
Download
REVIEW ARTICLE
Protein conjugates and probiotic interactions of dairy products for innovative and future functional foods
Nareekan Chaiwong1,2, Auengploy Chailangka3, Idris Kaida Zubairu1, Tri Indrarini Wirjantoro1, Pavalee Chompoorat Tridtitanakiat1, Juan Manuel Castagnini4, Mohsen Gavahian5, Mahsa Majzoobi6, Yuthana Phimolsiripol1,2*
1Faculty of Agro-Industry, Chiang Mai University, Chiang Mai, Thailand;
2Center of Excellence in Agro Bio–Circular–Green Industry, Chiang Mai University, Chiang Mai, Thailand;
3Livestock Products Division, Department of Livestock Development, Bangkok, Thailand;
4Research group in Innovative Technologies for Sustainable Food (ALISOST), Department of Preventive Medicine and Public Health, Food Science, Toxicology and Forensic Medicine, Faculty of Pharmacy, Universitat de València, Avda. Vicent Andrés Estellés, Burjassot, València, Spain;
5Department of Food Science, National Pingtung University of Science and Technology, Pingtung, Taiwan, ROC;
6Department of Food Science, RMIT University, Bundoora West, Melbourne, Australia
Abstract
This review critically assesses the strategies and mechanisms underlying interactions between protein conjugates and probiotics in dairy foods, aiming to detail how these conjugates stabilize probiotics, facilitate gut colonization, and stimulate immunomodulation. Protein–based carriers and complexes play key roles in improving probiotic protection, adhesion, and metabolic activity. Notable strategies include developing milk proteins (e.g., casein and whey proteins), plant–derived protein conjugates, and advanced encapsulation techniques for polysaccharide–protein complexes. The protein shell in conjugates protects probiotics from harsh conditions, enables targeted intestinal release, enhances mucosal adhesion, and contributes to enhanced antioxidant and anti–inflammatory effects. Such strategies also can improve probiotic survival and colonization and reduce inflammation (with increased IL–10 and decreased tumor necrosis factor–alpha), enhance mucus secretion (with a rise in mucin 2 [MUC2]), and elevate beneficial gut microbes. Advancements in probiotic research have enabled more precise and targeted applications using protein conjugates and encapsulation systems. These approaches form part of integrative nutrition strategies that deliver precision, protection, and personalization in the design of functional foods, contributing to food nutrition improvement. Moreover, protein conjugates are increasingly recognized as active modulators of probiotic function and immune signaling, rather than passive carriers, thereby offering new avenues for food production.
Key words: protein conjugates, probiotics, dairy foods, functional foods, encapsulation, Maillard reaction
*Corresponding Author: Yuthana Phimolsiripol, Faculty of Agro–Industry, Chiang Mai University, Chiang Mai 50100, Thailand. Email: [email protected]
Academic Editor: Rana Muhammad Aadil, PhD: National Institute of Food Science & Technology, University of Agriculture, Faisalabad, Pakistan
Received: 2 September 2025; Accepted: 7 December 2025; Published: 11 March 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Probiotics are widely used in functional dairy products because of their health benefits and consumer acceptance (Hasnain et al., 2024; Jang et al., 2024). However, their viability is often compromised during processing, storage, and gastrointestinal (GI) transit, limiting product efficacy (D’Amico et al., 2025). Protein conjugation has emerged as a promising strategy to enhance probiotic stability and functionality, primarily through the Maillard reactions and emerging technology–assisted or chemical approaches. Such conjugates protect probiotics from environmental stress, improve intestinal adhesion, and enhance dairy product quality (Vivek et al., 2023), positioning protein–probiotic interactions as a key innovation in functional food design.
The escalating demand for functional dairy products has stimulated growing interest in integrating probiotics with protein–based technologies to enhance nutritional and health benefits. Emerging processing approaches, such as ultrasound, ohmic heating, and high–pressure treatment, have been investigated for their effects on probiotic viability and metabolic performance (Gavahian et al., 2021). In this context, dairy–derived proteins have become key functional ingredients in food, clinical, and pharmaceutical formulations owing to their exceptional reconstitution, emulsifying, and foaming properties, which surpass those of lipids and carbohydrates (Wu et al., 2021). Beyond their physicochemical roles, protein hydrolysates have demonstrated prebiotic potential by supporting probiotic proliferation (Guo et al., 2022), while hydrogel–based protein–polysaccharide complexes have been developed for targeted probiotic delivery in therapeutic applications (Zheng et al., 2023). Complementary investigations conducted by Wu et al. (2024) have provided molecular insights into plant protein–probiotic interactions and underscored the structural and compositional constraints of plant–based matrices. Despite substantial progress in probiotic delivery and functional dairy research, few studies have systematically elucidated how the chemical nature and formation parameters of protein conjugates influence the survival, metabolic activity, adhesion, and host interactions of probiotics within dairy systems. Current reviews often address these aspects in isolation; for instance, Xiang et al. (2021) examined the effects of the Maillard reaction on milk proteins without linking these effects to probiotic functionality, underscoring the need for integrative perspectives on protein conjugate–probiotic interactions.
Recent research has demonstrated that under unfavorable conditions, Maillard–type conjugates formed from milk proteins, such as whey or casein, combined with carbohydrates, such as dextran or glucose, can enhance the viability of probiotics. For instance, Guo et al. (2022) demonstrated that whey protein (WP)–dextran encapsulated conjugates enhanced the heat and acid resistance of Lactiplantibacillus plantarum 21805 in yogurt formulations. Similarly, Wu et al. (2021a) demonstrated that glycosylation of whey protein via controlled dry heating of skim milk powder significantly enhanced protein thermal stability under clean–label conditions, enabling recombined evaporated milk emulsions to withstand heating at 120°C for 30 min without loss of stability. A’yun et al. (2020) reported a great improvement in the heat stability of whey protein isolate (WPI)–lactose conjugates, which could successfully stabilize oil–in–water (O/W) emulsions against heating (80°C for 20 min). In addition, the Maillard conjugates of whey protein hydrolysates (WPH) and linear dextrins (LD; degree of polymerization [DP] 23.78–64.96) were prepared by Pan et al. (2020) by dry heating, with glycation degree increasing with LD content. Glycation altered WPH secondary structure, reduced α–helix content, and enhanced antioxidant activity. Additionally, conjugation of casein–derived hydrophobic peptides with Acacia seyal polysaccharides markedly enhanced the stability of O/W emulsions (Hou et al., 2017). Thus, the Maillard–type protein–carbohydrate conjugates, such as those formed from whey, casein, or serum albumin with sugars such as dextran or glucose, hold strong potential to enhance probiotic stability and functionality in dairy products, even under heat, acid, and GI stress.
This review summarizes recent advances in protein conjugation strategies, evaluates their effects on probiotic functionality in dairy products, and highlights their potential for developing next–generation dairy products with improved health benefits and nutritional value. The review discusses key health impacts, including modulation of the gut microbiota, enhancement of the immune system, increased bioavailability of bioactive peptides, and antioxidant properties. Finally, current technological and regulatory challenges are considered along with future perspectives for the commercial application and continued development of protein–conjugation techniques in probiotic dairy products.
Overview of Probiotics and Protein Conjugation
Probiotics in dairy foods
Probiotics, defined as live microorganisms that confer health benefits when consumed in adequate amounts, remain a key focus of modern health food research (Zavišić et al., 2023; Gao et al., 2021; Hill et al., 2014). Probiotic food sources have their historical roots in fermented dairy products, including cheese and yogurt, which were traditionally produced through spontaneous fermentation by lactic acid bacteria (LAB) (Kaur et al., 2022), and are valued for their taste, shelf life, and promotion of intestinal health (Jang et al., 2024). Élie Metchnikoff, “father of innate immunity,” linked regular consumption of fermented dairy products to improved immunity, longevity, and many biochemical processes (Lee et al., 2025). Over time, scientific knowledge about probiotics has evolved to the point that the concept has expanded beyond microorganisms derived from food to include bacteria isolated from the human gastrointestinal tract (GIT), particularly those capable of promoting health and maintaining intestinal homeostasis (Zommiti et al., 2020).
Today, probiotics are widely available in various forms, including functional foods, nutritional supplements, and pharmaceutical preparations. Owing to their advantageous physicochemical characteristics, including buffering capacity, nutritional richness, and compatibility with microbial growth and survival, dairy products remain the most widely used and effective carriers of probiotics (Latif et al., 2023). This also reflects the worldwide acceptance of dairy–based probiotics as promoting overall health, lifespan, and disease prevention (Kaur et al., 2022).
Applications derived from dairy products benefit from a wide range of probiotic species. Among these, the most commonly employed genera include Lactobacillus, Bifidobacterium, Streptococcus, Lactococcus, Enterococcus, Leuconostoc, and Propionibacterium (Vivek et al., 2023). A broad spectrum of strains, such as Lactobacillus acidophilus, Lacticaseibacillus rhamnosus, Lacticaseibacillus casei, L. plantarum, Limosilactobacillus reuteri, B. bifidum, Bifidobacterium longum, and Bifidobacterium lactis, are regularly included in yogurt, fermented milk, cheese, and milk powders (Vivek et al., 2023). Importantly, there is no consensus on the lowest effective dose of probiotics; however, a reasonable daily intake of approximately 109 colony–forming units (CFU), taken consistently, is usually considered sufficient to confer health benefits (Latif et al., 2023).
