
Download

ORIGINAL ARTICLE
Anti-aging activities of black highland barley bran, chrysoeriol, and its glucuronide derivative on Caenorhabditis elegans
Gang Liu1†, Shuyuan Fan1†, Peng Zeng2, Yunshuang He3, Yuehan Shen3, Changbin Lin4, Chen Xia4*, Peijun Li5, Xueli Li3*
1College of Life Science, Sichuan Normal University, Chengdu, China;
2Solid-state Fermentation Resource Utilization Key Laboratory of Sichuan Province, Yibin University, Yibin, China;
3School of Laboratory Medicine, Chengdu Medical College, Chengdu, China;
4Institute of Agro-Products Processing Science and Technology, Sichuan Academy of Agricultural Sciences, Chengdu, China;
5School of Food and Biological Engineering, Hefei University of Technology, Hefei, China
†These authors contributed equally and shared first authorship.
Abstract
The anti-aging activities and underlying mechanisms of black highland barley bran extract (BHBBE) and its major phenolic components, chrysoeriol (CHR) and chrysoeriol-7-O-β-D-glucuronide (CHRG), were investigated in Caenorhabditis elegans (C. elegans). The effects of BHBBE, CHR, and CHRG at different concentrations were evaluated on lifespan, reproduction, locomotion, stress resistance (heat and ultraviolet [UV] stress), and antioxidant activities (such as superoxide dismutase and catalase activities as well as reactive oxygen species (ROS) and malondialdehyde levels). Furthermore, the expression of longevity-associated genes was examined. Central gene, daf-16, in the anti-aging process of BHBBE, CHR, and CHRG was verified by comparing ROS levels in wild-type N2 C. elegans with daf-16(mu86) mutant strains. Nuclear translocation as a function of DAF-16 protein was assessed. BHBBE, CHR, and CHRG significantly extended C. elegans’ lifespan, improved locomotion, and enhanced stress resistance, without adversely affecting reproduction. Their anti-aging effects were primarily mediated by DAF-16 nuclear translocation via insulin/insulin-like growth factor 1 signaling (IIS). Furthermore, this IIS signaling pathway was associated with antioxidant capacity and stress resistance. Therefore, BHBB, a by-product of grain processing, provides a sustainable, safe, and efficient strategy for anti-aging applications.
Key words: black highland barley bran, Caenorhabditis elegans, longevity, IIS/DAF-16 pathway, antioxidant
*Corresponding Authors: Chen Xia, Institute of Agro-Products Processing Science and Technology, Sichuan Academy of Agricultural Sciences, Chengdu, China. Email: [email protected]; Xueli Li, School of Laboratory Medicine, Chengdu Medical College, Chengdu, China. Email: [email protected]
Academic Editor: Hamit Koksel, PhD, Department of Food Engineering, Hacettepe University, Turkey
Received: 24 March 2025; Accepted: 11 December 2025; Published: 28 February 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Highland barley (Hordeum vulgare L. var. nudum hook. f), a variant of the grass family, is commonly referred to as naked barley because of its hull-less grains. This important cereal crop is primarily produced in the Qinghai-Tibet Plateau and in other Asian regions, such as China, Japan, Nepal, and India (Meints et al., 2021). In China, highland barley serves as a staple food for residents in high-altitude areas and is traditionally prepared as Tsampa, a roasted barley flour dish, also known as Zanba in Chinese (Obadi et al., 2021). Highland barley, as a nutrient-rich cereal, is widely utilized in brewing, baking, and functional foods (Geng et al., 2022; Lukinac and Jukić, 2022). It has demonstrated significant antioxidant properties, even stronger than common crops, because of its richness in phenolic compounds (Obadi et al., 2021). Among black, white, and blue barley cultivars, black highland barley exhibited the highest polyphenol levels (Yang et al., 2023). This variety was characterized by its black husk (Zhang et al., 2023). Notably, barley bran contains higher polyphenol levels, compared to peeled barley, with black highland barley bran (BHBB) recording a total phenol content of 237.83 mg/g (Blandino et al., 2015; Deng et al., 2024).
Black highland barley bran extract (BHBBE) and its two main components possess robust antioxidant capacity (Demirtas et al., 2013; Deng et al., 2021; Xiang et al., 2021). Polyphenols, as antioxidants, may mitigate aging by eliminating free radicals and modulating key genes in oxidative stress-related pathways. Moreover, these natural polyphenols have shown significant potential for market application (Ding et al., 2017; Okoro et al., 2021). Consequently, the anti-aging effects of BHBBE and its essential polyphenols, chrysoeriol-7-O-β-D--glucuronide (CHRG) and chrysoeriol (CHR), are investigated in this study.
Caenorhabditis elegans (C. elegans) served as a -frequently-used anti-aging model because of its short lifespan, and easily observed and conserved longevity pathways (Herndon et al., 2002). Lifespan extension in C. elegans led to various physiological changes, including enhanced resistance to heat and ultraviolet (UV) stress and reduced levels of reactive oxygen species (ROS) (Wang et al., 2020).
Genetic mechanisms associated with anti-aging in C. elegans are the modulation of insulin/insulin-like growth factor 1 (IGF-1) signaling pathway (IIS) and p38 mitogen-activated protein kinase (MAPK) pathway (Zhang et al., 2020). Extensive research has highlighted the pivotal role of these two pathways in regulating aging and disease processes (How et al., 2021; Park et al., 2021; Xiao et al., 2024; Ziv and Hu, 2011). Key genes, such as age-1 and daf-16, along with major gene regulators, such as sir-2.1, sek-1 and skn-1, are involved in these two pathways. Downstream genes, such as sod-3 and ctl-1, influenced the antioxidative capacity of C. elegans by regulating activities of enzymes, such as superoxide dismutase (SOD) and catalase (CAT).
Therefore, the anti-aging effects of BHBBE, CHR, and CHRG on C. elegans were investigated, including its lifespan, reproductive capacity, stress tolerance (heat and UV resistance), and antioxidant ability. The underlying mechanisms were further explored by examining the nuclear translocation of DAF-16 and analyzing the expression of key aging-related genes, such as sir-2.1, age-1, daf-16, sod-3, ctl-1, sek-1, and skn-1. Additionally, the critical role of the DAF-16 pathway was specifically verified by monitoring ROS levels in daf-16 (mu86) mutant strains under hydrogen peroxide (H2O2) stress. This investigation provides important insights into the anti-aging activity and antioxidant mechanism of BHBB, which may promote its future application in functional foods and nutritional interventions.
Materials and Methods
Materials
Black highland barley bran was provided by Tibet Chun Guang Food Co. Ltd. (Tibet, China). The C. elegans wild-type strain N2, TJ356 strain zIs356 (daf-16p::daf-16a/b::GFP + rol-6[su1006]), and daf-16 (mu86) mutant were obtained from the Caenorhabditis Genetics Center (CGC) (University of Minnesota, Minneapolis, MN, USA). The bacterial food source, Escherichia coli OP50 (E. coli OP50), was also procured from the CGC. Assay kits for SOD (Catalog No. A001-3-2), CAT (Catalog No. A007-1-1), and malondialdehyde (MDA; Catalog No. A003-1-2) were purchased from the Nanjing Institute of Biological Engineering (Nanjing, China). Fluorescent probe 2’,7’-dichloro-dihydrofluorescein diacetate (DCFH-DA; Catalog No. D6883) for ROS detection was obtained from Sigma-Aldrich (St. Louis, MO, US). Additionally, a bicinchoninic acid (BCA) protein assay kit (Catalog No. A045-4-2) for sample normalization was sourced from the Nanjing Jiancheng Bioengineering Institute. All other chemicals and reagents, including AB-8 macroporous resin, were supplied by Chengdu Chron Chemicals Co. Ltd. (Chengdu, China).
Preparation of extracts
Black highland barley bran (2,500 g) was extracted three times with 10 L of 80% ethanol at 70°C. The combined filtrates were concentrated at 50°C to yield BHBBE. The resulting BHBBE was dissolved in an equal volume of distilled water and loaded onto an AB-8 macroporous resin column. The polyphenols CHR and CHRG were separated from BHBBE by step-gradient elution with 0%, 20%, 40%, and 93% (v/v) aqueous ethanol. Fractions were monitored by high-performance liquid chromatography (HPLC), and those enriched in CHR or CHRG were combined and concentrated. Subsequently, 110 mg of CHR and 610 mg of CHRG were yielded after freeze--drying. The CHR and CHRG was identified using an Agilent 1290 Infinity III HPLC system (Agilent Technologies, USA). The purification was performed by preparative HPLC using a reversed-phase C18 column (XB-C18, 10 μm, 10 nm) maintained at room temperature. The separation was achieved under isocratic conditions with a mobile phase comprising acetonitrile and 0.1% formic acid (35:65 v/v) delivered at a flow rate of 1.0 mL/min. Detection was carried out with a UV detector set at 350 nm. For each injection, a volume of 0.4 mL was loaded. The structures of CHR and CHRG were characterized by nuclear magnetic resonance (NMR) spectroscopy. Samples were dissolved in deuterated methanol, and their 1H and 13C NMR spectra were recorded on a Bruker Avance Neo 400 MHz NMR spectrometer at 25°C (Bruker BioSpin, Billerica, MA, USA).
Maintenance and synchronization of C. elegans’ strains
C. elegans was cultured at 20°C on Nematode Growth Medium (NGM) fed with E. coli OP50 that was heat-inactivated (65°C, 30 min). The gravid nematodes were collected after treatment with M9 buffer (KH2PO4 + Na2HPO4 + NaCl). Eggs were released by vigorously shaking gravid nematodes in a solution containing bleach (1 mL 5% NaClO + 0.5-mL 0.5-M NaOH) for 10 min. After centrifugation to remove bleach, eggs were washed for three times with M9, and incubated overnight on NGM plates seeded with E. coli OP50. Synchronized L4 nematodes were collected after 48 h of incubation.
