3389/fimmu.2021.613185
Zhou, Y., Chu, M.H., Ahmadi, F., et al., 2023. A comprehensive review on phytochemical profiling in mushrooms: occurrence, biological activities, applications and future prospective. Food Reviews International 40(3): 924–951. 10.1080/87559129.2023.2202738
Zhu, B.J., Nai, G.Y., Pan, T.X., et al., 2023. Combining network pharmacology and bioinformatics to identify bioactive compounds and potential mechanisms of action of Sedum aizoon L in the treatment of atherosclerosis. Quality Assurance and Safety of Crops & Foods 15(3): 104–116. 10.15586/qas.v15i3.1333
Zou, H., Zhou, C., Li, Y., et al., 2020. Speciation analysis of arsenic in edible mushrooms by high-performance liquid chromatography hyphenated to inductively coupled plasma mass spectrometry. Food Chemistry 327: 127033. 10.1016/j.foodchem.2020.127033
Supplementary
Table S1. Detailed information on components identification of L. edodes.
| No. | Components | Formula | Adducts | Precursor ions (m/z) |
Peak area | Retention time (min) |
|---|---|---|---|---|---|---|
| 1. | Oxidized fatty acids | C18H34O4 | [M-H]- | 313.2379 | 389364 | 13.24663 |
| 2. | Organic acids | C6H8O7 | [M-H]- | 191.0194 | 354182.7 | 1.903083 |
| 3. | Linoleic acids and derivatives | C18H30O4 | [M-H]- | 309.2069 | 246438 | 12.98182 |
| 4. | Secondary alcohols | C9H17NO5 | [M-H]- | 218.1031 | 197291.8 | 4.29345 |
| 5. | Oxidized fatty acids | C18H30O4 | [M-H]- | 309.2074 | 79351.45 | 12.35152 |
| 6. | 6-Alkyl amino purines | C10H13N5 | [M-H]- | 202.1093 | 45440.13 | 2.145233 |
| 7. | 3’-O-methylated flavonoids | C16H14O6 | [M-H]- | 300.9998 | 39894.22 | 7.134133 |
| 8. | Diterpene glycosides | C52H82O21 | [M-H]- | 1041.528 | 29063.5 | 8.181566 |
| 9. | Medium-chain fatty acids | C9H16O4 | [M-H]- | 187.097 | 27965.81 | 8.1414 |
| 10. | Diarylethers | C37H41ClN2O6 | [M-H]- | 643.0568 | 22684.85 | 1.661933 |
| 11. | Phenylalanine and derivatives | C12H15NO4 | [M-H]- | 236.0926 | 14982.2 | 7.6181 |
| 12. | Medium-chain hydroxy acids and derivatives | C6H12O7 | [M-H]- | 195.0514 | 259383.6 | 1.541767 |
| 13. | Nucleoside and nucleotide analogues | C9H12N2O6 | [M-H]- | 243.0627 | 222588.6 | 1.862583 |
| 14. | Rotenones | C23H22O6 | [M-H]- | 393.134 | 59459.38 | 5.595883 |
| 15. | Purine nucleosides | C11H14N4O5 | [M-H]- | 281.0896 | 45041.5 | 3.083017 |
| 16. | Sugar alcohols | C5H12O5 | [M-H]- | 151.0615 | 27360.21 | 1.261617 |
| 17. | O-glucuronides | C8H14O7 | [M-H]- | 221.0935 | 22846.38 | 5.028917 |
| 18. | Oligosaccharides | C18H32O16 | [M+H]+ | 543.1328 | 37676.8 | 1.29325 |
| 19. | Dipeptides | C11H20N2O5 | [M-H]- | 259.1294 | 36801.76 | 6.284517 |
| 20. | Gallic acid and derivatives | C14H21N3O5 | [M+H]+ | 334.1407 | 32429.91 | 9.160983 |
| 21. | Fatty acyl glycosides of mono- and disaccharides | C11H17NO6 | [M-H]- | 258.0976 | 20816.99 | 6.406 |