In addition to probiotics, nondigestible food ingredients, known as prebiotics, are sometimes added to enhance the effectiveness of most fermented dairy products. Probiotic species, including Bifidobacterium and Lactobacillus, break down prebiotics, such as inulin, fructooligosaccharides (FOS), and galactooligosaccharides (GOS), thereby improving gut microbiota composition and mucosal immunity (Wang et al., 2020). Consequently, in synbiotic formulations, probiotics and prebiotics exhibit synergistic effects that promote colonization, enhance microbial survival, and yield more physiological outcomes in the host (Parhi et al., 2024). A comprehensive overview of probiotic microorganisms used in various dairy products and their applications are presented in Table 1.
Table 1. Probiotic microorganisms used in dairy products and their applications.
| Probiotic strains/species | Product type | Dose | References |
|---|---|---|---|
| Lactococcus lactis subsp. cremoris, Propionibacterium freudenreichii spp. Shermanii |
Cheese | 106–108CFU/g | Kondrotiene et al., 2023; Rabah et al., 2017 |
| L. acidophilus, Bifidobacterium spp., Streptococcus thermophilus (ABT culture) |
Cream cottage cheese (fortified with bovine colostrum) | 107–108CFU/g | Abdeen et al., 2024 |
| L. acidophilus | Probiotic cream cheese (with Opuntia ficus–indica mucilage) | 107–108CFU/g | Rodrigues et al., 2024 |
| Pichia cerevisiae, Pediococcus acidilactici, Pediococcus pentosaceus | Swiss cheese | 106–107CFU/g | Doğan and Ay, 2021 |
| L. plantarum L33 | White–brined cheese (co–cultured with probiotic) | 107CFU/g | Zareie et al., 2025 |
| Limosilactobacillus fermentum (probiotic) | Infant formula with dairy lipids + probiotic | 1.9 × 108CFU/mL | Lemaire et al., 2021 |
| L. rhamnosus GG | Fermented beverage kefir (from grains or starter culture) | 106–108CFU/mL | Yousefvand et al., 2022 |
| Lactobacillus helveticus,B. longum, L. rhamnosus GG |
Probiotic–fortified kefir | 107–108CFU/mL | Noori et al., 2025 |
| Saccharomyces cerevisiae,Candida utilis (Candida torulopsis) | Fermented milk | 107–108CFU/mL | Benedetti et al., 2024 |
| Lacticaseibacillus paracaseiJY025 | Probiotic–fortified milk powder | 106–107CFU/mL | Shen et al., 2024 |
| L. casei 431, L. acidophilus LA–5, L. paracasei L–26, L. rhamnosus,Bifidobacterium animalis subsp. lactis BB–12 | Sheep–milk symbiotic probiotic ice cream | 108CFU/g | Kowalczyk et al., 2022 |
| L. paracasei (Shirota), L. lactis subsp. lactis YIT 2027 |
Shirota–style fermented milk drink | 107–108CFU/g | Oana et al., 2023 |
| L. acidophilus,L. plantarum,B. lactis | Buffalo milk yogurt | 106–108CFU/g | Akbal et al., 2025 |
| L. plantarum,L. casei, L. rhamnosus,S. thermophilus, Lactobacillus delbrueckii spp. Lactis | Cashew/camel milk labneh (concentrated yogurt) | 107–109CFU/mL | Al–Sulbi and Shori, 2022 |
| B. bifidum,B. longum, Bifidobacterium breve,B. lactis, Bifidobacterium adolescentis |
Yogurt | 107CFU/mL | Sibanda et al., 2024; Talearngkul et al., 2023 |
| L. paracasei | Yogurt with added probiotic strains | 107–108CFU/g | Moiseenko et al., 2023 |
| S. thermophilus, | Yogurt, cheese | Huang et al., 2024 | |
| L. acidophilus,L. casei, L. rhamnosus,L. delbrueckii,L. plantarum,L. fermentum, Streptococcus intermedius, Lactobacillus salivarius, and Lactobacillus brevis |
Yogurt, fermented milk | 107–109CFU/mL | Dimitrellou et al., 2025 |
Notes: ABT: a mixed starter culture for fermented foods, combining Lactobacillus acidophilus, Bifidobacterium spp., and Streptococcus thermophilus culture; CFU: colony–forming unit.
Dairy products are among the most established and successful vehicles for probiotics in the food industry because of their inherent nutrient content, favorable physicochemical properties, and consumer acceptance (Jang et al., 2024). In particular, LAB play a vital role in ensuring microbiological safety, shelf life, and sensory quality of fermented dairy products by synthesizing organic acids and antimicrobial peptides and competing with pathogens through competitive inhibition (Ibrahim et al., 2021). Milk provides an ideal medium for delivering viable probiotic cultures because of its buffering capacity, nutritional density, and protein structure, which protect probiotics during GI transit (Gao et al., 2022). Among fermented dairy products, yogurt is the most commonly consumed and investigated vehicle for probiotic bacteria. Moreover, the sector has recently expanded to include probiotic–enriched fermented dairy products, such as pasteurized milk, ice cream, and infant formula, which use microencapsulation or post–fermentation addition techniques to ensure probiotic viability during storage (Sbehat et al., 2022).
Various studies have demonstrated that probiotic dairy products can regulate gut bacteria, improve lactose digestion, enhance the immune system, and alleviate manifestations of GI diseases (Mudgil et al. 2024a). The combinations of probiotics in both fermented and nonfermented dairy matrices reflect growing innovation in the design of functional dairy foods. These products are developed for their nutritional value and targeted health benefits, including relief from lactose intolerance, immune modulation, and gut microbiota balance. However, maintaining probiotic viability throughout processing, distribution, and storage remains a key challenge, particularly for nonfermented products, such as ice cream and infant formula. Therefore, to fully harness the health benefits of probiotic dairy products, especially nonfermented types, it is essential to enhance the stability of probiotics across processing and storage.
Protein–probiotic conjugation mechanisms
Protein conjugates interact with probiotics at the molecular level to enhance gastrointestinal stability, adhesion, and colonization via multiple biochemical and structural mechanisms. These strategies enable targeted delivery (Liu et al., 2025), enhance stress tolerance (Zhang et al., 2023a), and improve therapeutic efficacy in the gut environment (Peled et al., 2024; Yin et al., 2025). Table 2 presents key protein–probiotic conjugation mechanisms.
Table 2. Important mechanisms for protein–probiotic conjugation.
| Strategy | Mechanism | Functional outcome | Trial application | References |
|---|---|---|---|---|
| Bioorthogonal reactions | DBCO–modified probiotics react with azido–decorated D–alanine in gut bacteria | Enhanced adhesion, prolonged gut retention | Improved colonization, colitis relief | Song et al., 2022 |
| Maillard–conjugates | Core–shell conjugates of 2'–fucosyllactose and lactoferrin hydrolysates | Increased probiotic growth, SCFA production, and reduced inflammation | Enhanced gut health | Peled et al., 2024 |
| Surface protein interactions | LPxTG motif proteins bind phenols and oligosaccharides | Improved stress resistance and GI tolerance | Stronger gut colonization | Zhang et al., 2022a |
| Probiotic–drug conjugates | Surface–modified probiotics deliver drugs site–specifically | Synchronized colonization and localized therapy | Ulcerative colitis treatment | Yin et al.,2025 |
| Multicellular microcolonies | Encapsulation into acid–resistant, stress–relaxing colonies | Significantly enhanced gut survival and colonization | Clinical probiotic delivery | Liu et al., 2025 |
Notes: SCFA: short–chain fatty acids; DBCO: dibenzocyclooctyne; LPxTG: Leu–Pro–any–Thr–Gly.
The initial improvements in probiotic survival, adhesion, and localization are primarily driven by direct protein–probiotic interactions, whereas the major therapeutic outcomes, such as production of short–chain fatty acids (SCFA), immune modulation, and disease relief, are largely mediated by secondary microbiome–driven effects (Liu et al., 2025; Peled et al., 2024). In practice, full efficacy results from synergistic coupling of both mechanisms, rather than from either pathway alone. Furthermore, despite the potentiality of these mechanisms in the protein–probiotic interactions, they approach challenges related to strain specificity, environmental variability within the gut, and long–term microbiome interactions (Song et al., 2022; Zhang et al., 2023a), thus paving the way for further study, such as validation through in vivo to show feasibility for clinical translation.
Protein conjugation techniques used in dairy systems
Protein conjugates refer to covalently bonded complexes formed between proteins and other biomolecules, such as carbohydrates, polyphenols, or similar compounds, through various chemical or enzymatic processes, enhancing the functional properties of the resulting complexes. Structurally and functionally, protein–sugar conjugates can shield probiotics from environmental stresses, increase their adherence to the intestinal epithelium, and even improve the sensory quality of the final dairy product (Vivek et al., 2023; Zhang et al., 2023b). Conjugation can alter the physicochemical and biological characteristics of milk proteins. The growing demand for functional dairy products has therefore driven substantial advances in protein modification technologies. Among the various techniques available, studies have identified three main approaches: the Maillard–type conjugation, enzymatic cross–linking, and physical activation, which currently hold the greatest potential. The methods and parameters used in recent studies on protein–carbohydrate conjugates in dairy products are presented in Table 3.