According to our previous findings (Wang et al., 2023), BHBBE, CHR, and CHRG exhibited optimal in vitro antioxidant activity at concentrations ranging from 200 to 1,000 μg/mL; therefore, concentrations of 200, 500, and 1,000 μg/mL were selected for subsequent experiments.
Lifespan assay
C. elegans was treated with varying concentrations of BHBBE, CHR, and CHRG to evaluate their impact on lifespan. Synchronized L4 nematodes were transferred to NGM plates supplemented with E. coli OP50, 50-µM 5-Fluorouracil 2’’-deoxyriboside (FUdR), and corresponding concentrations (200, 500, and 1,000 μg/mL) of BHBBE, CHR, and CHRG at 20°C, with 30 nematode specimens per plate. FUdR could prevent the reproduction of nematodes without interfering with their development and aging after maturation. Daily observations were made to record the number of deaths until the death of last nematode, and the experiment was repeated thrice. C. elegans was considered dead if they did not move or respond within 10 s on being lightly touched with a platinum wire. Nematodes that escaped from the culture dish or died during the experiment were excluded from data analysis.
Fecundity assay
In light of the positive relationship between lifespan extension and reproductive capacity in aging research (Son et al., 2011), the effects of BHBBE and its major components were investigated on the fecundity of C. elegans. The L4 stage nematodes were treated using the same method as done in the lifespan assay. Nematodes were divided into four groups, with each group containing three gravid adults. These groups were cultured at 20°C on NGM plates seeded with E. coli OP50. The treatment groups were supplemented with BHBBE, CHR, and CHRG at concentrations of 200, 500 and 1,000 μg/mL, while the control group received no supplementation. Subsequently, they were transferred to fresh NGM plates daily until egg-laying was completed. All egg-laying plates were incubated for 48 h to promote hatching, and the number of recorded larvae was considered equivalent to the number of eggs laid.
Movement assay
C. elegans’ motility was assessed using the body-swing count test. L4 stage nematodes were treated using the same method as done in the lifespan assay at 20°C. This time point was considered the beginning of the experiment. Over the subsequent 7 days, 10 nematodes were randomly selected daily. Using a platinum wire, nematodes were gently stimulated, and the number of swings performed by each nematode in 1 min was recorded. A swing was counted when C. elegan moved from one side to the other, observed either in the head or tail region.
Stress resistance
C. elegans treated with BHBBE, CHR, and CHRG was subjected to acute heat resistance tests. The L4 stage nematodes were treated as described in the lifespan assay and cultured at 20°C for 5 days. Each plate contained 30 nematodes. They were transferred at 37°C on the sixth day until all were dead. Nematode deaths were recorded hourly based on the criteria outlined in the lifespan assay.
Similar to thermotolerance assay, UV stress tests were conducted on 30 randomly selected C. elegans from each group. After 5 days of culture at 20°C, the nematodes were exposed to UV radiation for 2 h on the sixth day. Daily death counts were recorded, and the survival rate was calculated.
Antioxidant enzyme activity and oxidative stress markers
The effects of BHBBE, CHR, and CHRG on the antioxidant activity of C. elegans were evaluated by measuring antioxidant enzyme activities and oxidative damage markers. Synchronized L4-stage nematodes were treated according to the methodology described in the lifespan assay and cultured at 20°C for 5 days. Approximately 1,000 nematodes per group were collected, washed thrice with M9 buffer, and subjected to three rapid freeze-thaw cycles in liquid nitrogen. The samples were then homogenized and centrifuged (12,000 ×g, 4°C, 2 min) to obtain supernatant.
The supernatant was then analyzed using commercial kits to quantify SOD and CAT activities as well as MDA content, and protein concentrations were determined with the BCA method. For ROS detection, 50 μL of supernatant was added to an equal volume of DCFH-DA, and the mixture was incubated for 2 h at 20°C in the dark. Fluorescence was then measured at excitation and emission wavelengths of 485 nm and 538 nm, respectively. ROS levels were expressed as relative fluorescence units (% control).
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
C. elegans was cultured on NGM medium supplemented with 200 μg/mL of BHBBE, CHR, and CHRG to investigate their potential lifespan-extending effects and epigenetic regulation. Minimum 2,000 nematodes were cultured per plate. They were then washed thrice with M9 buffer, and messenger RNA (mRNA) was extracted using the Animal Total RNA Isolation kit (Foregene Co., Ltd., Chengdu, China). The centrifuge protocol was repeated thrice. The concentration and purity of mRNA were assessed using an ultra-micro spectrophotometer. Reverse transcription was conducted using the 5X All-In-One RT MasterMix (with AccuRT Genomic DNA Removal kit) (ABM). RT-qPCR was conducted using EvaGreen Express 2X qPCR MasterMix-No Dye (ABM) following the manufacturer’s instructions. The expression of the daf-16 gene and its downstream target genes was quantified using the act-1 gene as an internal reference. Relative gene expression levels were calculated using the delta delta crossing thresholds (2-ΔΔCT) method. Primer sequences are provided in Table 1.
Table 1. Sequences of gene primers.
| Gene | Gene ID | Forward Primer 5’–3’ | Reverse Primer 5’–3’ |
|---|---|---|---|
| act-1 (reference gene) | 181097 | GTCAGTGGTGGACCTGACCT | AGGGGTCTACATGGCAACTG |
| sek-1 | 181043 | ATGCTCGGTGAGTATTGG | TCATTGATAAACCGAGCC |
| sod-3 | 181748 | GGCTAAGGATGGTGGAGAAC | ACAGGTGGCGATCTTCAAG |
| ctl-1 | 180837 | CGGATACCGTACTCGTGATGAT | CCAAACAGCCACCCAAATCA |
| skn-1 | 177343 | CACGCCGTCAGCGAAGTA | ATGCTCGGTGAGTATTGG |
| age-1 | 174762 | CCTGAACCGACTGCCAATC | GTGCTTGACGAGATATGTGTATTG |
| daf-16 | 172981 | TCAAGCCAATGCCACTACC | TGGAAGAGCCGATGAAGAAG |
| sir-2.1 | 177924 | ACTGAGATGCTCCATGACAATAAG | GCAAGACGAACCACACGAAC |
Note: ROS levels in the daf-16 mutant of C. elegans during oxidative stress.
Intracellular ROS levels were measured in nematodes under oxidative stress conditions in order to investigate the correlation of the daf-16 gene with the antioxidative effects of BHBBE, CHR, and CHRG. The wild-type N2 and the daf-16 (mu86) mutant strains were used in this experiment. The methods for nematode synchronization and treatment with BHBBE, CHR, and CHRG were consistent with those described in the lifespan assay. After 5 days of treatment, nematodes from each group were collected and exposed to 10-μM H2O2 for 2 h to induce acute oxidative stress. The ROS levels were then measured using the same method as described previously.
DAF-16 nuclear localization assay
Visualization of DAF-16 subcellular localization using a DAF-16::GFP reporter strain was employed to examine its potential role in the anti-aging mechanism of BHBBE, CHR, and CHRG. C. elegans was treated and cultured as described in the lifespan assay. After 5 days of treatment at 20°C, nematodes were transferred onto a 2% agarose pad on a microscope slide and paralyzed with 10-mM sodium azide (NaN3). The localization of DAF-16::GFP was classified as nuclear if ≥50% of intestine cells had positive nuclear GFP punctae, intermediate if more than 10%, <50% of intestine cells had positive nuclear GFP punctae, or cytosol if ≤10% of intestine cells had positive nuclear GFP punctae. For each group, at least 50 nematodes were blindly scored for DAF-16::GFP localization and categorized as nuclear, cytosolic, or intermediate. Data are presented as the percentage of animals exhibiting nuclear translocation (Ke et al., 2020).
Statistical analysis
Each experiment comprised three biological replicates. The results were reported as mean ± standard deviation (SD), with the exception of the lifespan and DAF-16 nuclear translocation assays. Lifespan curves were plotted using the Origin 2021 software (Origin Lab Corporation, Northampton, MA, US). One-way ANOVA and Tukey’s test were used to assess the significance of differences in experimental data. Statistical analyses were performed using the SPSS software (SPSS Inc., Chicago, IL, US), with the significance level set at P < 0.05.
Results
Lifespan extension by BHBBE, CHR, and CHRG
Chrysoeriol and CHRG were successfully isolated from BHBBE, and their structures were identified. The isolation was monitored by preparative HPLC (Supplementary Figure S1), and the structural identification was confirmed by NMR spectroscopy (Supplementary Figure S2). Their chemical structures are shown in Figure 1.
Figure 1. (A) Structure of CHR; (B) structure of CHRG.