| 22. | Dicarboxylic acid and derivatives | C20H34O7 | [M+H]+ | 409.2202 | 15026.07 | 10.57958 |
| 23. | Beta hydroxy acid (BHAs) and derivatives | C5H8O5 | [M-H]- | 147.0291 | 66964.04 | 1.903083 |
| 24. | Tricarboxylic acids and derivatives | C6H6O6 | [M+H]+ | 192.0512 | 42917.2 | 4.683067 |
| 25. | Purine nucleosides | C10H13N5O4 | [M-H]- | 266.0905 | 35266.7 | 2.145233 |
| 26. | Long-chain fatty acids | C16H32O3 | [M-H]- | 271.227 | 20259.31 | 18.42863 |
| 27. | Peptides | C11H18N2O4 | [M-H]- | 241.1199 | 18800.83 | 6.60765 |
| 28. | Nicotinamides | C13H12N2O2 | [M-H]- | 227.0798 | 15118.66 | 1.261617 |
| 29. | N-acetylneuraminic acid | C11H19NO9 | [M-H]- | 308.0997 | 45247.01 | 1.261617 |
| 30. | Flavonoid O-glycosides | C21H20O11 | [M-H]- | 447.2018 | 23633.68 | 12.31135 |
| 31. | Long-chain fatty acids | C18H34O5 | [M-H]- | 329.2351 | 198892.7 | 11.36573 |
| 32. | N-fructosyl amino acids | C11H17NO8 | [M-H]- | 290.0861 | 31198.29 | 1.862583 |
| 33. | Cinnamic acid and derivatives | C9H10O3 | [M-H]- | 165.0568 | 25600.38 | 7.457117 |
| 34. | O-glycosyl compounds | C22H36O12 | [M-H]- | 491.2145 | 23579.73 | 7.0938 |
| 35. | Naphthopyranones | C28H22O10 | [M+H]+ | 557.149 | 59677.49 | 1.372917 |
| 36. | Phenolic glycosides | C18H18O9 | [M-H]- | 377.0864 | 50035.88 | 1.261617 |
| 37. | Prenylated isoflavones | C21H20O5 | [M-H]- | 351.1205 | 22181.2 | 6.001017 |
| 38. | Indoline | C8H9N | [M+H]+ | 120.0822 | 54571.89 | 3.703167 |
| 39. | Amino acids | C10H15NO4 | [M-H]- | 212.0914 | 14093.35 | 8.543384 |
| 40. | 3-O-methylated flavonoids | C16H12O7 | [M+H]+ | 334.0868 | 18400.59 | 1.5729 |
| 41. | Caffeic acid and derivatives | C15H18O9 | [M-H]- | 341.0855 | 125466.6 | 4.4657 |
| 42. | Terpene glycosides | C23H34O16 | [M-H]- | 565.1792 | 36561.27 | 1.983583 |
| 43. | Xanthines | C6H6N4O2 | [M-H]- | 165.0401 | 30156.22 | 1.221283 |
| 44. | Dicarboxylic acids and derivatives | C4H6O4 | [M+H]+ | 119.0356 | 32159.07 | 4.683067 |
| 45. | Dipeptides | C8H16N2O3 | [M-H]- | 187.1105 | 57869.88 | 3.8903 |
| 46. | N-fructosyl amino acids | C12H23NO7 | [M-H]- | 292.141 | 154678.1 | 2.1049 |
| 47. | O-glycosyl compounds | C12H22O11 | [M-H]- | 341.1109 | 94117.92 | 1.261617 |
| 48. | Purine nucleosides | C12H17N5O5 | [M-H]- | 310.1134 | 40945.61 | 1.421617 |
| 49. | Flavonoid-7-O-glycosides | C20H22O10 | [M-H]- | 421.1041 | 21765.38 | 1.261617 |
| 50. | Perfluorosulfonic acid (PFSA) | C3HClF6O4S | [M-H]- | 280.9092 | 35606.07 | 4.745433 |
| 51. | 21-Hydroxysteroids | C21H30O5 | [M-H]- | 361.1995 | 22452.5 | 15.78948 |
| 52. | Indole-3-acetic acid and derivatives | C10H9NO2 | [M-H]- | 174.0589 | 23899.34 | 8.743867 |
| 53. | Phenolic glycosides | C24H34O9 | [M-H]- | 465.21 | 19084.85 | 10.28078 |
| 54. | Phenylquinolines | C23H24N2O | [M-H]- | 343.1847 | 30786.72 | 8.10125 |