Table 3. Methods of protein–carbohydrate conjugations in dairy–based probiotics.
| Conjugation | Method | Dairy products | Probiotic strains | Properties | References |
|---|---|---|---|---|---|
| WPI–Dextran | Dry heating at 80°C for 5 h, 79% RH | Milk beverage | L. plantarum | • Storage at 4°C for 90 days with a reduction of 0.33 log CFU/mL • GI survival improved, and heat resistance demonstrated at 65–85°C |
Guo et al., 2022 |
| WPI–XOS | Dry heating 65°C, 79% RH, 24–48 h | Yogurt | L. rhamnosus | • Spray drying survival 99.83 ± 8.44% • Storage reduction is less than 1 log CFU/g over 10 weeks at 4°C • GI digestion reduction 0.20 log CFU/g after 4 h • Final count 4.09 × 109 CFU/g |
Liao et al., 2021 |
| Gelatin–XOS | Dry heating 65°C, 79% RH | Spray–dried capsule | L. fermentum | • Increased GI survival and heat resistance • Maximum survivability of 13.91% observed at 60–min MRP |
Li et al., 2023 |
| SPI–Xylose | Maillard 70°C, 65% RH, 24 h | Spray–dried powder | L. acidophilus | • Spray drying survival 21.36–25.31% for MRP–based samples, compared to 20.17% for the mixture | Du et al., 2023 |
| Milk exosome–protein conjugates | Low–temp exosome adsorption (4°C), pH 6.8 | Functional milk | Akkermansia muciniphila | • Initial viable count 1 × 108 CFU/mL • Encapsulation efficiency 90.37 ± 0.45% • In vitroGI survival rate 94.53 ± 0.26% • Nontoxic to cells and promoted cell health |
Hao et al., 2025 |
| Protein–oligosaccharide | Shell–cross–linked conjugation | Functional delivery system | Mixed LAB strains | • Targeted colon delivery and improved bioactive delivery • Growth of L. caseiincreased two–fold • Peptide fraction above 5 kDa 64.4–82.9% compared to 12% for native |
Peled and Livney, 2021 |
| WP–carboxymethylated dextran + sugar (trehalose/lactose/sucrose) | 50–60°C, RH–controlled oven, 24–48 h | Yogurt matrix | L. plantarum LP90 | • Freeze–drying survival trehalose 97.8% and lactose 98.5% • GI tolerance greater than 5 h |
Li et al., 2025 |
Note: WPI: whey protein isolate; XOS: xylooligosaccharide; SPI: soy protein isolate; GI: gastrointestinal; LAB: lactic acid bacteria; CFU: colony forming unit; MRP: Maillard reaction products.
The Maillard, enzymatic, and chemical conjugation each provides distinct mechanistic avenues for improving probiotic functionality in dairy systems, although their benefits and limitations diverge. The Maillard conjugation, driven by the covalent attachment of reducing sugars to lysine residues, reinforces protein matrix stability while simultaneously enhancing the protective barrier against acid and bile stress (Gao et al., 2021; Zhang et al., 2023b). By contrast, enzymatic conjugation, most notably via microbial transglutaminase (MTGase), exhibits greater substrate specificity, producing ε–(γ–glutamyl)–lysine cross–links that not only stabilize probiotic encapsulation but also enhance mucosal adhesion and promote the controlled release of bioactive peptides during digestion (Milczek, 2018). Chemical conjugation expands versatility by enabling the covalent grafting of proteins with polyphenols or polysaccharides to form complexes that provide strong shielding against environmental stressors; however, concerns over off–flavors, altered digestibility, and regulatory approval limitations restrict their translational potential (Zhang et al., 2023a).
Polysaccharide–protein complexes (PPCs) offer significant advantages over single–protein carriers in dairy systems, serving as biocompatible alternatives to synthetic emulsifiers (Sun et al., 2022). They enhance colloidal stability during processing and storage, improve the encapsulation and bioaccessibility of bioactive compounds, and positively modify textural and sensory attributes, collectively leading to greater consumer acceptance of dairy products (Liu et al., 2025). Despite these functional benefits, the industrial application of PPCs is constrained by scalability and reproducibility challenges. Protein–polysaccharide interactions are highly sensitive to processing conditions, such as pH and ionic strength, often resulting in variability in complex formation and performance (Goh et al., 2020). Furthermore, maintaining consistent quality during large–scale production and meeting clean–label and regulatory requirements remain critical hurdles that must be addressed to enable broader adoption of PPCs in commercial dairy systems (Liu et al., 2025).
Maillard–protein conjugation
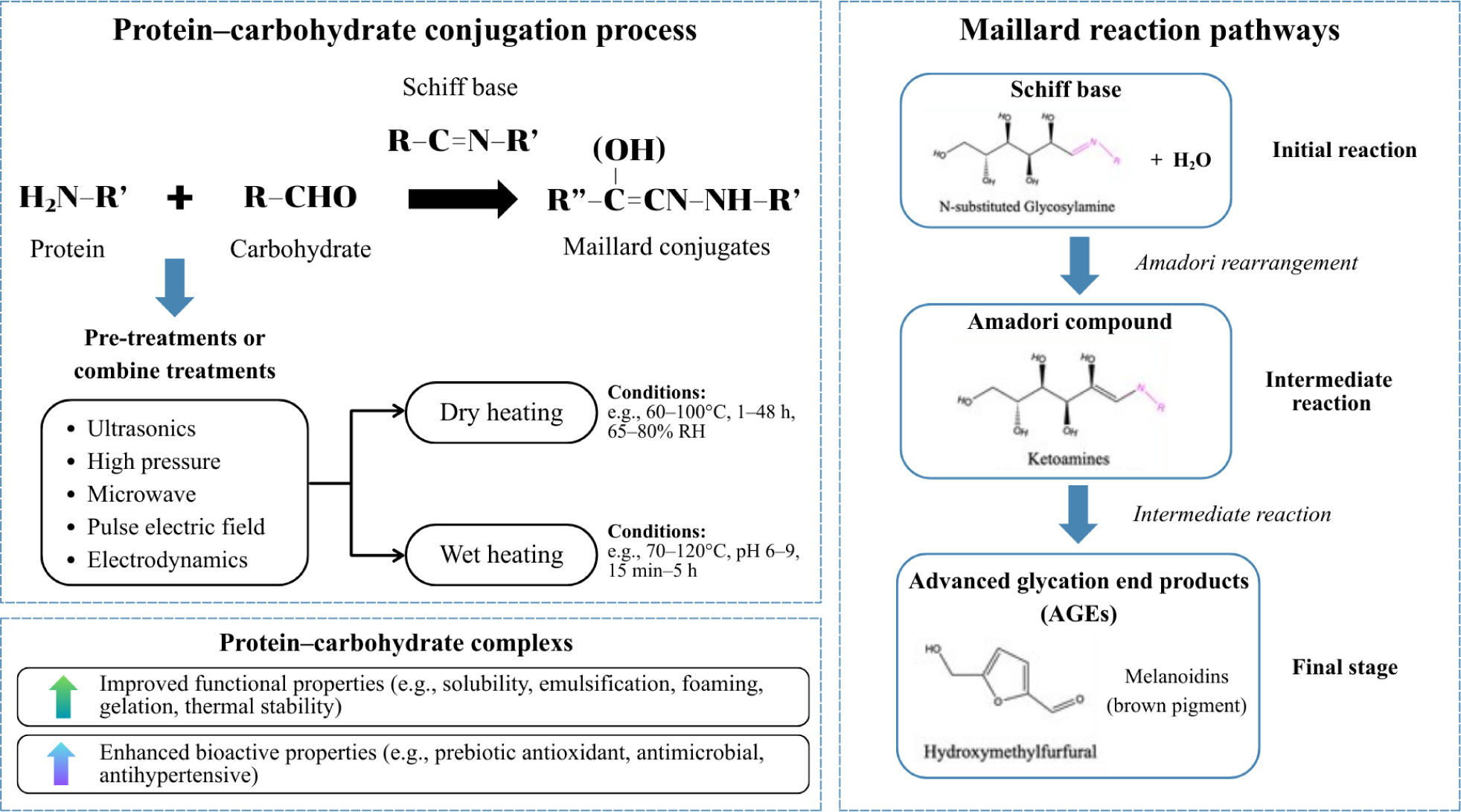
The Maillard reaction is a nonenzymatic glycation process in which reducing sugars react with protein amino groups to form Schiff bases, ultimately leading to stable ketoamine linkages (Figure 1) (Liu et al., 2020). In dairy systems, this reaction is typically performed under dry or semi–dry conditions at moderate temperatures (50–80°C) and approximately 70% relative humidity (RH) for 24–72 h (da Silva et al., 2019). The reaction alters protein structures by exposing hydrophilic groups and increasing surface activity, thereby enhancing their functional and protective roles. It has been widely applied to milk proteins, including WPI (Xu et al., 2019), casein (Yuan et al., 2024), and milk protein concentrate (Wu et al., 2021b), with saccharides, such as inulin, dextran, Arabic gum, and xylooligosaccharides (XOS)/GOS. While this processing can improve the nutritional quality and quality of foods, it requires precise control. Excessive heating may cause undesirable browning, nutritional losses of essential amino acids, and the formation of advanced glycation end products (AGEs), which are increasingly concerning for health–conscious consumers. For the same reason, Bi et al. (2023) recommended exposure at 70°C for 15 s as optimal conditions to preserve both quality and nutritional value in a study on the effect of pasteurization on the Maillard reaction in lactose–free milk.
Figure 1. Schematic illustration of protein–carbohydrate conjugation pathways. Physical pretreatments followed by dry or wet heating promote Maillard conjugation, enhancing the functional and bioactive properties of the resulting protein–carbohydrate complexes.