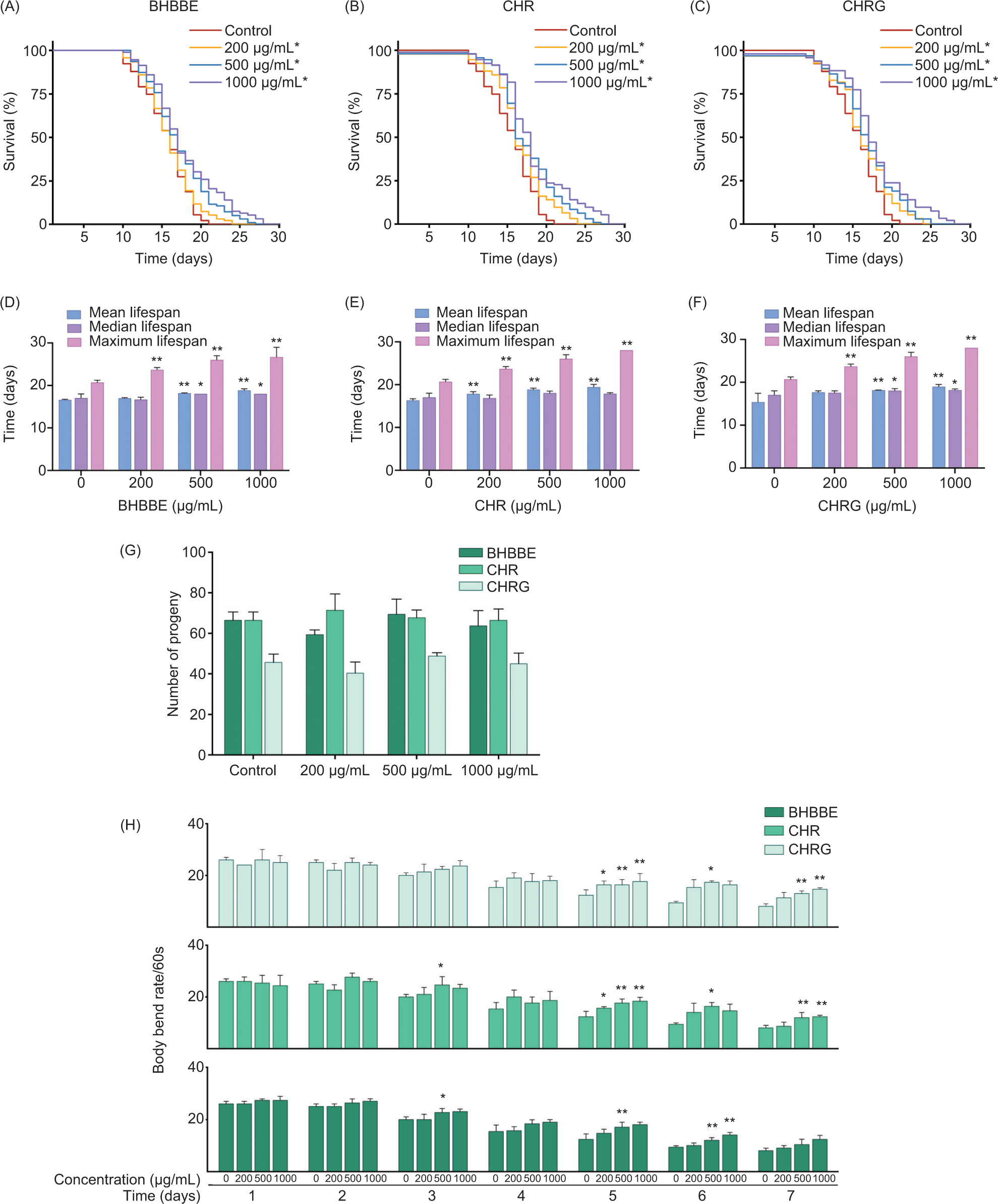
The anti-aging effects of BHBBE, CHR, and CHRG on C. elegans were assessed by recording the lifespan (Figures 2(A–C)). Compared to the control group, the survival curves of nematodes treated with varying concentrations of BHBBE, CHR, and CHRG shifted to the right, indicating that BHBBE, CHR, and CHRG remarkably extended the survival rate of wild-type nematodes in a dose-dependent manner (P < 0.05). Notably, a significant extension was already achieved at a concentration of 200 μg/mL for all three compounds (P < 0.05).
Figure 2. Effects of BHBBE, CHR, and CHRG on lifespan, locomotion, and reproduction of C. elegans. (A–C) Survival curves of C. elegans treated with 200, 500 and 1,000 μg/mL BHBBE, CHR, and CHRG, respectively, exhibiting trends in lifespan -extension compared to the control. (D–F) Impact of BHBBE, CHR, and CHRG on the mean, median, and maximum lifespan of C. -elegans. (G) Number of offspring produced by C. elegans treated with BHBBE, CHR, and CHRG. (H) Frequency of oscillations in C. -elegans treated with BHBBE, CHR, and CHRG. *P < 0.05; **P < 0.01 vs. control group.
Mean and median lifespans reflect the overall trend in lifespan, while the median lifespan is not affected by extreme values. Maximum lifespan reveals potential biological limits. Therefore, in order to comprehensively evaluate the effects of BHBBE, CHR, and CHRG on the lifespan of C. elegans, mean, median, and maximum lifespans of nematodes were assessed. In Figures 2(D–F), the mean lifespan of nematodes treated with 500 and 1,000 μg/mL of BHBBE, CHR, and CHRG was significantly higher than that of the control group (P < 0.01). Remarkably, only CHR at the lowest concentration (200 μg/mL) remarkably extended the mean lifespan (P < 0.01). Both 500 and 1,000-μg/mL BHBBE and CHRG efficiently extended the median lifespan (P < 0.05). All concentrations of BHBBE, CHR, and CHRG significantly extended the maximum lifespan (P < 0.01).
Motility and reproductive capacity
None of the tested compounds (i.e., BHBBE, CHR, and CHRG) showed statistically significant differences in egg-laying number, compared to the control group (P > 0.05), illustrating that BHBBE, CHR, and CHRG did not affect the reproduction of C. elegans (Figure 2G).
The effects of BHBBE, CHR, and CHRG on the motility of C. elegans were assessed by counting body bend frequencies from day 1 to day 7 (Figure 2H). BHBBE and its two major polyphenols improved the motility of nematodes from day 5. BHBBE at 500 μg/mL and 1,000 μg/mL efficiently enhanced motility on day 6 (P < 0.01). Both CHR and CHRG at 500 and 1,000 μg/mL significantly improved motility on day 5 and day 7 (P < 0.01).
Enhancement of heat stress and UV irradiation resistance
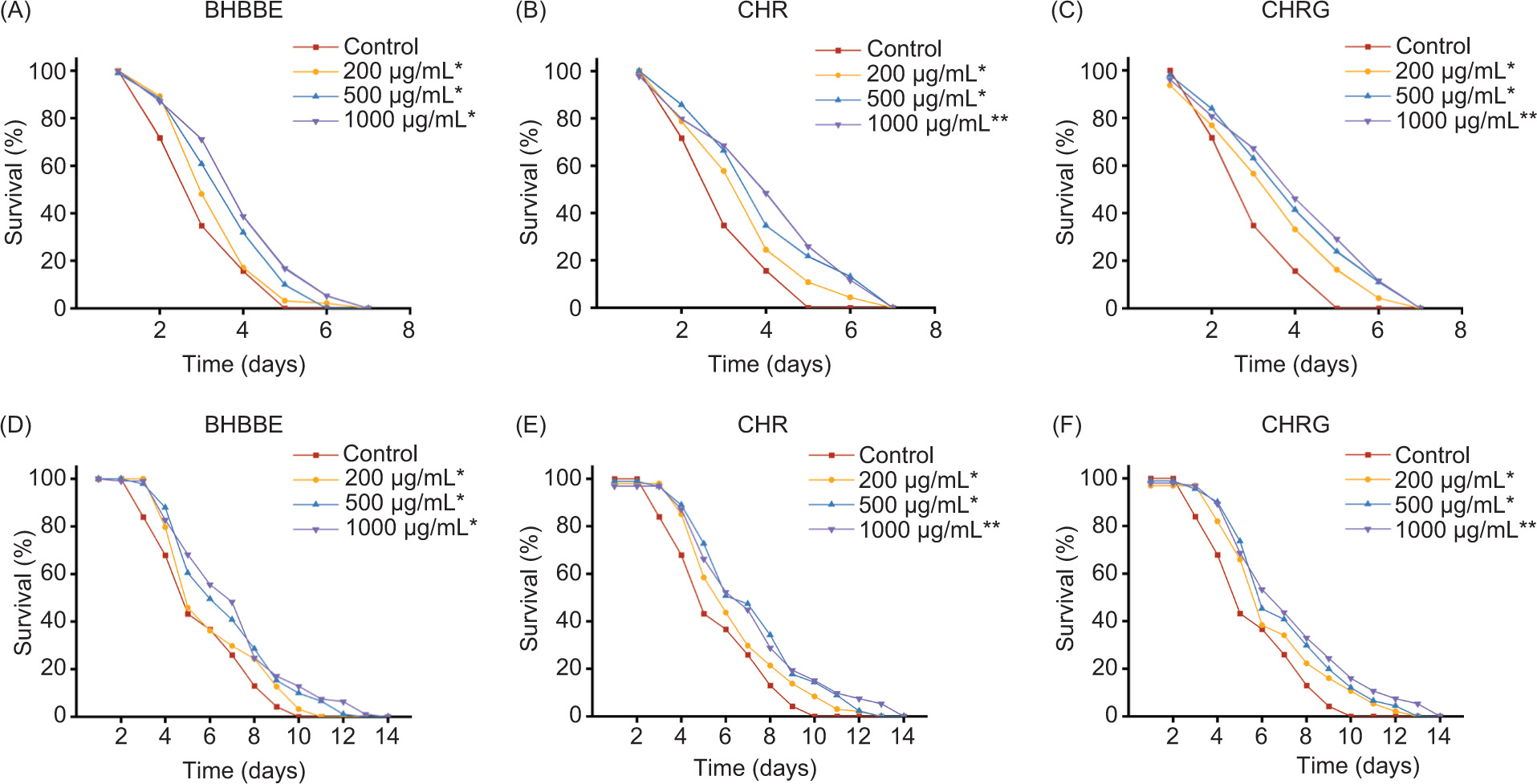
The resistance of C. elegans to both heat and UV stress was significantly enhanced by treatment with BHBBE, CHR, and CHRG (Figure 3). A progressive increase in lifespan was observed with increasing concentration, and significant prolonging in the lifespan of stressed nematodes by BHBBE, CHR, and CHRG was observed from 200 μg/mL (P < 0.05). Given that, 200 μg/mL of BHBBE and its two polyphenols were effective in prolonging of lifespan and improving stress resistance, this concentration was subsequently selected for mechanistic studies.
Figure 3. Survival rate of C. elegans treated with BHBBE, CHR, and CHRG under (A–C) heat stress, or (D–F) UV irradiation. *P < 0.05; **P < 0.01 vs. control group.