| 55. | Glutamic acid and derivatives | C21H22N4O6S | [M-H]- | 457.1156 | 33433.43 | 1.4616 |
| 56. | Phenolic glycosides | C16H24O7 | [M-H]- | 327.1415 | 27273.35 | 1.221283 |
| 57. | Pyrimidine nucleosides | C9H13N3O5 | [M-H]- | 242.0817 | 16997.86 | 10.44595 |
| 58. | 7-O-methylisoflavones | C21H22O8 | [M-H]- | 401.2393 | 43095.44 | 14.29997 |
| 59. | Saccharolipids | C32H56O14 | [M+H]+ | 687.3612 | 33179.88 | 7.409417 |
| 60. | Coumarin and derivatives | C12H10O4 | [M-H]- | 217.0469 | 28455.31 | 1.221283 |
| 61. | Phenolic glycosides | C14H17NO8 | [M-H]- | 326.1235 | 549333.8 | 3.566817 |
| 62. | Germacranolides and derivatives | C22H30O8 | [M-H]- | 421.183 | 63248.78 | 5.474383 |
| 63. | Saccharolipids | C21H34O10 | [M+H]+ | 447.2263 | 39286.58 | 9.160983 |
| 64. | Dipeptides | C9H18N2O3 | [M-H]- | 201.123 | 51130.34 | 3.849817 |
| 65. | 7-O-methylated flavonoids | C19H18O7 | [M-H]- | 357.1019 | 29381.62 | 1.181117 |
| 66. | Hydroxypyrimidines | C5H7N3O2 | [M+H]+ | 142.065 | 22333.67 | 5.2352 |
| 67. | Phenolic glycosides | C13H18O8 | [M+H]+ | 341.0672 | 12574.11 | 10.38377 |
| 68. | Medium-chain fatty acids | C10H18O4 | [M-H]- | 201.1131 | 62232.11 | 8.9907 |
| 69. | Indoline | C16H10N2O2 | [M-H]- | 261.0627 | 17371.99 | 1.261617 |
| 70. | Macrolides and analogues | C16H20O6 | [M-H]- | 307.1142 | 36616.81 | 1.221283 |
| 71. | Chromeno[2,3-b]pyridine-5-ones | C16H14N2O4 | [M-H]- | 297.0834 | 48058.59 | 1.5016 |
| 72. | Hydroxy fatty acids | C7H12O5 | [M-H]- | 175.0606 | 45316.15 | 5.839366 |
| 73. | Glutamic acid and derivatives | C5H9NO4 | [M-H]- | 146.0458 | 95387.94 | 1.221283 |
| 74. | Purine nucleosides | C10H13N5O5 | [M-H]- | 282.085 | 436324.4 | 2.024083 |
| 75. | Oxolanes | C11H20O4 | [M-H]- | 215.0833 | 83713.98 | 6.001017 |
| 76. | Leucine and derivatives | C6H13NO2 | [M-H]- | 130.0871 | 79256.39 | 2.064567 |
| 77. | Sugar alcohols | C6H14O6 | [M-H]- | 181.0717 | 592400.9 | 1.261617 |
| 78. | Indole | C9H9N | [M+H]+ | 132.0803 | 42559.17 | 3.743117 |
| 79. | Naphthofurans | C33H42N2O6 | [M+H]+ | 1125.622 | 99433.64 | 6.97645 |
| 80. | Alpha amino acids and derivatives | C5H7NO3 | [M-H]- | 128.0369 | 137024.4 | 1.421617 |
| 81. | Flavonoid-7-O-glycosides | C33H40O15 | [M-H]- | 675.237 | 42808.21 | 3.566817 |
| 82. | Steviol glycosides | C32H50O13 | [M+H]+ | 643.3314 | 30073.94 | 7.205767 |
| 83. | Linoleic acid and derivatives | C18H32O2 | [M-H]- | 279.2319 | 95280.72 | 19.10748 |
| 84. | Oxidized fatty acids | C18H34O5 | [M-H]- | 329.2325 | 648702.8 | 10.28078 |
| 85. | Linoleic acid and derivatives | C18H30O2 | [M-H]- | 277.216 | 11723.33 | 18.10755 |
| 86. | 3-Alkylindoles | C9H9NO | [M+H-H2O]+ | 130.0662 | 40869.95 | 3.703167 |
| 87. | Oxidized fatty acids | C18H32O4 | [M-H]- | 311.2222 | 774960.1 | 12.31135 |
| 88. | Alkaloids | C23H30N2O4 | [M-H]- | 397.219 | 72047.72 | 10.28078 |