The Maillard conjugation substantially improves the functional properties of milk proteins, including solubility, heat stability, emulsifying ability, and antioxidant activity (Xu et al., 2019). These enhancements directly translate into improved probiotic survival during GI stress and thermal processing, making this approach particularly valuable for yogurt and fermented milk products. In such matrices, Maillard–modified proteins not only protect probiotic cells but also enhance texture and provide prebiotic benefits, supporting synbiotic formulations. Consistently, higher counts of Lactobacillus and Bifidobacterium have been reported in conjugate–fortified dairy, compared with nonconjugated carriers (Chaiwong et al., 2025; Liao et al., 2021).
Extending this strategy to plant–based proteins presents challenges because of structural heterogeneity, reduced solubility, and lower lysine availability, all of which can hinder conjugation efficiency. For example, pea protein–polydextrose conjugates produced by wet heating have been shown to improve solubility, emulsifying, and foaming properties, with the protein–to–polydextrose ratio, temperature, and time as key factors (Khan et al., 2024). Similarly, soy protein isolate–oligosaccharide Maillard conjugates enhanced the antioxidant activity of the carrier and improved Lactobacillus casei survival under simulated digestion, maintaining 7.4 log CFU/mL, compared to 1.6 log CFU/mL in free cells (Zhong et al., 2021). Moreover, Nghiep (2024) found that optimized energy–nitrogen combinations enhance nutrient value and protein efficiency, affecting feed intake and nutrient digestibility in Saanen crossbred goats, suggesting that the conjugation effect of protein–polysaccharides is of great importance in dairy systems. Despite such progress, plant–based Maillard reaction products (MRPs) still fall short of the probiotic protection typically achieved with dairy carriers, underscoring the need for further optimization of conditions and formulations.
Importantly, the Maillard conjugation utilizes food–safe materials and provides multifaceted functionality without relying on synthetic additives. It improves solubility, emulsification, antioxidant activity, and probiotic stability (Chaiwong et al., 2025). Nonetheless, uncontrolled reactions may induce browning, off–flavors, and the formation of AGEs, which pose nutritional and health concerns. When carefully managed, however, the Maillard–protein conjugation remains a powerful strategy for probiotic delivery, particularly in dairy applications (Dong et al., 2023; Kathuria et al., 2023). Moreover, recent work has highlighted the potential of two–stage complexation–covalent conjugation followed by complex coacervation as a promising route to engineer more stable protein–carbohydrate systems (Zhang et al., 2023b).
Enzymatic protein–conjugation
Enzymatic protein conjugation involves specific biocatalysts that form covalent bonds between proteins or between proteins and other biopolymers, thereby tailoring their functional and structural properties. Among these, MTGase is the most widely used enzymatic conjugation method in dairy systems. Other enzymes, such as laccases, tyrosinases, and peroxidases, catalyze protein–polyphenol or protein–protein cross–linking, potentially conferring functional benefits in plant–based systems due to their broader substrate ranges (Gouseti et al., 2023). MTGase catalyzes the formation of ε–(γ–glutamyl) lysine bonds via acyl transfer between glutamine and lysine residues (Milczek, 2018), thereby generating larger, more stable protein networks. Reactions usually occur under mild conditions (30–50°C, near–neutral pH) and are completed within 1–4 h (da Silva et al., 2019), making the process compatible with probiotic viability. In dairy matrices, MTGase is effective at low doses, is safe, and enhances the texture of yogurt and cheese by strengthening casein gels, thereby improving viscosity and gel strength, and enabling physical entrapment of probiotic cells (da Silva et al., 2019; Milczek, 2018). Furthermore, Halavach (2024) demonstrated that enzymatic hydrolysis of whey and colostrum proteins using alcalase and protozyme (5%, 50–60°C) generated low–molecular–weight peptides (<10 kDa) with high immunoglobulin hydrolysis (82.4%). These hydrolysates are potentially hypoallergenic and suitable for functional foods and nutraceutical applications. Compared to chemical cross–linking, enzymatic conjugation offers advantages in precision, safety, and the avoidance of unwanted by–products, such as AGEs (Kathuria et al., 2023).
Despite the advantages of enzymatic conjugation in dairy systems, the cost of enzymes, batch–to–batch variability in activity, and inhibition by food components (e.g., metal ions, phenolics, or extreme pH levels) may limit scalability (Kathuria et al., 2023). Additionally, although MTGase efficiently cross–links caseins owing to their flexible, open structures, plant proteins (e.g., soy, pea, and oats) pose greater challenges. Their globular conformation and reduced lysine/glutamine accessibility reduce conjugation efficiency (Chen et al., 2020), often necessitating pretreatment (e.g., denaturation, hydrolysis, or high–pressure processing) to expose reactive sites. This distinction highlights a crucial difference: applying the same approach to plant–based matrices often compromises sensory quality by producing excessive gel rigidity and off–flavors, or necessitates higher enzyme inputs, thereby increasing regulatory concerns (Sim et al., 2021). However, such effects may be more tolerable in plant–based alternatives (Santoso et al., 2024; Yi et al., 2024). Therefore, a critical balance must be achieved between functionality and sensory acceptance. Furthermore, future research should critically compare enzyme systems across both domains, tailoring approaches to optimize reactivity while preserving desirable sensory attributes.
Chemicals–protein conjugation
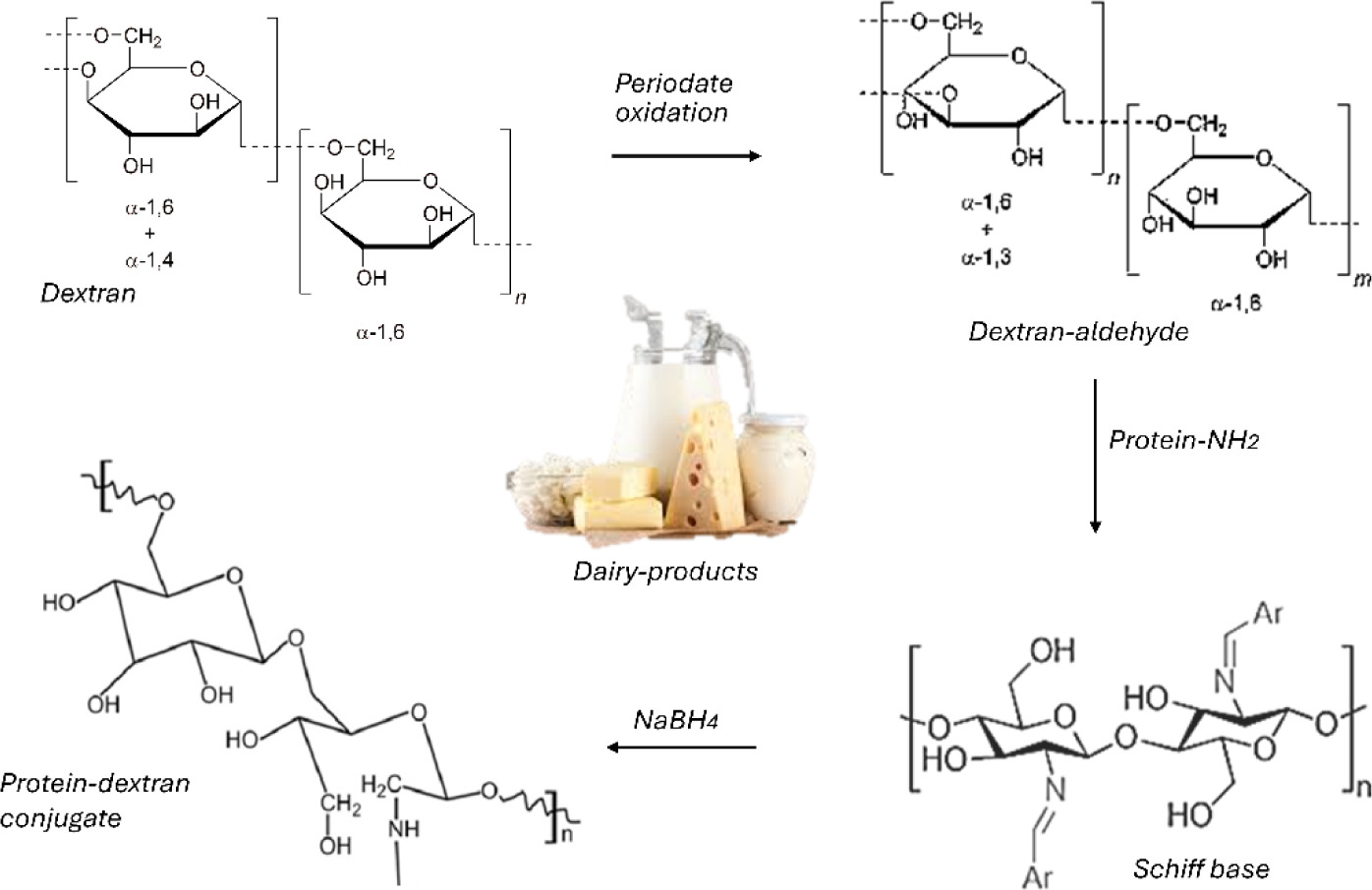
Chemical conjugation, which has limited applications in commercial dairy products due to regulatory constraints, employs external cross–linking agents to form covalent bonds between protein molecules or to graft functional groups, such as polyphenols, oligosaccharides, or polysaccharides (Figure 2), onto proteins (Zanjani et al., 2018). These reactions target nucleophilic functional groups on protein surfaces, including amines, carboxyls, thiols, and phenolic residues. Among these, common food–compatible cross–linkers include genipin (from Gardenia jasminoides), EDC (1–ethyl–3–(3–dimethylaminopropyl) carbodiimide), and oxidized polysaccharides with reactive aldehyde groups (Wei and Huang, 2019; Zanjani et al., 2018).