Enhancement of antioxidant capacity
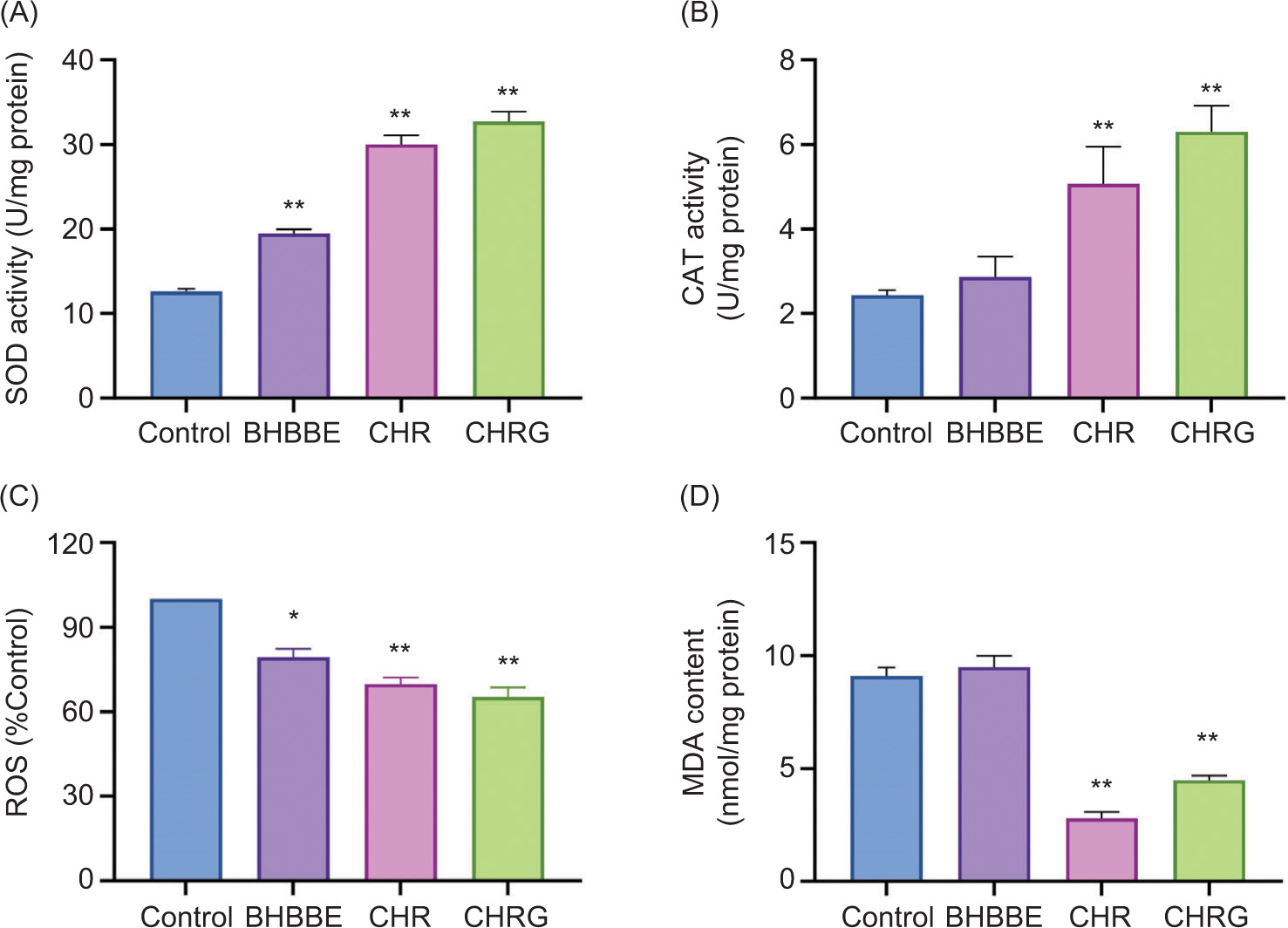
To explore potential biochemical basis for these phenotypic improvements, the basal activity of key antioxidant enzymes and the level of oxidative damage were measured in cultured nematodes. As shown in Figure 4A, treatment with BHBBE, CHR, and CHRG significantly increased SOD activity in C. elegans by 54.8%, 143.6%, and 159.8%, respectively (P < 0.05), compared to the untreated control. Similarly, CAT activity was also enhanced (Figure 4B) with increases of 17.9% (ns), 108.5% (P < 0.05), and 159.5% (P < 0.05). In addition, ROS levels were significantly reduced by BHBBE, CHR, and CHRG, showing the respective decrease of 20.6% (P < 0.05), 30.2% (P < 0.01), and 34.8% (P < 0.01) (Figure 4C). Regarding MDA content, CHR and CHRG significantly reduced it by 69.4% and 50.8%, respectively (P < 0.05), while BHBBE did not cause a significant change in MDA (Figure 4D).
Figure 4. Antioxidant activity of N2 C. elegans treated with 200-μg/mL BHBBE, CHR, and CHRG. (A) SOD activity. (B) CAT -activity. (C) ROS level. (D) MDA content. *P < 0.05; **P < 0.01 vs. control group.
Gene expression analysis in C. elegans
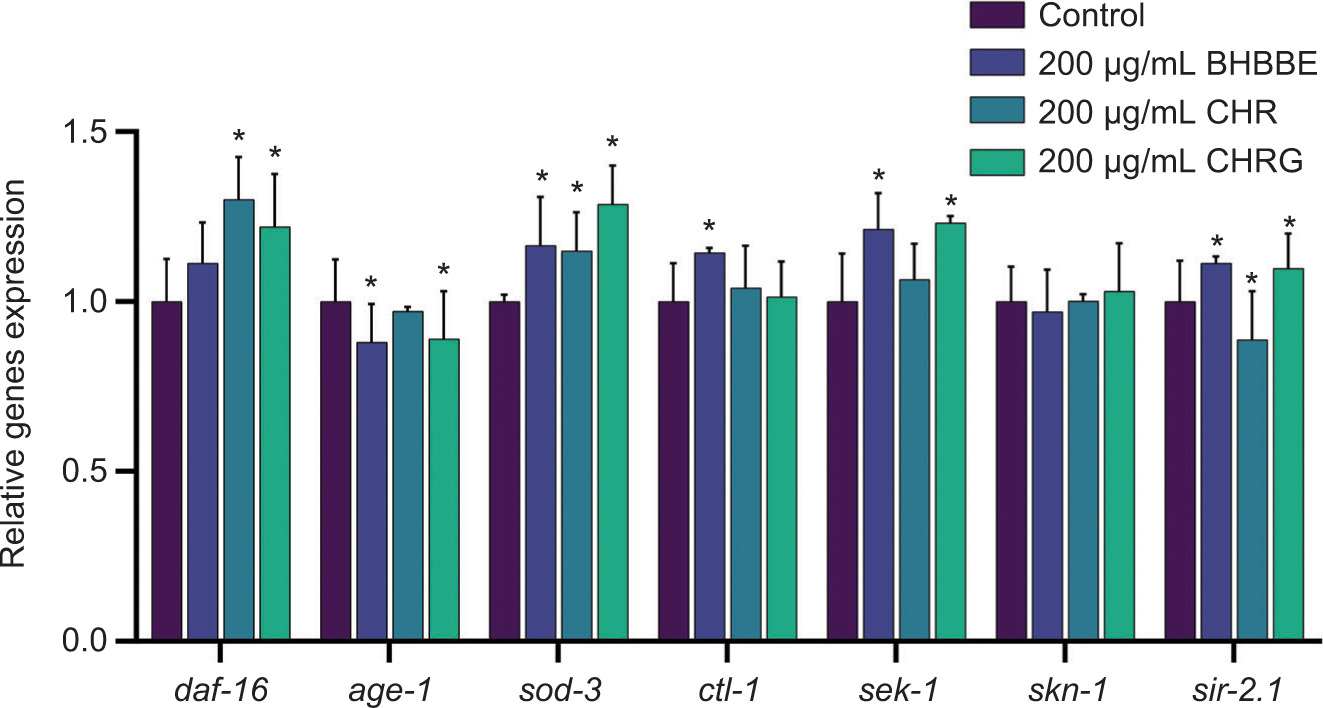
Expressions of the longevity-related genes in C. elegans, such as daf-16, age-1, sod-3, ctl-1, sek-1, skn-1, and sir-2.1, were adjusted by BHBBE, CHR, and CHRG (Figure 5). Specifically, compared to the control group, expression of the sod-3 gene was significantly increased by 1.17-fold, 1.15-fold, and 1.29-fold following respective BHBBE, CHR and CHRG treatments (P < 0.05). Expression of the sir-2.1 gene was remarkably increased by 1.11-fold and 1.10-fold by respective BHBBE and CHRG treatments (P < 0.05), while it was significantly decreased to 88.86% following CHR treatment (P < 0.05). Expression of the sek-1 gene was efficiently increased by 1.21-fold and 1.23-fold by respective BHBBE and CHRG treatments (P < 0.05). Expression of the daf-16 gene was remarkably increased by 1.30-fold and 1.22-fold by respective CHR and CHRG treatments (P < 0.05). Besides, expression of the age-1 gene was efficiently decreased to 88.00% and 89.00% (P < 0.05) by respective BHBBE and CHRG treatments. Expression of the ctl-1 gene remarkably increased by 1.14-fold by BHBBE treatment (P < 0.05). No significant change was observed in the expression of the skn-1 gene. Overall, treatments with BHBBE, CHR, and CHRG influenced the lifespan of C. elegans by modulating the expression of genes in aging pathways.
Figure 5. Effects of 200-μg/mL BHBBE, CHR, and CHRG on the expression of longevity-related genes in C. elegans. *P < 0.05; **P < 0.01 vs. control group.