| 89. | Coumarins and derivatives | C15H16O4 | [M-H]- | 259.0548 | 17378.57 | 10.36578 |
| 90. | Purine nucleosides | C10H12N4O5 | [M-H]- | 267.073 | 87090.22 | 2.024083 |
| 91. | Pyrenes | C16H10O | [M-H]- | 217.0721 | 73213.62 | 5.474383 |
| 92. | Amino acids | C9H11NO2 | [M-H]- | 164.0713 | 297771.6 | 3.405 |
| 93. | Steroidal saponins | C38H60O12 | [M+Na]+ | 731.3843 | 30056.7 | 7.57175 |
| 94. | Purine nucleosides | C10H12N4O6 | [M-H]- | 283.0699 | 97514.69 | 2.508383 |
| 95. | Alpha amino acids and derivatives | C4H6N2O2 | [M+H]+ | 115.055 | 150391.1 | 5.2352 |
| 96. | BHAs and derivatives | C4H6O5 | [M+H]+ | 135.0319 | 97262.93 | 2.057717 |
| 97. | Alpha amino acids and derivatives | C3H6N2O2 | [M+H]+ | 103.0556 | 131141.7 | 3.4952 |
| 98. | Quinolones and derivatives | C9H7NO2 | [M+H]+ | 162.0558 | 50670.63 | 1.733733 |
| 99. | Cardenolide glycosides and derivatives | C41H64O14 | [M+H]+ | 803.4265 | 21263.3 | 7.9019 |
| 100. | Organic acids | C4H6O4 | [M-H]- | 117.0195 | 24347.64 | 1.5016 |
| 101. | Medium-chain hydroxy acids and derivatives | C10H20O3 | [M-H]- | 187.132 | 53034.64 | 9.269183 |
| 102. | Delta valerolactones | C24H36O5 | [M+H]+ | 427.2336 | 37145.64 | 7.942234 |
| 103. | Medium-chain hydroxy acids and derivatives | C6H12O7 | [M-H]- | 195.0509 | 332932.5 | 1.221283 |
| 104. | Indole and derivatives | C10H11NO | [M+H]+ | 144.0814 | 41152.14 | 6.0575 |
| 105. | Tricarboxylic acids and derivatives | C6H8O7 | [M-H]- | 191.0196 | 183291.3 | 1.381283 |
| 106. | Coumaric acid and derivatives | C31H38O17 | [M+H]+ | 705.1863 | 24483.38 | 1.29325 |
| 107. | Cardenolide glycosides and derivatives | C49H76O19 | [M+H]+ | 991.5059 | 29161.29 | 8.063884 |
| 108. | Purine nucleosides | C11H15N5O5 | [M-H]- | 296.0985 | 75328.03 | 2.957017 |
| 109. | Indolyl carboxylic acids and derivatives | C11H12N2O2 | [M-H]- | 203.0825 | 189869.1 | 5.191233 |
| 110. | 6,7-dihydroxycoumarins | C10H8O4 | [M+H]+ | 193.0501 | 13898.28 | 10.38377 |
| 111. | L-alpha-amino acids | C5H9NO4 | [M+H]+ | 148.0625 | 87936.63 | 1.693733 |
| 112. | BHAs and derivatives | C4H6O5 | [M-H]- | 133.0139 | 270234.9 | 1.381283 |
| 113. | 7-O-methylated isoflavonoids | C22H26O5 | [M+H]+ | 393.1607 | 28570.62 | 1.5729 |
| 114. | Vitamin K compounds | C15H14O3 | [M-H]- | 241.0834 | 73192.02 | 3.364833 |
| 115. | Precocenes | C15H18O4 | [M+H]+ | 263.1386 | 34388.84 | 9.160983 |
| 116. | Alkaloids and derivatives | C11H16N2O2 | [M+H]+ | 209.1306 | 15719.67 | 12.60347 |
| 117. | Sulfinic acid | C2H7NO2S | [M+H]+ | 110.0356 | 34988.04 | 2.057717 |
| 118. | Aryl alkyl ketones | C30H40NNaO4 | [M+H]+ | 502.2927 | 92229.73 | 12.60347 |
| 119. | Hydroxypyrimidines | C5H6N2O2 | [M+H]+ | 127.0566 | 18418.14 | 6.0575 |
| 120. | Carbazoles | C12H11NO | [M+H]+ | 208.0848 | 26025.08 | 1.693733 |
Table S2. Detailed information on the components identification of L. decastes.