Figure 2. Schematic illustration of dextran–protein conjugate formation via periodate oxidation and reductive amination. Dextran is first oxidized by sodium periodate to produce aldehyde groups, which subsequently react with the amino groups of proteins to form Schiff bases. The imine linkages are then reduced by sodium borohydride (NaBH4) to yield stable covalent dextran–protein conjugates.
Chemical conjugation approaches in dairy applications are primarily limited to food–grade, safe cross–linking agents. For example, genipin–cross–linked protein matrices have been utilized to encapsulate probiotic strains, such as L. reuteri, thereby enhancing their viability during spray drying and exposure to simulated GI fluids (Laurujisawat et al., 2025). Additionally, chemical grafting of proteins to dietary fibers (e.g., pectin or alginate derivatives) or polyphenolic compounds (e.g., tannic acid and catechins) can create protein–polyphenol or protein–fiber complexes with protective, antioxidant, and antimicrobial properties within the constraints of food safety regulations (Wei and Huang, 2019).
Protein conjugation via chemical methods offers high versatility and stability, enabling the design of multifunctional delivery systems resistant to acid, heat, and mechanical stress. However, potential limitations include off–flavors, discoloration (especially with genipin), and reduced digestibility of excessively cross–linked proteins. Additionally, the regulatory status of some cross–linkers may pose challenges, as only specific agents, such as genipin, are approved for food use, and any unreacted chemicals must be removed or proven unsafe (Laurujisawat et al., 2025; Wei and Huang, 2019; Zanjani et al., 2018). Thus, chemical–protein conjugation offers strong functional benefits but is limited by regulatory restrictions, sensory changes, and the need for safe, food–grade cross–linkers.
Mechanism of interactions of protein conjugates in the dairy system
The description of the Maillard, enzymatic, and chemical conjugation techniques provides an overview of their synthesis; however, a mechanistic understanding is essential to explain how these conjugates interact with probiotic cell surfaces. At the molecular level, electrostatic attraction, hydrophobic interactions, and hydrogen bonding are primarily responsible for binding between conjugated biopolymers and bacterial membranes (Liu et al., 2023; Sun et al., 2023). The cationic amino acid residues of proteins in the conjugates can interact with the negatively charged teichoic acids, peptidoglycan, and exopolysaccharides on the probiotic cell walls, whereas the hydrophobic regions anchor to the lipid components of the membrane (Sun et al., 2022). In Maillard– and enzymatically derived conjugates, reactive carbonyl and amino groups can form weak covalent or hydrogen–bonding interactions with surface proteins or polysaccharides, thereby enhancing adhesion stability (Liu et al., 2023). Additionally, surface–layer (S–layer) proteins, lipoteichoic acids, and extracellular polysaccharides of Lactobacillus and Bifidobacterium species act as specific binding sites that mediate these interactions (Bönisch et al., 2018; Du et al., 2023). The conjugate–cell–surface associations reinforce membrane integrity, reduce permeability to environmental stressors, and enhance probiotic resilience during processing and GI transit (Sun et al., 2023).
This study confirms that the effectiveness of conjugation varies among probiotic strains because of differences in surface proteins and cell wall composition, making the response largely strain–dependent, as highlighted in Table 3. Furthermore, there is no universal ratio; most studies report effective conjugation between 1:1 and 1:3 (w/w), but the optimal proportion depends on the strain and the conjugation method used. The conjugation approaches in this review provide insight into a continuum between functional efficacy and practical applicability, suggesting that future innovation may dwell in hybrid systems that combine the biological precision of enzymatic strategies with the structural resilience of both Maillard and chemical pathways, as indicated by several studies in Table 3.
Role of Protein Conjugates on Probiotic Functionality of Dairy Products
Role of conjugation on the survival of probiotics in dairy products
Probiotics often experience viability loss under various processing conditions, including thermal treatments, spray drying, freeze drying, and prolonged storage (Manyatsi et al., 2024). However, protein conjugation has proven highly effective in addressing these challenges. Several studies have demonstrated that conjugating proteins, such as WPI, sodium caseinate, soy protein isolates, and plant–derived proteins such as peas and rice, with carbohydrates or polyphenolic compounds via various encapsulation processes enhances the proportion of probiotic survival (Xu et al., 2019). Pumpkin carotenoids complexed with casein and soy proteins showed improved stability and functionality after spray drying (Werasakulchai et al., 2025). Protein–carotenoid powders are suitable for use in food. This is consistent with protein conjugation strategies in dairy foods to enhance bioactive delivery.
For example, Liao et al. (2021) prepared the Maillard reaction conjugates of WPI and XOS to encapsulate L. rhamnosus by spray drying. Results showed that WPI–XOS conjugates improved the survival of L. rhamnosus during spray drying with enhanced storage stability (at 4°C) for up to 8 weeks. Similarly, Song et al. (2024) investigated WPI–soybean hull polysaccharide conjugates that encapsulated L. casei by freeze–drying at –80°C. The result showed that probiotics encapsulated with conjugates can survive during processing and storage. This results in higher viability during freeze–drying (87.35% vs. 24.95% for unencapsulated) and better thermal resistance during pasteurization at 60°C and 80°C. Furthermore, ultrasound–assisted multilayer Pickering emulsions prepared from WPI conjugated with (–)–epigallocatechin–3–gallate (EGCG) protected L. plantarum well during pasteurization at 63°C for 30 min. The probiotic survival proportion after pasteurization for the unconjugated sample was 48.5%, compared to 74.25% for the WPI–EGCG conjugated multilayer emulsion. This indicates that the conjugated system improved thermal protection by approximately 25.75% (He et al., 2023).
Additionally, Mudgil et al. (2024b) investigated the stability of L. reuteri DSM 17938 encapsulated within pea and rice protein conjugates with inulin. The results showed that conjugating plant proteins with prebiotic inulin and rice protein achieved the highest percentage of efficiency (96.99%), compared to pea protein conjugates (92.87%). Protein conjugates form dense protective layers around probiotics, shielding them from environmental stresses, including heat, moisture, oxygen, and acidic conditions (Li et al., 2025). Specifically, the Maillard reaction enhances barrier properties by forming stable covalent bonds between proteins and carbohydrates, resulting in robust microcapsules that prevent probiotic degradation during processing and storage (Gao et al., 2021; Liao et al., 2021; Minj and Anand, 2022).
Protection of probiotics against gastrointestinal stressors
During digestion, probiotics encounter several stressors: in the stomach, the environment is highly acidic (pH 1.5–3.0) and contains digestive enzymes, such as pepsin; in the small intestine, exposure to bile salts and pancreatin at near–neutral pH (6.5–7.5) further threatens cell viability; and in the colon, anaerobic conditions and microbial competition present additional survival hurdles (Li et al., 2025; Minj and Anand, 2022; Werasakulchai et al., 2025). To improve probiotic viability, recent research has focused on the use of protein–carbohydrate Maillard conjugates formed by covalent bonding between proteins (e.g., WPI, soy protein isolate, and caseinate) and carbohydrates (e.g., oligosaccharides and polysaccharides) as effective protective encapsulating agents (Du et al., 2023; Liao et al., 2021; Mudgil et al., 2024b). Similarly, Liao et al. (2021) reported enhanced resistance of L. rhamnosus encapsulated in WPI–XOS Maillard conjugates under simulated gastric conditions. When exposed to simulated gastric fluid (pH 2.0, for 2 h at 37°C), free cells exhibited a drastic reduction in viability, dropping to below 4.0 log CFU/g, while encapsulated cells maintained over 7.5 log CFU/g, indicating strong acid resistance and potential for delivery to the intestines.
Moreover, probiotics encapsulated within WPH–maltodextrin conjugates demonstrated improved survival under simulated gastric acid conditions (pH 2.5 for 2 h at 37°C). L. acidophilus encapsulated maintained a viability of 7.4 log CFU/g, compared with a sharp decline to 3.2 log CFU/g in the control. Furthermore, upon exposure to bile salts (0.3% for 3 h at 37°C), the encapsulated cells remained above 7.0 log CFU/g, demonstrating the conjugate’s effectiveness in maintaining probiotic viability across GI conditions (Minj and Anand, 2022). Furthermore, the hydrophilic–hydrophobic balance and viscoelastic gel structure of protein conjugates delay digestive penetration and modulate controlled release, enabling colon–targeted delivery (Gao et al., 2021; Minj and Anand, 2022). The challenge in moving forward exists in optimizing conjugate formulations for large–scale food applications while ensuring safety by avoiding the overproduction of AGEs.
Protein conjugates enhance probiotic survival under GI stress by creating a multi–layered protective matrix that reduces direct exposure of cells to gastric acid, bile salts, and digestive enzymes (Li et al., 2025; Werasakulchai et al., 2025). Mechanistically, conjugated proteins reduce proton permeability and bind bile salts, thereby mitigating membrane disruption in probiotic cells. Studies in simulated GI models demonstrate that whey protein–oligosaccharide conjugates increase survival proportions of Lactobacillus rhamnosus and Bifidobacterium longum, compared to unconjugated carriers (da Silva et al., 2019; Liao et al., 2021; Rolim et al., 2021). Preliminary human intervention studies further suggest that probiotic delivery with whey protein conjugates results in higher recovery of viable Lactobacillus in the small intestine, supporting improved colonization and persistence (Rolim et al., 2021). These findings suggest that protein conjugation strategies not only serve as physical shields but also modulate the digestive microenvironment, thereby creating translational opportunities for next–generation probiotic formulations in functional dairy systems.