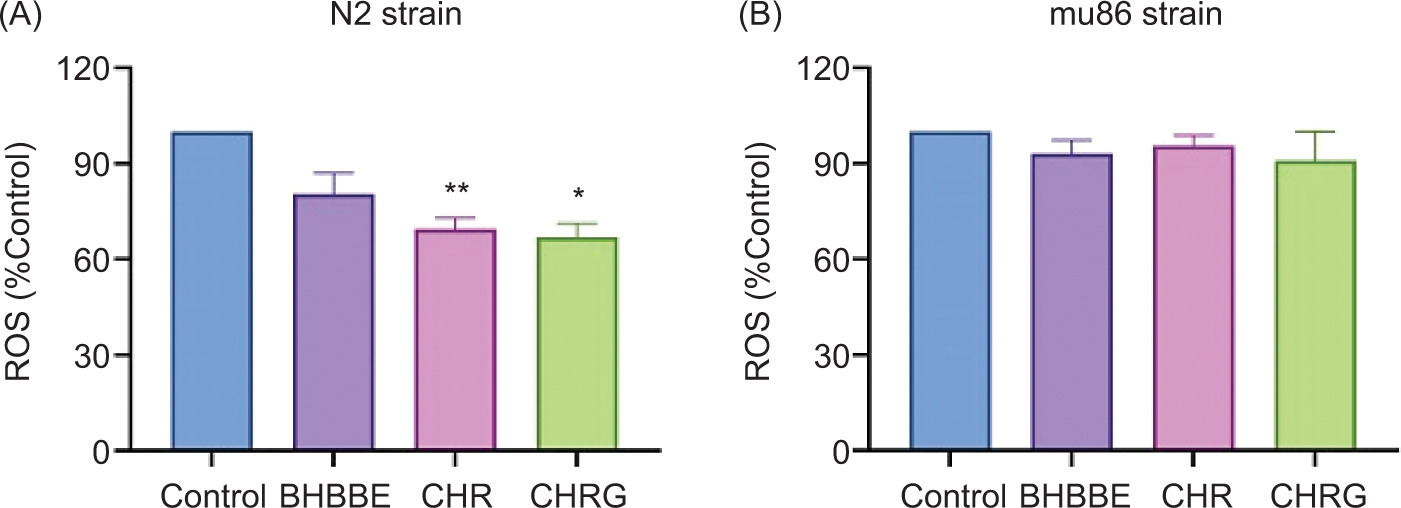
ROS levels in daf-16 mutant C. elegans under oxidative stress
Role of the crucial gene daf-16 was further verified by comparing N2 wild-type and daf-16 (mu86) mutant C. elegans under H2O2 stress (Figure 6). Results demonstrated that the antioxidative capacity of BHBBE, CHR, and CHRG was largely dependent on the daf-16 gene. The efficacy of BHBBE, CHR, and CHRG in attenuating oxidative stress was evident in N2 C. elegans, as shown by the respective reduction of 19.6%, 30.6%, and 33.1% in fluorescence intensity, compared to the control group (Figure 7A). In contrast, daf-16 mutant nematodes exhibited no significant differences in fluorescence -intensity between the treatment and control groups (Figure 7B). These results suggest that the antioxidant effects of BHBBE, CHR, and CHRG appeared to be abolished in the absence of the functional daf-16 gene.
Figure 6. Effects of BHBBE, CHR, and CHRG on ROS levels in (A) wild-type N2, and (B) DAF-16 (mu86) mutant C. elegans under H2O2-induced oxidative stress. *P < 0.05; **P < 0.01 vs. control group.
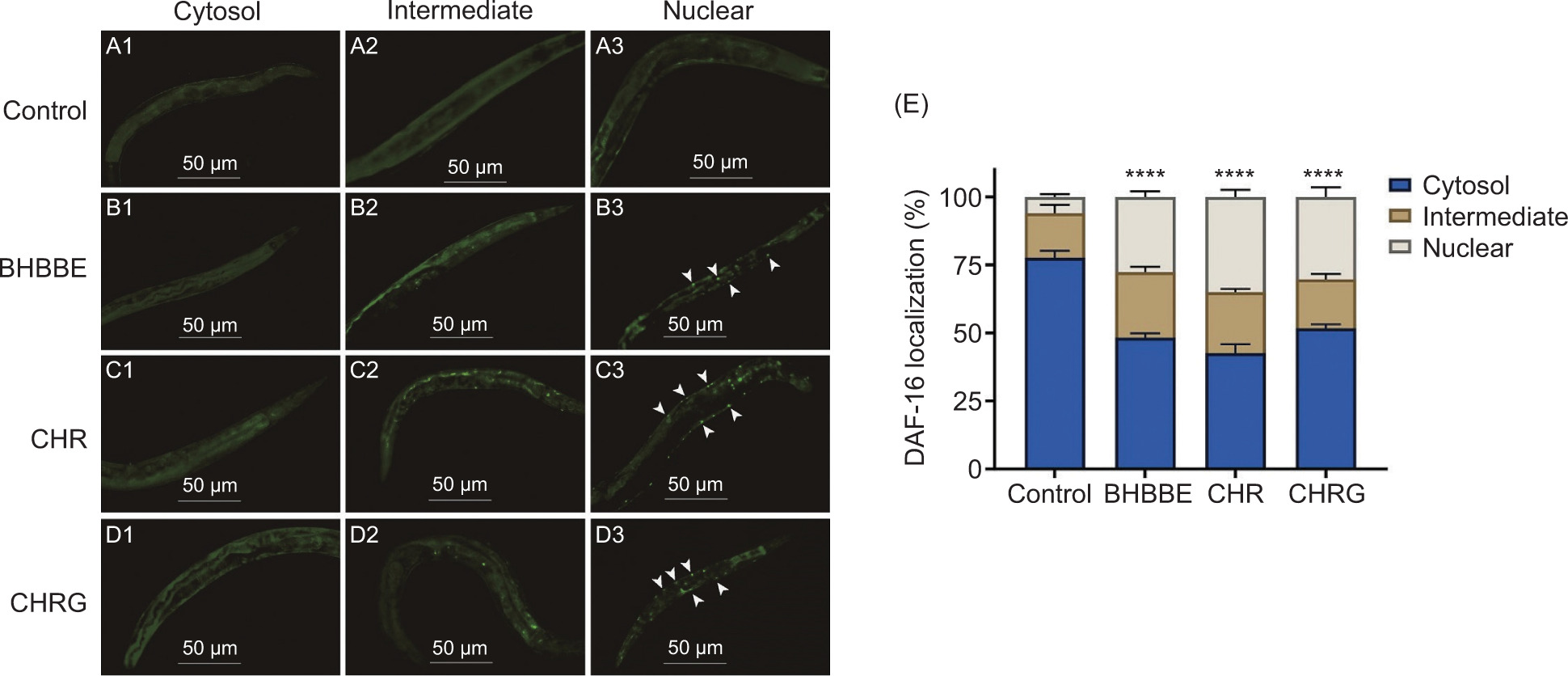
Figure 7. DAF-16 nuclear translocation in C. elegans. (A–D) DAF-16::GFP subcellular distribution (cytosolic, intermediate, and nuclear) in C. elegans treated with BHBBE, CHR, and CHRG. (E) Quantitative analysis of DAF-16 localization, presented as the percentage of C. elegans with DAF-16 predominantly localized to cytosol, intermediate, or nucleus. ****P < 0.0001 (for nuclear) vs. control group.
DAF-16 nuclear localization
The TJ356 strain contains a DAF-16::GFP fusion protein, which contributes to improving the detectability of DAF-16 protein. The effects of BHBBE, CHR, and CHRG on the localization of DAF-16 protein are shown in Figure 7. DAF-16::GFP was uniformly distributed throughout the cytoplasm in control C. elegans (Figure 7A). In contrast, treatment with 200-μg/mL BHBBE, CHR, and CHRG induced its nuclear translocation, as evidenced by the formation of bright nuclear puncta (white arrows, Figures 7B–D). Quantification revealed that all three treatments significantly increased the proportion of animals exhibiting DAF-16 nuclear translocation, compared to the control (P < 0.0001; Figure 7E). Among them, CHR induced the most pronounced nuclear accumulation, followed by CHRG and BHBBE.
Discussion
Highland barley, a distinct crop native to the plateau regions of China, possesses a range of functionalities, including the treatment of chronic and metabolic ailments and the enhancement of antioxidant activity (Guo et al., 2020; Li et al., 2022). These nutritional and health advantages can be attributed to the richness of phenolic compounds in highland barley, with approximately 80% of total phenols concentrated in the bran and embryo (Zhu et al., 2015). Thus, phenolic compounds in highland barley bran have a great potential for utilization (Zhang et al., 2021). Dominant phenols, such as CHR and CHRG, were found in all 14 varieties of highland barley bran in our earlier research and demonstrated the remarkable antioxidant properties of CHR and CHRG (Deng et al., 2021). The antioxidant property closely correlates with delaying aging (da Costa et al., 2016). The anti-aging and antioxidant abilities were strongly related to the treatment of different chronic ailments, such as osteoporosis, neurodegeneration, heart disease, diabetes, and cancer (Higgins-Chen et al., 2021; Rodrigues et al., 2021). Consequently, the anti-aging impact of BHBBE and its dominant phenols, such as CHR and CHRG, was assessed on C. elegans, and their possible mechanisms were discussed.
The mean lifespan of C. elegans was extended by BHBBE, CHR, and CHRG, with CHR showing the strongest effect. Compared to CHR, O-glycosylation occurring at C7 position in CHRG may substantially alter its biological activity. Hydroxylation at C7 position impeded the interaction with peroxide radicals, resulting in a significant reduction in the antioxidant activity of phenolic compounds (Mishra et al., 2003). Glycosylation was demonstrated to diminish the anticancer activity of CHR (Takasawa et al., 2008). The negative effect potentially stemmed from the substitution of hydroxyl groups with bulky sugar moieties, which led to a reduction of active hydroxyl groups and an increase of steric hindrance. Thus, decline in the anti-aging activity of CHRG might be caused by its glycosylation. CHR, as an aglycone of CHRG, showed a stronger lifespan extension effect, compared to CHRG. The lowest lifespan extension effect was observed in BHBBE. The bran extracts contained 10.85% of CHR, 82.19% of CHRG, and other compounds, which may affect BHBBE’s lifespan extension, compared to pure CHR and CHRG.