| No. | Components | Formula | Adducts | Precursor ions (m/z) |
Peak area | Retention time (min) |
|---|---|---|---|---|---|---|
| 1. | Oxidized fatty acids | C18H34O5 | [M-H]- | 329.2328 | 688134.6 | 11.24757 |
| 2. | Oxidized fatty acids | C18H30O4 | [M-H]- | 309.206 | 549905.2 | 12.16518 |
| 3. | Secondary alcohols | C9H17NO5 | [M-H]- | 218.1035 | 332371.8 | 1.882917 |
| 4. | Oxidized fatty acids | C18H32O6 | [M-H]- | 343.2122 | 249427.7 | 10.88658 |
| 5. | Isobenzofurans | C12H20O6 | [M-H]- | 259.1188 | 99500.4 | 7.545767 |
| 6. | Tricarboxylic acids and derivatives | C6H8O7 | [M-H]- | 191.0197 | 87099.98 | 1.320283 |
| 7. | Hydrolyzable tannins | C14H6O8 | [M-H]- | 300.9989 | 74182.92 | 7.099133 |
| 8. | Lipids | C23H44NO7P | [M-H]- | 476.2775 | 67559.25 | 14.3249 |
| 9. | Dipeptides | C11H20N2O5 | [M-H]- | 259.1297 | 62994.46 | 1.400433 |
| 10. | Long-chain fatty acids | C16H32O3 | [M-H]- | 271.2279 | 59068.56 | 18.39802 |
| 11. | Phenoxyacetic acid derivatives | C20H32O4 | [M+H]+ | 359.2187 | 48512.67 | 13.0559 |
| 12. | Alkyl-phenylketones | C18H28ClNO2 | [M+H]+ | 290.2108 | 40847.5 | 8.422833 |
| 13. | Isoindolones | C25H35NO5 | [M+H]+ | 447.2852 | 34771.75 | 16.38438 |
| 14. | Gluco/mineralocorticoids, progestogins and derivatives |
C27H30Cl2O6 | [M+H]+ | 543.1307 | 33428.57 | 1.163717 |
| 15. | Medium-chain fatty acids | C9H16O4 | [M-H]- | 187.0977 | 30968.6 | 8.744534 |
| 16. | Leucothol and grayanotoxane diterpenoids | C22H36O7 | [M+H]+ | 430.2798 | 27825.4 | 8.995133 |
| 17. | Medium-chain fatty acids | C12H22O6 | [M-H]- | 261.1345 | 24333.88 | 7.058633 |
| 18. | N-fructosyl amino acids | C12H23NO7 | [M-H]- | 292.1393 | 21977.02 | 1.681083 |
| 19. | Dipeptides | C14H18N2O3 | [M-H]- | 261.1248 | 478412.1 | 5.6777 |
| 20. | Prenylated flavanones | C25H28O4 | [M-H]- | 391.1892 | 250268 | 10.5256 |
| 21. | Long-chain fatty acids | C18H34O3 | [M-H]- | 297.2432 | 37254.85 | 17.10558 |
| 22. | Medium-chain fatty acids | C12H20O4 | [M-H]- | 227.1294 | 27128.78 | 8.785033 |
| 23. | 3-Methylindoles | C9H9N | [M-H]- | 130.0666 | 21324.65 | 8.42155 |
| 24. | Indole-3-acetic acid derivatives | C10H9NO2 | [M-H]- | 174.0557 | 19317.33 | 8.42155 |
| 25. | Quassinoids | C20H30O8 | [M-H]- | 397.1863 | 11529.69 | 8.110733 |
| 26. | Nucleoside and nucleotide analogues | C9H12N2O6 | [M-H]- | 243.0629 | 189407.9 | 1.400433 |
| 27. | Oxidized fatty acids | C18H34O3 | [M-H]- | 297.2429 | 153862.4 | 17.38823 |
| 28. | Dipeptides | C13H16N2O5 | [M-H]- | 279.098 | 78626.05 | 2.861033 |
| 29. | Methoxyphenols | C11H14O4 | [M-H]- | 209.0812 | 71460.02 | 8.110733 |
| 30. | Alkyl-phenylketones | C15H17NO4 | [M-H]- | 274.1189 | 69269.67 | 5.191067 |
| 31. | Medium-chain fatty acids | C15H23NO4 | [M-H]- | 280.156 | 32681.96 | 10.5256 |
| 32. | Medium-chain fatty acids | C10H18O4 | [M-H]- | 201.1139 | 27231.96 | 8.987017 |
| 33. | O-glycosyl compounds | C12H22O11 | [M-H]- | 341.1097 | 190576.7 | 2.124567 |
| 34. | Medium-chain fatty acids | C7H12O4 | [M-H]- | 159.0655 | 53388.01 | 5.7182 |
| 35. | Cholines | C5H14NO | [M+H]+ | 104.1078 | 45776.67 | 8.543317 |
| 36. | Oxolanes | C11H20O4 | [M-H]- | 215.1297 | 26657.86 | 9.834483 |