Role of Protein Conjugates on the Quality of Dairy Products
Functional quality of dairy products with protein–carbohydrate conjugation
Researchers have demonstrated that conjugated proteins with carbohydrates, formed through the Maillard reaction, can seriously impact the technological and functional properties of the final conjugates, particularly in areas such as solubility, heat stability, emulsification, foaming properties, gelation, and texture (Deng et al., 2025; Urango et al., 2024; Zhang et al., 2023b). Primarily, conjugates made through the Maillard–type glycation or enzymatic cross–linking exhibit amphiphilic characteristics that enable them to serve as protective matrices and functional stabilizers in dairy–based systems. The key function of protein–carbohydrate conjugates in probiotic dairy systems is their ability to act as encapsulating agents. Protein conjugates can form protective matrices around probiotics through microencapsulation processes (Loyeau et al., 2021; Werasakulchai et al., 2025).
Additionally, several studies have demonstrated diverse applications of protein conjugates in dairy products. Mudgil et al. (2025) showed that camel milk protein complexed with polyphenols from dates’ seeds achieved substantially improved emulsification stability and nearly three–fold increase in foaming capacity compared to nonconjugated protein powders. Chaiwong et al. (2025) reported that whey protein–GOS conjugates (1:2 ratio, heated for 6 h at 90°C) exhibited superior solubility and emulsion stability compared to whey protein alone, demonstrating better technological properties than other conjugate ratios tested. Additionally, Akalan et al. (2024) added 1% protein conjugate prepared from brown rice protein and microcrystalline cellulose, or 1% brown rice protein, to set yogurt. The protein conjugate and brown rice protein considerably improved dry matter, water–holding capacity, firmness, consistency, viscosity index, and cohesiveness while decreasing syneresis in yogurt samples compared to plain yogurt. The application of protein conjugates also extends to plant–based dairy alternatives. Soy protein isolate–maltodextrin conjugates can be effectively used as substitutes for conventional proteins in various applications, including enhancing foaming stability and improving solubility (Choi et al., 2025). Similarly, microalgal proteins have emerged as promising ingredients for dairy alternatives, often outperforming terrestrial plant proteins because of their balanced amino acid profile and superior techno–functional properties (Samarathunga et al., 2025). In another investigation on ice cream incorporating WPI and rare sugars (D–allose and D–psicose) for glycation, it was reported that ice cream with WPI glycated with rare sugars had a higher overrun than ice cream with untreated WPI (Puangmanee et al., 2008).
In addition to protecting probiotics, protein conjugates markedly influence the flavor and physicochemical properties of dairy products. Key quality characteristics of yogurt, cheese, and milk–based beverages include enhanced emulsification capacity, gel–forming ability, and water–holding capacity, which define texture, viscosity, and mouthfeel (Mudgil et al., 2025). For instance, conjugates generated by the Maillard reaction between WPI and inulin have been shown in yogurt systems to reduce whey separation and increase product creaminess (Żbikowska et al., 2020). For dried milk, a conjugated WPH with maltodextrin was used as a carrier to protect B. animalis spp. lactis ATCC 27536 and L. acidophilus ATCC 4356 (Minj and Anand, 2022). The protein conjugate provided better probiotic protection during spray drying, resulting in a final probiotic count of 8.98 log CFU/g, compared to an initial count of 10.58 log CFU/mL before drying.
Gut microbiome analysis can be more effectively integrated into protein–probiotic conjugate design by using microbial composition and functional outputs to guide the selection of compatible prebiotics and probiotic strains for targeted outcomes. For instance, nondigestible dextrin, α–cyclodextrin, and dextran selectively enhance acetate and propionate production in human fecal fermentation models, highlighting how microbiome–driven SCFA profiles can inform conjugate design aimed at gut and metabolic health (Sasaki et al., 2018). Clinical evidence further demonstrates the value of microbiome–informed selection, as Lactobacillus–, Bifidobacterium–, and multi–strain formulations, such as a multi–strain formulation comprising four Lactobacillus strains, three Bifidobacterium strains, and one Streptococcus salivarius subsp. Thermophilus strain modulates inflammatory pathways, enhances mucosal barrier function, and reduces systemic markers of inflammation in inflammatory bowel disease and ulcerative colitis (Ganji–Arjenaki and Rafieian-Kopaei, 2018; Groeger et al., 2013). Together, these findings support a targeted design strategy in which microbiome profiling and host biomarkers directly inform the development of protein–probiotic conjugates with predictable anti–inflammatory and gut–protective effects.
Sensory attributes of protein conjugate containing dairy products
Gharibzahedi and Altintas (2024) further reported the sensory properties of set yogurt added with a protein conjugate, showing a slight difference with an optimum condition of 4.65% (w/w) transglutaminase–cross–linked protein isolate and 10.1 log CFU/mL L. acidophilus LA–5® producing yogurt with maximum lightness, firmness, apparent viscosity, and probiotic survivability, combined with minimum syneresis. Moreover, the yogurt with transglutaminase–cross–linked protein isolate exhibited less sensory deterioration than other yogurt samples during refrigerated storage over 2 weeks.
Nevertheless, the issue of product color after adding the protein conjugate was reported by researchers who supplemented the conjugate in ice cream and cream products. Seo and Yoo (2022b) used milk protein isolate–κ–carrageenan conjugates as part of the whipping cream ingredients and noted that excessively high levels of the conjugates affected the color of the whipping creams, making them appear more yellow A similar drawback was discovered if imitation Mozzarella cheese was supplemented with conjugates of cricket protein and FOS (Chailangka et al., 2023). The findings revealed that imitation Mozzarella cheese with 30% or higher conjugates received lower sensory scores for its appearance, color, odor, texture, flavor, and overall acceptability.
Protein sources in protein–probiotic conjugation
Protein conjugates from both milk and plant sources play important roles in stabilizing probiotics, enhancing their adhesion, and modulating host immunity. However, they function through distinct mechanisms and with varying effectiveness. Milk proteins, such as whey and casein, are particularly effective for microencapsulation, protecting probiotics during processing and GI transit, and their surface properties enhance adhesion to the intestinal mucosa (Abd El–Salam and El–Shibiny, 2015). They also influence gut immunity by promoting beneficial microbiota and SCFA production, thus reducing inflammation (Peled et al., 2024). In contrast, plant proteins primarily interact with exopolysaccharides in fermented products to maintain probiotic structural integrity and support functional effects, including antioxidative and anti–inflammatory activities. However, their encapsulation and immune–modulatory capacities are less well characterized (Zang et al., 2025). These findings suggest that milk–derived proteins offer robust and predictable benefits for probiotic stabilization and immune modulation. In contrast, plant proteins provide complementary advantages in specific applications, such as fermented foods, making the choice of protein system dependent on health goals, product type, and dietary considerations.
Role of Protein Conjugates in Delivering Nutritional and Health Advantages of Probiotics in Dairy Products
Probiotics can take the form of live cultures, inactivated cultures, postbiotics, and prebiotics, and are regarded as integral to functional dairy products. Owing to the presence of probiotics in dairy products, these products functionally improve overall digestive performance by influencing the intestinal microbiota, particularly when combined with protein conjugates. It has been found to confer various health benefits, including alleviating lactose intolerance, modulating the immune system, and controlling cholesterol levels (Mokhtari et al., 2019). Furthermore, it is asserted that probiotics, when combined or encapsulated in protein–conjugated products within a dairy product matrix, could enhance their benefits through the stabilization and delivery of probiotics, thereby increasing their bioavailability and modulating immune responses and gut flora (El Jeni et al., 2024).