A close correlation was observed between reproduction and lifespan (Baxi et al., 2017). Results demonstrated that the lifespan of C. elegans was extended by BHBBE, CHR, and CHRG, while C. elegans’ progeny numbers were not affected remarkably. This effect of extending nematodes’ lifespan without compromising reproductive health was akin to that observed with epigallocatechin-3-gallate (Song et al., 2024). Additionally, polyphenols from other traditional medicinal plants showed the same effect. For instance, extracts from Glochidion zeylanicum leaves remarkably extended the lifespan of nematodes without impacting their reproductive capacity (Duangjan et al., 2019).
The aging of C. elegans was accompanied by a gradual decline in its motility, which reflected the health status of nematodes (Hsu et al., 2009; Li et al., 2019). The locomotion of nematodes was observed to slow down with aging. BHBBE, CHR, and CHRG enhanced C. elegans’ locomotion, evidently after 5 days of treatment. Epigallocatechin-3-gallate increased the body bend frequency, head thrash ability, and pharyngeal pumping rate of nematodes (Brown et al., 2006). Olive polyphenols also had a strong beneficial effect on the locomotion of C. elegans (Di Rosa et al., 2020). Thus, polyphenols such as CHR, CHRG, and BHBBE that improved motility and anti-aging effect had a promising application in functional foods.
Besides, BHBBE and its two main polyphenols resisted the aging of C. elegans triggering from environmental stress. BHBBE, CHR, and CHRG could still extend the lifespan of nematodes after exposure to 37°C heat stress or UV irradiation for 2 h. Blueberry polyphenols also enhanced stress resistance and extended the lifespan of C. elegans (Wang et al., 2018).
Oxidative activity played a crucial role in C. elegans aging and age-related diseases, such as neurodegenerative diseases, cardiovascular diseases, diabetes, and cancers (Maglioni and Ventura, 2016; Salmon et al., 2010). The accumulation of ROS and its damage is the primary cause of aging according to free radical theory (Brintz, 2020). SOD and CAT, as key antioxidant enzymes, protected cells by eliminating ROS or other oxidative products (Jomova et al., 2023). Excessive free radical induced lipid peroxidation, and thus produced oxidative products such as MDA. High SOD and CAT activity indicated strong free radical scavenging ability, while low MDA showed reduced free radicals and promoted antioxidant effect.
The daf-16 gene may be the pivotal gene for the anti-aging and anti-oxidative effects of BHBBE and its two polyphenols. Because ROS levels were significantly reduced by two phenols in wild-type N2 nematodes under oxidative stress, no notable change was observed in the daf-16 mutants. The daf-16 gene is a crucial one in the IIS pathway. The IIS pathway was a highly conserved aging progress that controlled stress resistance and lifespan of C. elegans (Morley et al., 2002). The age-1 gene negatively regulated the daf-16 gene expression, while sir-2.1 as a member of Sirtuin family, positively regulated DAF-16 protein translocation (Kobayashi et al., 2005; Zečić and Braeckman, 2020). Expressions of the age-1 gene could be inhibited by up-regulating the sir-2.1 gene by BHBBE, CHR, and CHRG, which show the possible anti-aging mechanism of IIS pathway. Improvement in BHBBE, CHR, and CHRG in DAF-16 nuclear translocation was also observed. The DAF-16 transcription factor, an ortholog of human FOXO proteins in IIS pathway, was capable to activate the expression of the sod-3 and ctl-1 genes (Motta et al., 2020; Sun et al., 2017; Wu et al., 2012). These activated genes up-modulated the activity of anti-oxidative enzymes SOD and CAT that decreased oxidative products such as ROS and MDA (Braeckman et al., 2003; Yang et al., 2021). BHBBE, CHR, and CHRG enhanced antioxidant enzyme activity (SOD and CAT), reduced ROS levels, and decreased MDA content. BHBBE, CHR, and CHRG also increased the expression of the sod-3 gene, and BHBBE increased the expression of the ctl-1 gene. Therefore, BHBBE, CHR, and CHRG up-regulated the activities of SOD and CAT by activating the sod-3 and ctl-1 genes, thereby bolstering the antioxidant defense system in C. elegans. Additionally, the expression of the sod-3 gene enhanced SOD activity and improved muscle function (Damiano et al., 2019). Therefore, BHBBE, CHR, and CHRG may enhance SOD enzyme activity and improve nematodes’ locomotion by up-regulating the sod-3 gene.
Results showed that BHBBE and CHRG increased the expression of the sek-1 gene, which is an upstream gene of the p38 MAPK pathway in C. elegans and is closely associated with oxidative stress response (Inoue et al., 2005). The transcription factor SKN-1 was related to enhance antioxidant capacity and promote longevity, and the sek-1 gene may interact with IIS pathway through the daf-16 gene (Tanaka-Hino et al., 2002; van der Hoeven et al., 2011).
Potential mechanism of BHBBE, CHR, and CHRG to prolong longevity is summarized in Figure 8. Thus, the daf-16 gene was a crucial target through the anti-aging, stress-resistant, and anti-oxidant pathways in C. elegans. Environmental stress, such as UV radiation, high-temperature, and oxidation, could reduce lifespan by phosphorylating DAF-16 protein (Davis, 2000; Oh et al., 2005). BHBBE, CHR, and CHRG extended the lifespan of nematodes subjected to heat stress and UV irradiation, presumably through the phosphorylation of DAF-16 protein in IIS pathway. Among them, CHR was the most efficient to enhance the daf-16 gene expression and to extend lifespan. BHBBE and CHRG reduced the age-1 gene expression while increasing the sir-2.1 gene expression. BHBBE, CHR, and CHRG also increased the expression of the sod-3 and ctl-1 genes, thereby enhancing SOD and CAT activity as well as anti-aging effect of C. elegans. Thus, the longevity and antioxidant effects of BHBBE and CHRG are mediated by the age-1, sir-2.1, and sek-1 genes, which regulate the translocation of DAF-16 protein.
Figure 8. The possible mechanisms of BHBBE, CHR, and CHRG against aging of C. elegans.
Conclusions
Black highland barley bran extract and its primary phenolic compounds, CHR and CHRG, significantly extended the lifespan, enhanced locomotor capacity, and improved stress resistance in C. elegans without affecting fecundity. Furthermore, the anti-aging effects of BHBBE, CHR, and CHRG were associated with the promotion of nuclear translocation of DAF-16 in IIS pathway. The activation of DAF-16 was correlated with improved antioxidant and stress phenotypes, ultimately contributing to the extension of lifespan. CHRG was most effective in up-regulating the daf-16 gene expression and extending lifespan of C. elegans. This research provides a mechanistic foundation for developing high-polyphenol cereals, such as rye, anthocyanin-rich wheat, and oats, into anti-aging functional foods.
Data Availability
The authors confirmed that the data supporting the findings of this study are available within the article and its supplementary material.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Acknowledgment
Thanks to all institutions and collaborators who provided funding and technical support for this research.
Author Contributions
Gang Liu: conceptualization, fund acquisition, and methodology; Shuyuan Fan: investigation and writing—original draft; Peng Zeng: visualization and methodology; Yunshuang He and Yuehan Shen: investigation; Changbin Lin: data curation and supervision; Chen Xia: fund acquisition and project administration; Peijun Li: formal analysis; and Xueli Li: fund acquisition and writing—review and editing.
Conflict of Interest
The authors declared that they had no conflict of interest.
Funding
This study was financially supported by Science and Technology Department of Tibet (XZ202401ZY0002), the Solid-State Fermentation Resource Utilization Key Laboratory of Sichuan Province (2021GTJC04 & 2022GTZD02), Sichuan Authentic Traditional Chinese Medicine Innovation Team Project (SCCXTD202019), the Meat Processing Key Laboratory of Sichuan Province (23-R-01), the Sichuan Province Science and Technology Support Program (2024YFFK0429), the Cuisine Science Key Laboratory of Sichuan Province (PRKX2024Z06), and the 1+9 Unveiling of the Marshal Project of Sichuan Academy of Agricultural Sciences (1+9KJGG007).