| 37. | Phenolic glycosides | C12H16O6 | [M-H]- | 255.0882 | 26324.81 | 6.124183 |
| 38. | Glutamic acid and derivatives | C21H22N4O6S | [M-H]- | 457.1197 | 291020.1 | 1.360433 |
| 39. | Flavonoid 8-C-glycosides | C26H28O14 | [M-H]- | 563.2087 | 124534.1 | 2.251083 |
| 40. | Sesquiterpenoids | C30H30O8 | [M-H]- | 517.1877 | 38981.13 | 1.200617 |
| 41. | N-acyl-alpha amino acids | C8H16N2O3 | [M-H]- | 187.1071 | 38002.27 | 1.400433 |
| 42. | Pyridines and derivatives | C12H17N5 | [M-H]- | 230.142 | 26419.77 | 6.57165 |
| 43. | Triterpenoids | C30H40O4 | [M+H]+ | 465.296 | 19141.55 | 15.35178 |
| 44. | Purine nucleosides | C10H13N5O5 | [M-H]- | 282.0854 | 131522.8 | 1.6411 |
| 45. | Flavonoid O-glycosides | C21H20O11 | [M-H]- | 447.1997 | 37202.37 | 13.47867 |
| 46. | Butenolides | C9H14O4 | [M-H]- | 185.083 | 22890.21 | 9.552167 |
| 47. | Naphthopyrans | C21H28O6 | [M-H]- | 375.1825 | 140189.2 | 12.87817 |
| 48. | 21-Hydroxysteroids | C21H30O5 | [M-H]- | 361.2008 | 135548.9 | 15.36285 |
| 49. | Flavonoid O-glycosides | C21H22O10 | [M-H]- | 433.1739 | 35099.22 | 6.936967 |
| 50. | Imidazolyl carboxylic acid and derivatives | C10H18N2O3 | [M-H]- | 213.1228 | 28937.12 | 1.72125 |
| 51. | 1,4-Dioxanes | C14H24N2O7 | [M-H]- | 331.1523 | 27146.39 | 6.205184 |
| 52. | Oligopeptides | C19H27N3O6 | [M-H]- | 392.1841 | 89753.66 | 7.058633 |
| 53. | Medium-chain hydroxy acids and derivatives | C11H18O5 | [M-H]- | 229.1068 | 70034.16 | 8.58305 |
| 54. | Naphthalenes | C21H25N | [M+H]+ | 292.2046 | 19484.05 | 12.57427 |
| 55. | Indoles and derivatives | C16H23N5O | [M-H]- | 300.1814 | 25168.98 | 9.310184 |
| 56. | Fatty acid esters | C21H36O6 | [M-H]- | 383.2422 | 100392.2 | 14.08692 |
| 57. | Peptides | C12H24N2O3 | [M-H]- | 243.1733 | 45353.91 | 5.63705 |
| 58. | Acyloins | C21H30O5 | [M-H]- | 361.2041 | 118311.8 | 15.36285 |
| 59. | Oligopeptides | C28H44N4O4 | [M-H]- | 499.3311 | 22417.2 | 12.38345 |
| 60. | O-glucuronides | C8H14O7 | [M-H]- | 221.0919 | 290090.6 | 2.660033 |
| 61. | Isoflavones | C15H10O5 | [M-H]- | 269.0551 | 60320.98 | 8.542883 |
| 62. | Oxime ethers | C7H10N4O3 | [M-H]- | 235.1112 | 58757.01 | 2.619717 |
| 63. | Furopyrans | C26H32O8 | [M+H]+ | 473.2118 | 22371.86 | 5.646584 |
| 64. | Phenolic glycosides | C18H18O9 | [M-H]- | 377.0854 | 268756.6 | 1.200617 |
| 65. | Medium-chain fatty acids | C14H18O5 | [M-H]- | 265.1057 | 46270.35 | 8.030416 |
| 66. | 3’,5’-Cyclic purine nucleotides | C10H12N5O6P | [M-H]- | 328.0477 | 17821.92 | 1.6411 |
| 67. | 3-Alkylindoles | C13H16N2O2 | [M-H]- | 231.1245 | 32742.01 | 7.7481 |
| 68. | Phenolic glycosides | C14H17NO8 | [M-H]- | 326.1246 | 37233.94 | 1.76175 |
| 69. | Dicarboxylic acid and derivatives | C4H6O4 | [M+H]+ | 119.0369 | 153099.9 | 1.602367 |
| 70. | Cinnamic acid and derivatives | C18H21NO4 | [M-H]- | 314.1139 | 45394.23 | 7.058633 |
| 71. | Limonoids | C26H36O7 | [M+H]+ | 483.3055 | 30284.66 | 12.06363 |
| 72. | Eudesmanolides, secoeudesmanolides, and derivatives |
C25H38O12 | [M-H]- | 529.2255 | 58202.28 | 1.84275 |
| 73. | Phosphatidylcholines | C42H82NO8P | [M+H]+ | 760.5799 | 178083.9 | 15.63423 |
| 74. | Short-chain keto acids and derivatives | C5H8O3 | [M+H]+ | 117.0581 | 46462.05 | 1.88935 |