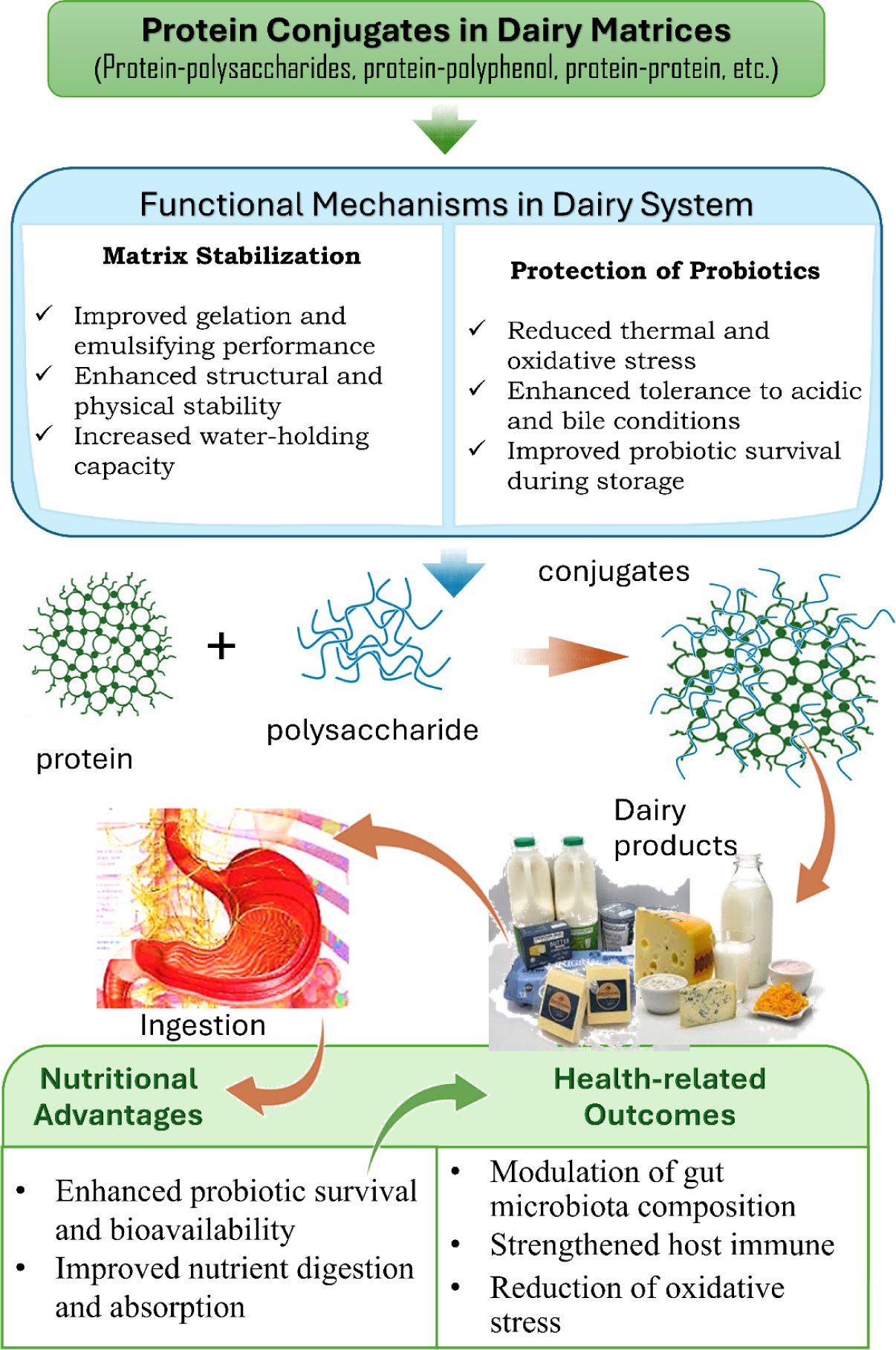
Recent studies also suggest that protein–carbohydrate conjugates may enhance the biofunctional activity of probiotic systems. After digestion, these conjugates release bioactive peptides that exhibit antioxidant, antibacterial, and antihypertensive properties (Minj and Anand, 2022). Additionally, oxygen–sensitive bacteria (A. muciniphila) have been encapsulated using emerging delivery methods, including those employing milk exosomes coupled to bioactive proteins. A recent in vitro study in mice has shown that such systems can enhance mucosal immunity and metabolic balance compared with unencapsulated systems (Hao et al., 2025). Protein conjugates also aid in regulating immune responses, thereby enhancing probiotics’ ability to support immune function (Hao et al., 2025). Moreover, they enhance the bioavailability of probiotics and bioactive peptides, thereby maximizing their health benefits (Hadjimbei et al., 2022; Li et al., 2023), as illustrated in Figure 3. Most evidence supporting the benefits of protein–probiotic conjugates remains largely confined to in vitro studies and animal models.
Figure 3. Overview of the functional roles of protein conjugates in probiotic dairy systems, highlighting their contribution to probiotic bioavailability, nutrient absorption, and host health protection. Source: Hadjimbei et al. (2022); Hao et al. (2025); Li et al. (2023); Lim et al. (2024); Rashidi et al. (2021).
Evidence studies on the nutritional and health roles of protein conjugates in probiotic–based dairy products
Recent studies have explored the potential of protein conjugates to enhance the nutritional and health–promoting effects of probiotics in food systems. For instance, in a study conducted by Massounga Bora et al. (2021), WPI and (−)–epigallocatechin–3–gallate conjugates were evaluated as carriers for L. acidophilus, demonstrating enhanced in vitro antioxidant (78%) and antidiabetic (52%) activities as well as improved probiotic survival ability and surface hydrophobicity. Similarly, Yu et al. (2021) examined the effects of casein and chicken–protein diets on recovery from dextran sulfate sodium (DSS)–induced colitis in mice. While DSS caused no physiological differences between the diet groups, the casein diet increased the beneficial gut bacteria. These findings collectively highlight the role of protein–based conjugates and dietary proteins in improving probiotic delivery, physiological functioning, and GI health.
Evidence suggests that protein conjugates enhance the survival, adhesion, and colonization of probiotics by forming protective matrices and promoting the targeted release of bioactive peptides and prebiotic–like sugars (Li et al., 2023; Mudgil et al., 2024a). Mechanistically, MTGase–mediated cross–linking strengthens hydrophobic and electrostatic interactions with intestinal epithelium, while the Maillard–derived reductones and melanoidins provide antioxidant protection to both probiotics and the host. Rashidi et al. (2021) reported that human clinical trials have demonstrated that dairy products containing protein–modified matrices increase Lactobacillus and Bifidobacterium abundance, improve cytokine profiles (e.g., elevated interleukin–10 [IL–10] and reduced tumor necrosis factor–alpha [TNF–α]) (Santiago–López et al., 2018), and enhance systemic antioxidant markers, confirming the translational potential of these functional dairy systems.
Microbiota modulation
The human gut microbiota, often referred to as a “forgotten organ,” comprises approximately 95% of all the cells in the human body and contains an estimated 1011–1012 CFU/g of intestinal content (Fucarino et al., 2022; Yao et al., 2021). The gut microbiota plays a vital role in maintaining epithelial barrier function and supporting metabolic and biochemical activities. Probiotics have been shown to promote intestinal eubiotics through various processes, including the synthesis of SCFAs, enhancing gut barrier integrity, competitive exclusion of pathogens, and modulation of immunological responses (Gao et al., 2022).
Protein conjugates modulate gut microbiota by enhancing probiotic adhesion and supplying selective metabolic substrates. MTGase–mediated casein conjugates strengthen hydrophobic and electrostatic interactions with intestinal epithelial cells, improving colonization and persistence (Gao et al., 2022; Wang et al., 2020). In parallel, the gradual hydrolysis of protein–carbohydrate conjugates releases short peptides and reducing sugars that act as prebiotic–like substrates, promoting the growth of Bifidobacterium and Lactobacillus while suppressing less favorable taxa through nutrient competition (Abdeen et al., 2024). Human studies support these mechanisms, showing an increased fecal abundance of beneficial genera and a favorable Firmicutes–Bacteroidetes ratio in subjects consuming protein–modified dairy matrices (Vivek et al., 2023). Thus, protein conjugates serve as both protective carriers and active modulators of microbiota composition.
Freeze–drying L. plantarum with whey protein and sodium alginate enhanced the survival of probiotics compared to spray drying (Liu et al., 2019). This treatment process is crucial for maintaining the viability of probiotics during processing conditions. Further supporting this, studies on probiotic cheese highlight the positive effects of probiotics on gut microbiota. For example, supplementation with goat cheese containing L. rhamnosus EM1107 in mice challenged with S. enteritidis decreased Salmonella colonization while promoting the development of beneficial bacteria (Rolim et al., 2021). Thus, the combination of protective carriers, such as whey protein and sodium alginate, during freeze–drying effectively enhances probiotic viability, which, as evidenced in probiotic–enriched cheese studies, translates into improved gut microbiota balance and pathogen suppression.
Studies have also investigated the various benefits of probiotics in ice cream, apart from the encapsulation process. Chaikham and Rattanasena (2017) studied ice cream complemented with L. acidophilus LA–5 and L. casei 01 probiotic flora, providing the update about the modulation effect of the product against colon microbiome, which led to an increase in helpful secondary metabolites (e.g., butyrate, acetate, lactic acid, and propionate) because of the supplementation. This demonstrates how probiotics in dairy products can contribute to the production of metabolites that enhance digestive health.
Immune response and regulation in dairy foods
The effectiveness of Lactobacillus and Bifidobacterium probiotics in dairy products supports their role in enhancing the immune system, thereby reducing the risk of inflammatory diseases. When incorporated into dairy products with protein conjugates, these probiotics often exhibit improved stability in dairy products; they are better protected during digestion when combined with protein conjugates, such as whey protein, which enhances and regulates gut homeostasis and immune functioning (Liu et al., 2019).
Protein conjugates enhance probiotic–host immune interactions by stabilizing cell–surface proteins and facilitating mucosal adhesion, which amplifies recognition by dendritic cells and epithelial receptors (e.g., toll–like receptors 2 and 4 (TLR2 and TLR4) (Gao et al., 2022). This interaction modulates cytokine profiles, increasing anti–inflammatory mediators (IL–10) while reducing pro–inflammatory markers (TNF–α and interleukins–6 [IL–6]) (Kariyawasam et al., 2021). Some conjugates also release bioactive peptides with direct immunomodulatory activity, such as opioid–like or angiotensin–converting enzyme (ACE) inhibitory peptides (Han et al., 2020). Human intervention studies with dairy–based protein conjugates have reported reductions in systemic inflammation and enhanced mucosal immunity, supporting their translational relevance.