REFERENCES
Baxi, K., Ghavidel, A., Waddell, B., Harkness, T.A. and de Carvalho C.E. 2017. Regulation of lysosomal function by the DAF-16 forkhead transcription factor couples reproduction to aging in Caenorhabditis elegans. Genetics 207: 83–101. 10.1534/genetics.117.204222
Blandino, M., Locatelli, M., Gazzola, A., Coïsson, J.D., Giacosa, S., Travaglia, F., Bordiga, M., Reyneri, A., Rolle, L., and Arlorio M. 2015. Hull-less barley pearling fractions: nutritional properties and their effect on the functional and technological quality in bread-making. Journal of Cereal Science 65: 48–56. 10.1016/j.jcs.2015.06.004
Braeckman, B.P., Houthoofd, K., and Vanfleteren, J.R. 2003. Energy metabolism, anti-oxidant defense and aging in Caenorhabditis elegans. In: Model Systems in Aging, 99–144. 10.1007/978-3-540-37005-5_5
Brintz, C. 2020. Free-radical theory of aging. In: Gellman, Marc D. (ed.) Encyclopedia of Behavioral Medicine, 899–899. 10.1007/978-3-030-39903-0_191
Brown, M.K., Evans, J.L., and Luo, Y. 2006. Beneficial effects of natural antioxidants EGCG and α-lipoic acid on life span and age-dependent behavioral declines in Caenorhabditis elegans. Pharmacology Biochemistry and Behavior 85: 620–628. 10.1016/j.pbb.2006.10.017
da Costa, J.P., Vitorino, R., Silva, G.M., Vogel, C., Duarte, A.C., and Rocha-Santos, T. 2016. A synopsis on aging—Theories, mechanisms and future prospects. Ageing Research Reviews 29: 90–112. 10.1016/j.arr.2016.06.005
Damiano, S., Muscariello, E., La Rosa, G., Di Maro, M., Mondola, P. and Santillo M. 2019. Dual role of reactive oxygen species in muscle function: can antioxidant dietary supplements counteract age-related sarcopenia? International Journal of Molecular Science 20: 3815. 10.3390/ijms20153815
Davis R.J. 2000. Signal transduction by the JNK group of MAP kinases. Cell 103: 239–252. 10.1016/s0092-8674(00)00116-1
Demirtas, I., Erenler, R., Elmastas, M., and Goktasoglu, A. 2013. Studies on the antioxidant potential of flavones of Allium vineale isolated from its water-soluble fraction. Food Chemistry 136: 34–40. 10.1016/j.foodchem.2012.07.086
Deng, J.L., Xiang, Z.Y., Lin, C.B., Zhu, Y.Q., Yang, K.J., Liu, T.H., Xia, C., Chen, J., Zhang, W.H., Zhang, Y.H., and Zhu, B.Y. 2021. Identification and quantification of free, esterified, and insoluble-bound phenolics in grains of hulless barley varieties and their antioxidant activities. Food Science and Technology (LWT) 151: 112001. 10.1016/j.lwt.2021.112001
Deng, J.L., Yu, M.Y., Yang, Y.H., Liu, T.H., Xiang, Z.Y., Chen, J., Yang, K.J., Zhan, R.L., Zhu, B.Y., Zhu, Y.Q., Yang, X., Wan, Y. and Xia C. 2024. Studies of phytochemical constituents by UPLC-QTOF-MS/MS of black hulless barley bran and its antioxidation and α-glucosidase inhibition effect. Arabian Journal of Chemistry 17: 105644. 10.1016/j.arabjc.2024.105644
Ding, A.J., Zheng, S.Q., Huang, X.B., Xing, T.K., Wu, G.S., Sun, H.Y., Qi, S.H., and Luo, H.R. 2017. Current perspective in the discovery of anti-aging agents from natural products. Natural Products and Bioprospecting 7: 335–404. 10.1007/s13659-017-0135-9
Di Rosa, G., Brunetti, G., Scuto, M., Trovato Salinaro, A., Calabrese, E.J., Crea, R., Schmitz-Linneweber, C., Calabrese, V., and Saul N. 2020. Healthspan enhancement by olive polyphenols in C.-elegans wild type and Parkinson’s models. International Journal of Molecular Sciences 21: 3893–3915. 10.3390/ijms21113893
Duangjan, C., Rangsinth, P., Gu, X.J., Zhang, S.X., Wink, M., and Tencomnao, T. 2019. Glochidion zeylanicum leaf extracts exhibit lifespan extending and oxidative stress resistance properties in Caenorhabditis elegans via DAF-16/FoxO and SKN-1/Nrf-2 signaling pathways. Phytomedicine 64: 153061. 10.1016/j.phymed.2019.153061
Geng, L., Li, M., Zhang, G., and Ye, L. 2022. Barley: a potential cereal for producing healthy and functional foods. Food Quality and Safety 6: fyac012. 10.1093/fqsafe/fyac012
Guo, T., Horvath, C., Chen, L., Chen, J., and Zheng, B. 2020. Understanding the nutrient composition and nutritional functions of highland barley (Qingke): a review. Trends in Food Science & Technology 103: 109–117. 10.1016/j.tifs.2020.07.011
Herndon, L.A., Schmeissner, P.J., Dudaronek, J.M., Brown, P.A., Listner, K.M., Sakano, Y., Paupard, M.C., Hall, D.H., and Driscoll, M. 2002. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419: 808–814. 10.1038/nature01135
Higgins-Chen, A.T., Thrush, K.L., and Levine, M.E. 2021. Aging biomarkers and the brain. Seminars in Cell & Developmental Biology 116: 180–193. 10.1016/j.semcdb.2021.01.003
How, C.M., Lin, T.-A., and Liao, V.H.-C. 2021. Early-life chronic di(2-ethylhexyl)phthalate exposure worsens age-related long-term associative memory decline associated with insulin/IGF-1 signaling and CRH-1/CREB in Caenorhabditis elegans. Journal of Hazardous Materials 417: 126044. 10.1016/j.jhazmat.2021.126044
Hsu, A.-L., Feng, Z.Y., Hsieh, M.Y., and Xu, X.Z.S. 2009. Identification by machine vision of the rate of motor activity decline as a lifespan predictor in C. elegans. Neurobiology of Aging 30: 1498–1503. 10.1016/j.neurobiolaging.2007.12.007
Inoue, H., Hisamoto, N., An, J.H., Oliveira, R.P., Nishida, E., Blackwell, T.K., and Matsumoto, K. 2005. The C. elegans p38 MAPK pathway regulates nuclear localization of the-transcription factor SKN-1 in oxidative stress response. Genes Development 19: 2278–2283. 10.1101/gad.1324805
Jomova, K., Raptova, R., Alomar, S.Y., Alwasel, S.H., Nepovimova, E., Kuca, K., and Valko M. 2023. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Archives of Toxicology 97: 2499–2574. 10.1007/s00204-023-03562-9
Ke, T., Antunes Soares, F.A., Santamaría, A., Bowman, A.B., Skalny, A.V., and Aschner, M. 2020. N,N’ bis-(2-mercaptoethyl) isophthalamide induces developmental delay in Caenorhabditis elegans by promoting DAF-16 nuclear localization. Toxicology Reports 7: 930–937. 10.1016/j.toxrep.2020.07.012
Kobayashi, Y., Furukawa-Hibi, Y., Chen, C., Horio, Y., Isobe, K., Ikeda, K., and Motoyama, N. 2005. SIRT1 is critical regulator of FOXO-mediated transcription in response to oxidative stress. International Journal of Molecular Medicine 16: 237–243. 10.3892/ijmm.16.2.237
Li, X., Du, Y., Tu, Z., Zhang, C., and Wang, L. 2022. Highland barley improves lipid metabolism, liver injury, antioxidant capacities and liver functions in high-fat/cholesterol diet mice based on gut microbiota and LC-MS metabonomics. Food Bioscience 50: 102094–102094. 10.1016/j.fbio.2022.102094
Li, G., Gong, J., Liu, J., Liu, J., Li, H., Hsu, A.-L., Liu, J., and Xu, X.Z.S. 2019. Genetic and pharmacological interventions in the aging motor nervous system slow motor aging and extend life span in C. elegans. Science Advances 5: eaau5041–eaau5053. 10.1126/sciadv.aau5041
Lukinac, J., and Jukić, M. 2022. Barley in the production of cereal-based products. Plants 11: 3519–3526. 10.3390/plants11243519
Maglioni, S., and Ventura, N. 2016. C. elegans as a model organism for human mitochondrial associated disorders. Mitochondrion 30: 117–125. 10.1016/j.mito.2016.02.003
Meints, B., Vallejos, C., and Hayes, P. 2021. Multi-use naked barley: a new frontier. Journal of Cereal Science 102: 103370. 10.1016/j.jcs.2021.103370
Mishra, B., Priyadarsini, K.I., Kumar, M.S., Unnikrishnan, M.K., and Mohan, H. 2003. Effect of O-glycosilation on the antioxidant activity and free radical reactions of a plant flavonoid, chrysoeriol. Bioorganic & Medicinal Chemistry 11: 2677–2685. 10.1016/S0968-0896(03)00232-3