| 75. | Indolyl carboxylic acids and derivatives | C11H12N2O2 | [M-H]- | 203.0828 | 528820.9 | 3.034817 |
| 76. | Benzoic acid esters | C14H15N5O6S | [M-H]- | 380.0634 | 29223.68 | 1.6011 |
| 77. | Steroid esters | C31H48O6 | [M-H]- | 515.3584 | 21294.85 | 14.20557 |
| 78. | Naphthopyrans | C28H36O7 | [M-H]- | 483.2335 | 47921.86 | 9.390667 |
| 79. | Alkyl-phenylketones | C22H22FN3O2 | [M-H]- | 378.1661 | 112742.9 | 6.408834 |
| 80. | Phenolic glycosides | C15H18O8 | [M-H]- | 325.1616 | 83333.98 | 14.24523 |
| 81. | Isoindolones | C24H35NO4 | [M-H]- | 400.2318 | 43089.91 | 14.08692 |
| 82. | C20-Gibberellin 6-carboxylic acids | C20H26O5 | [M-H]- | 345.1666 | 27058.02 | 7.343117 |
| 83. | Phenylalanine and derivatives | C9H11NO2 | [M-H]- | 164.0717 | 294260.7 | 1.80225 |
| 84. | Hydroxy fatty acids | C7H12O5 | [M-H]- | 175.0614 | 245560.1 | 4.853917 |
| 85. | Flavonoid-7-O-glycosides | C27H30O15 | [M-H]- | 593.2231 | 74361.05 | 2.003733 |
| 86. | Glutamine and derivatives | C28H38N2O7 | [M+H]+ | 515.2795 | 39850.05 | 9.115617 |
| 87. | Naphthopyranones | C14H8O6 | [M-H]- | 271.0136 | 39124.95 | 10.23745 |
| 88. | Lipids | C27H54NO12P | [M-H]- | 614.3338 | 71140.57 | 8.542883 |
| 89. | Lipids | C26H50NO7P | [M+H]+ | 520.3397 | 42430.07 | 14.5195 |
| 90. | Oxidized fatty acids | C18H32O4 | [M-H]- | 311.2226 | 2047702 | 13.38845 |
| 91. | Alpha amino acids and derivatives | C3H7N3O2 | [M-H]- | 116.0515 | 48463.46 | 6.85615 |
| 92. | BHAs and derivatives | C4H6O5 | [M+H]+ | 135.0305 | 49891.13 | 1.683867 |
| 93. | 7-O-Methylated flavonoids | C19H18O7 | [M-H]- | 357.1031 | 19128.16 | 1.160783 |
| 94. | Biphenyls and derivatives | C18H22O6 | [M-H]- | 333.1292 | 47906.8 | 5.47505 |
| 95. | 2’-Hydroxy-dihydrochalcones | C15H14O4 | [M-H]- | 257.1035 | 56146.97 | 8.030416 |
| 96. | Long-chain fatty acids | C16H32O2 | [M-H]- | 255.2331 | 19809.23 | 20.24078 |
| 97. | Hydroquinolones | C9H7NO | [M-H]- | 144.0465 | 68995.61 | 8.116533 |
| 98. | Leucine and derivatives | C6H13NO2 | [M-H]- | 130.0873 | 155203.4 | 1.6411 |
| 99. | Naphthofurans | C20H28O3 | [M+Na]+ | 339.1891 | 14199.97 | 15.7706 |
| 100. | 3-Alkylindoles | C9H9NO | [M+H-H2O]+ | 130.0659 | 29115.12 | 8.422833 |
| 101. | Linoleic acid and derivatives | C18H32O2 | [M-H]- | 279.2327 | 149488 | 19.10913 |
| 102. | Long-chain fatty acids | C18H34O2 | [M-H]- | 281.2493 | 47593.36 | 20.36357 |
| 103. | Ketals | C36H61NaO11 | [M+H]+ | 694.389 | 39496.04 | 1.724517 |
| 104. | 1,4-Benzodiazepines | C17H14N2O2 | [M-H]- | 277.1546 | 24834.18 | 6.61215 |
| 105. | Hydroxybenzaldehydes | C23H29ClO4 | [M-H]- | 403.2713 | 22729.26 | 9.914967 |
| 106. | Oxidized fatty acids | C18H34O4 | [M-H]- | 313.2387 | 239890.3 | 12.70548 |
| 107. | Stilbenes | C14H12O3 | [M-H]- | 227.1403 | 164394.3 | 1.92325 |
| 108. | Indoles | C9H9N | [M+H]+ | 132.082 | 41733.77 | 6.142617 |
| 109. | Chromones | C13H14O4 | [M-H]- | 233.0757 | 17061.21 | 1.160783 |
| 110. | Naphthofurans | C20H34O4 | [M+Na]+ | 361.2313 | 24442.77 | 14.09052 |
| 111. | Cinnamaldehydes | C9H8O2 | [M-H]- | 147.0448 | 21781.39 | 1.80225 |
| 112. | Alkaloids | C23H30N2O4 | [M-H]- | 397.2198 | 173761.3 | 10.35678 |