Morley, J.F., Brignull, H.R., Weyers, J.J., and Morimoto, R.I. 2002. The threshold for polyglutamine-expansion protein aggregation and cellular toxicity is dynamic and influenced by aging in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America 99: 10417–10422. 10.1073/pnas.152161099
Motta, H., Roos, D., Tabarelli, G., Rodrigues, O., Avila, D., and Quines, C. 2020. Activation of SOD-3 is involved in the antioxidant effect of a new class of β-aryl-chalcogenium azide compounds in Caenorhabditis elegans. Anais da Academia Brasileira de Ciências 92: e20181147. 10.1590/0001-3765202020181147
Obadi, M., Sun, J., and Xu, B. 2021. Highland barley: chemical composition, bioactive compounds, health effects, and applications. Food Research International 140: 110065. 10.1016/j.foodres.2020.110065
Oh, S.W., Mukhopadhyay, A., Svrzikapa, N., Jiang, F., Davis, R.J. and Tissenbaum, H.A. 2005. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proceedings of the National Academy of Sciences of the United States of America 102: 4494–4499. 10.1073/pnas.0500749102
Okoro, N.O., Odiba, A.S., Osadebe, P.O., Omeje, E.O., Liao, G., Fang, W., Jin, C., and Wang, B. 2021. Bioactive phytochemicals with anti-aging and lifespan extending potentials in Caenorhabditis elegans. Molecules 26(23): 7323–7323. 10.3390/molecules26237323
Park, S., Kim, B.-K., and Park, S.-K. 2021. Supplementation with phosphatidylethanolamine confers anti-oxidant and anti-aging effects via hormesis and reduced insulin/IGF-1-like signaling in C. elegans. Mechanisms of Ageing and Development 197: 111498. 10.1016/j.mad.2021.111498
Rodrigues, L.P., Teixeira, V.R., Alencar-Silva, T., Simonassi-Paiva, B., Pereira, R.W., Pogue, R., and Carvalho, J.L. 2021. Hallmarks of aging and immunosenescence: connecting the dots. Cytokine & Growth Factor Reviews 59: 9–21. 10.1016/j.cytogfr.2021.01.006
Salmon, A.B., Richardson, A., and Pérez, V.I. 2010. Update on the oxidative stress theory of aging: does oxidative stress play a role in aging or healthy aging? Free Radical Biology and Medicine 48: 642–655. 10.1016/j.freeradbiomed.2009.12.015
Son, L.T., Ko, K.-M., Cho, J.H., Singaravelu, G., Chatterjee, I., Choi, T.-W., Song, H.-O., Yu, J.-R., Park, B.-J., Lee, S.-K., and Ahnn, J. 2011. DHS-21, a dicarbonyl/l-xylulose reductase (DCXR) ortholog, regulates longevity and reproduction in Caenorhabditis elegans. FEBS Letters 585: 1310–1316. 10.1016/j.febslet.2011.03.062
Song, Y.M., Liu, D.Y., Xie, J.Y., Xie, J.H., Chen, Y., Chen, X.Y., Hu, X.B., and Yu, Q. 2024. Protective effects of EGCG on-acrolein-induced Caenorhabditis elegans and its mechanism of life extension. Food & Function 15: 5855–5867. 10.1039/d3fo05394f
Sun, X.J., Chen, W.D., and Wang, Y.D. 2017. DAF-16/FOXO transcription factor in aging and longevity. Frontiers in Pharmacology 8: 548. 10.3389/fphar.2017.00548
Takasawa, R., Takahashi, S., Saeki, K., Sunaga, S., Yoshimori, A., and Tanuma, S.-i. 2008. Structure–activity relationship of human GLO I inhibitory natural flavonoids and their growth inhibitory effects. Bioorganic & Medicinal Chemistry 16: 3969–3975. 10.1016/j.bmc.2008.01.031
Tanaka-Hino, M., Sagasti, A., Hisamoto, N., Kawasaki, M., Nakano, S., Ninomiya-Tsuji, J., Bargmann, C.I., and Matsumoto, K. 2002. SEK-1 MAPKK mediates Ca2+ signaling to determine neuronal asymmetric development in Caenorhabditis elegans. Embo Reports 3: 56–62. 10.1093/embo-reports/kvf001
van der Hoeven, R., McCallum, K.C., Cruz, M.R., and Garsin, D.A. 2011. Ce-Duox1/BLI-3 generated reactive oxygen species trigger protective SKN-1 activity via p38 MAPK signaling during infection in C. elegans. PLOS Pathogens 7: e1002453. 10.1371/journal.ppat.1002453
Wang, H., Li, T., Chen, L., Zheng, B.S., and Liu, R.H. 2020. Effects of orange extracts on longevity, healthspan, and stress resistance in Caenorhabditis elegans. Molecules 25: 351–368. 10.3390/molecules25020351
Wang, H.L., Liu, J., Li, T., and Liu, R.H. 2018. Blueberry extract promotes longevity and stress tolerance via DAF-16 in Caenorhabditis elegans. Food & Function 9: 5273–5282. 10.1039/c8fo01680a
Wang, X., Yang, K., Liu, T., Xie, S., Wang, W., Zhao, J., Liu, G., and Xia, C. 2023. Structural analysis of functional factors and antioxidant function of blackbarley bran. Food Science and Technology 48: 142–148. 10.13684/j.cnki.spkj.2023.07.021
Wu, H., Zhao, Y., Guo, Y., Xu, L., and Zhao, B.L. 2012. Significant longevity-extending effects of a tetrapeptide from maize on Caenorhabditis elegans under stress. Food Chemistry 130: 254–260. 10.1016/j.foodchem.2011.07.027
Xiang, Z.Y., Deng, J.L., Yang, K.J., Zhu, Y.Q., Xia, C., Chen, J., and Liu, T.H. 2021. Effect of processing on the release of phenolic compounds and antioxidant activity during in vitro digestion of hulless barley. Arabian Journal of Chemistry 14: 103447. 10.1016/j.arabjc.2021.103447
Xiao, Y., Zhang, Y., Li, L., Jiang, N., Yu, C., Li, S., Zhu, X., Liu, F., and Liu, Y. 2024. Cynaroside extends lifespan and improves the neurondegeneration diseases via insulin/IGF-1 signaling pathway in Caenorhabditis elegans. Archives of Gerontology and Geriatrics 122: 105377. 10.1016/j.archger.2024.105377
Yang, Y., Fan, B., Mu, Y.W., Li, Y., Tong, L.T., Wang, L.L., Liu, L.Y., Li, M.M., Sun, P.P., Sun, J., and Wang, F.Z. 2023. A comparative metabolomics study of polyphenols in highland barley (Hordeum vulgare L.) grains with different colors. Food Research International 174: 113672. 10.1016/j.foodres.2023.113672
Yang, L., Ye, Q., Zhang, X.G., Li, K., Liang, X.S., Wang, M., Shi, L.R., Luo, S.H., Zhang, Q., and Zhang, X.M. 2021. Pyrroloquinoline quinone extends Caenorhabditis elegans’ longevity through the insulin/IGF1 signaling pathway-mediated activation of autophagy. Food Function 12: 11319–11330. 10.1039/d1fo02128a
Zečić, A., and Braeckman, B.P. 2020. DAF-16/FoxO in Caenorhabditis elegans and its role in metabolic remodeling. Cells 9: 109–125. 10.3390/cells9010109
Zhang, L., Cui, D., Ma, X., Han, B., and Han, L. 2023. Comparative analysis of rice reveals insights into the mechanism of colored rice via widely targeted metabolomics. Food Chemistry 399: 133926. 10.1016/j.foodchem.2022.133926
Zhang, S.W., Li, F., Zhou, T., Wang, G.X., and Li, Z. 2020. Caenorhabditis elegans as a useful model for studying aging mutations. Frontiers in Endocrinology (Lausanne) 11: 554994–555003. 10.3389/fendo.2020.554994
Zhang, Y.Z., Yin, L.Q., Huang, L., Tekliye, M., Xia, X.D., Li, J.Z., and Dong, M.S. 2021. Composition, antioxidant activity, and neuroprotective effects of anthocyanin-rich extract from purple highland barley bran and its promotion on autophagy. Food Chemistry 339: 127849–127860. 10.1016/j.foodchem.2020.127849
Zhu, Y., Li, T., Fu, X., Abbasi, A.M., Zheng, B., and Liu, R.H. 2015. Phenolics content, antioxidant and antiproliferative activities of dehulled highland barley (Hordeum vulgare L.). Journal of Functional Foods 19: 439–450. 10.1016/j.jff.2015.09.053
Ziv, E., and Hu, D. 2011. Genetic variation in insulin/IGF-1 signaling pathways and longevity. Ageing Research Reviews 10: 201–204. 10.1016/j.arr.2010.09.002
Supplementary
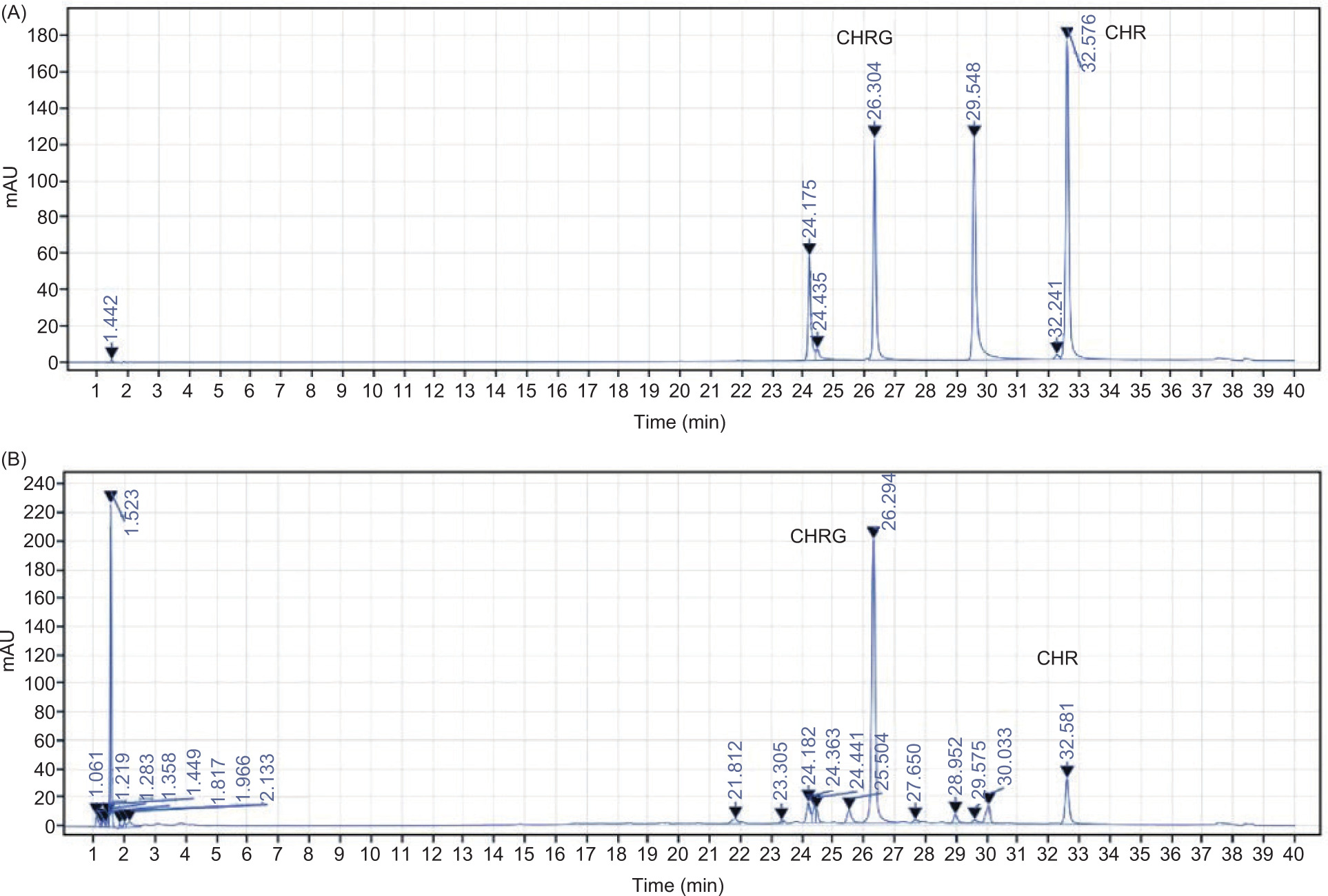
Figure S1. Purification chromatogram of CHR and CHRG from BHBBE by preparative HPLC. (A) HPLC profile of standard references: nordihydroguaiaretic acid, lignin, CHR, and CHRG. (B) HPLC chromatogram showing the isolated CHR and CHRG fractions from the BHBBE sample.
Figure S2. (A) 1H-NMR and (B) 13C-NMR spectra for the structural identification of CHR and CHRG.