| 113. | Linear diarylheptanoids | C19H20O5 | [M-H]- | 327.1941 | 14524.31 | 7.5861 |
| 114. | N-acyl amines | C20H36N4O8 | [M+H]+ | 483.2491 | 54466.5 | 14.5195 |
| 115. | Histidine and derivatives | C9H15N3O2S | [M-H]- | 228.0879 | 32241.37 | 6.287017 |
| 116. | Alpha amino acids and derivatives | C4H6N2O2 | [M+H]+ | 115.0565 | 87360.12 | 6.142617 |
| 117. | Indoles and derivatives | C24H26N2O2 | [M+H]+ | 375.2136 | 41256.94 | 10.32722 |
| 118. | Flavins | C17H20N4O6 | [M-H]- | 375.1325 | 46508.74 | 6.124183 |
| 119. | Phenylpiperidines | C22H28N2O2 | [M+H]+ | 353.2285 | 21989.78 | 10.28705 |
| 120. | Oxidized fatty acids | C9H18O3 | [M-H]- | 173.1177 | 15370.58 | 8.259733 |
| 121. | Cerveratrum-type alkaloids | C27H43NO3 | [M+H]+ | 452.3127 | 247793.1 | 8.875134 |
| 122. | Indoles | C8H7N | [M+H]+ | 118.0668 | 90743.75 | 2.753133 |
| 123. | Organic acids | C10H18O5 | [M-H]- | 217.1077 | 66271.49 | 7.343117 |
| 124. | Cardenolide glycosides and derivatives | C29H44O9 | [M+H]+ | 559.2831 | 18728.07 | 6.142617 |
| 125. | Aconitane-type diterpenoid alkaloids | C25H41NO7 | [M+H]+ | 468.3044 | 41632.81 | 7.88835 |
| 126. | Phosphocholines | C5H15NO4P | [M+H]+ | 184.0741 | 73973.06 | 6.547433 |
| 127. | Alpha amino acids and derivatives | C3H6N2O2 | [M+H]+ | 103.0542 | 177650 | 1.88935 |
| 128. | Glutamic acid and derivatives | C5H9NO4 | [M-H]- | 146.0446 | 32350.85 | 1.200617 |
| 129. | Alpha amino acids | C4H7NO2 | [M+H]+ | 102.0476 | 43224.8 | 1.88935 |
| 130. | Ketals | C26H | [M+H]+ | 429.2699 | 39331.47 | 8.9153 |
| 131. | Tyrosine and derivatives | [M+H]+ | 198.0653 | 29220.79 | 6.102283 | |
| 132. | BHAs and derivatives | 36O5 | [M-H]- | 133.0139 | 338616.7 | 1.320283 |
| 133. | Alkaloids and derivatives | C9H11NO4 | [M+H]+ | 209.1313 | 18565.44 | 12.57427 |
| 134. | Strychnos alkaloids | C4H6O5 | [M-H]- | 353.1841 | 30656.95 | 14.8629 |
| 135. | Kaurane diterpenoids | C11H16N2O2 | [M+H]+ | 376.2572 | 230025.8 | 12.57427 |
| 136. | Benzazepines | C21H26N2O3 | [M+H]+ | 412.2065 | 19489.04 | 8.543317 |
| 137. | Sulfinic acid | C22H33NO4 | [M+H]+ | 110.0358 | 21117.81 | 1.683867 |
| 138. | Aconitane-type diterpenoid alkaloids | C24H29NO5 | [M+H]+ | 466.2893 | 61282.63 | 7.807367 |
| 139. | Pyridines and derivatives | C2H7NO2S | [M+H]+ | 184.0741 | 129057.4 | 8.503317 |
| 140. | Fluorenes | C25H39NO7 | [M+H]+ | 432.2973 | 23864.71 | 11.78715 |
| 141. | Dicarboxylic acid and derivatives | C10H11NO | [M-H]- | 115.0043 | 89568.43 | 1.320283 |
| 142. | Tricarboxylic acid and derivatives | C27H39NO2 | [M-H]- | 191.0196 | 388035.5 | 1.681083 |
| 143. | Naphthopyrans | C4H4O4 | [M-H]- | 441.2347 | 21192.03 | 6.246 |
| 144. | Indole-3-acetic acid derivatives | C6H8O7 | [M-H]- | 174.0554 | 17986 | 7.38395 |
Figure S1. Full-scan ion abundance profile chromatogram in different ion modes of L. edodes and L. decastes. (A) Positive ion mode chromatogram of L. edodes, (B) negative ion mode chromatogram of L. edodes.
Figure S1. Full-scan ion abundance profile chromatogram in different ion modes of L. edodes and L. decastes. (C) positive ion mode chromatogram of L. decastes, and (D) negative ion pattern chromatogram of L. decastes.