A study conducted on fermented milk samples produced from a blend of Lactobacillus and Bifidobacterium strains demonstrated immune–enhancing effects. Consuming fermented milk containing the mentioned probiotics has been linked to increased immune markers, such as immunoglobulin A (IgA), IL–6, and IL–10, which are crucial for enhancing animal mucosal immunity (Santiago–López et al., 2018). The findings of this study provided information on how probiotic fermentation can more favorably modulate immune responses and contribute to the overall health benefits. Additionally, Mai et al. (2021) found that probiotic fermented milk has shown potential in alleviating digestive– and respiratory–related problems in young children. The findings indicated that L. casei Shirota in probiotic milk drastically reduced constipation, diarrhea, and respiratory infections in children aged 3–5 years.
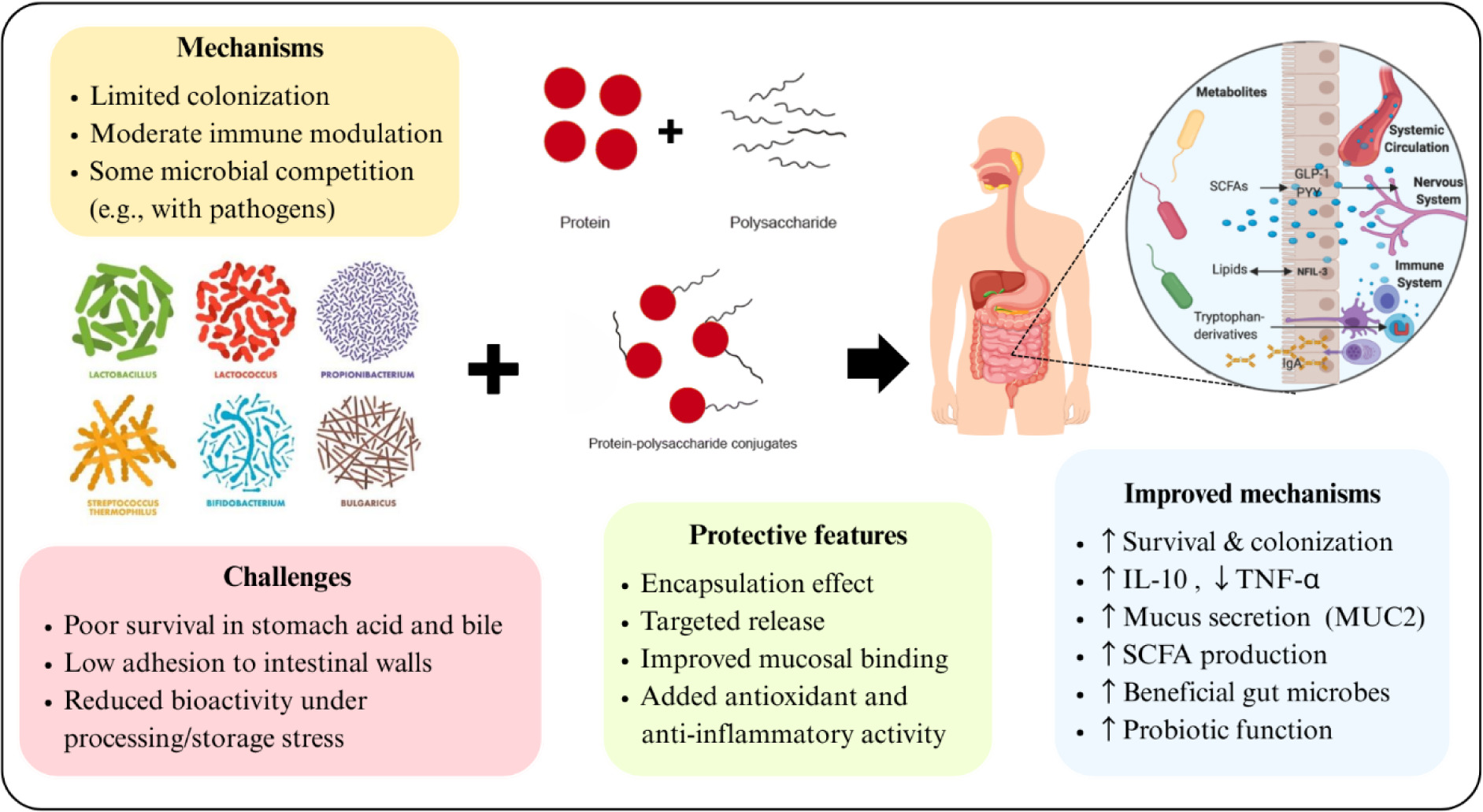
These findings suggest that adding protein conjugates to dairy–based probiotics enhances their capacity to modulate the immune system. As shown in Figure 4, protein conjugation enhances probiotic function, including improved gastric survival, targeted intestinal delivery, enhanced mucosal adhesion, and synergistic bioactivities (e.g., antioxidant and anti–inflammatory effects). Compared to free probiotics, conjugated probiotics more effectively modulate gut microbiota, strengthen the intestinal barrier, and support immune regulation.
Figure 4. Schematic illustration of the protective role of protein–polysaccharide conjugates in improving probiotic functionality. Native probiotics face challenges, such as poor survival in gastric and bile conditions, low intestinal adhesion, and reduced bioactivity during processing and storage. Source: Hao et al. (2025); Jang et al. (2024); Latif et al. (2023); Lim et al. (2024).
Furthermore, fermented milk with probiotic strains provides health benefits by producing bioactive peptides, including ACE inhibitors and antioxidative peptides. Production of high gamma–aminobutyric acid has been observed, with up to 5.5 g/L when S. thermophilus is present and up to 8.3 g/L when co–cultured with L. rhamnosus in casein hydrolysate–enriched milk (Han et al., 2020). Bifidobacterium and Lactobacillus probiotics are essential components of the human gut microbiota, playing a crucial role in restoring intestinal barrier function and enhancing immune function (El Jeni et al., 2024).
Although probiotic–protein immunomodulatory interactions remain underexplored, available evidence suggests that they are strain– and matrix–dependent. Recent studies have demonstrated that Lactobacillus strains can stimulate regulatory T cells to produce IL–10 and transforming growth factor–beta (TGF–β), increase IL–6 secretion in a TLR–2–dependent manner, and induce clonal expansion of IgA–producing B cells. Muhammad et al. (2026) showed that multi–species synbiotic supplementation, containing six probiotic species with inulin and FOS, significantly increased serum IL–10 levels and fecal SCFAs in adult men with dyslipidemia after 12 weeks. This highlights the need for systematic comparisons across probiotic strains and protein or prebiotic matrices to identify formulations with targeted immunomodulatory effects.
Bioavailability enhancement of probiotics and bioactive peptides
Protein conjugates play a crucial role in enhancing the bioavailability of probiotics in dairy products. They are commonly referred to as wall materials, including carbohydrates (e.g., starch and maltodextrin) and proteins (e.g., casein and whey protein). The absence of these wall materials can highlight the viability of probiotics during spray drying, resulting in a complete loss of viability of probiotics (Liu et al., 2019). Binary systems, which utilize proteins and polysaccharides to form conjugates, provide better protection for probiotic systems (Deng et al., 2025).
Protein conjugates enhance bioavailability by forming protective matrices that delay gastric degradation and facilitate targeted intestinal release (Mao et al., 2022; Wu et al., 2021b). Cross–linked proteins maintain probiotic viability until the colon while simultaneously enhancing solubility and uptake of associated bioactive peptides (Sbehat et al., 2022). Once released, peptides can cross epithelial barriers more efficiently because of improved resistance against proteolysis (Lemaire et al., 2021). Clinical studies using conjugated whey proteins in fermented dairy products have shown higher plasma levels of bioactive peptides and improved metabolic outcomes, underscoring the connection between structural protection and functional absorption (Gu et al., 2024; Huang et al., 2024).
Research has demonstrated that spray drying L. zeae LB1 with a combination of Arabic gum and sodium caseinate results in higher survival proportions at pH 2.0, compared to using sodium caseinate alone (Liu et al., 2016). Probiotics face stress in handling processes, storage, and digestive conditions, but their survival is critical for efficacy. The most common international standards on fermented products require a minimum of 107 CFU/g for L. acidophilus and 106 CFU/g for B. bifidum in fermented milk, with some specific variations in some countries (Stuivenberg et al., 2022). Ensuring probiotic viability through protein conjugate encapsulation helps to meet these regulatory requirements.
Protein–probiotic conjugates enhance probiotic stability during food processing, storage, and GI transit, although their effectiveness depends strongly on the protein matrix and conjugation strategy used. Plant– and dairy–based proteins, including rice, pea, whey, and soy protein isolates combined with inulin, have shown improved conjugation efficiency and protection of Lactobacillus reuteri against thermal and storage stresses, with rice and whey protein systems often providing superior stability (Mudgil et al., 2024b). Emerging technologies, such as 3D printing coupled with freeze–drying, further increase survival, achieving up to 96–98% viability of Lactiplantibacillus plantarum during extended storage and digestion (Yoha et al., 2021). In addition, highly resilient strains, such as bacillus coagulans, maintain over 99% viability under severe processing conditions, while alternative proteins, such as Spirulina isolates, show promising lyoprotective effects, outperforming whey protein under certain conditions (Maity et al., 2021). Consquently, selecting appropriate protein carriers and conjugation methods is critical for enhancing probiotic stability in functional food applications.
Antioxidant properties
Conjugation of milk carbohydrates with proteins through the Maillard reaction enhances protein functional properties, resulting in more heat–stable, soluble, and emulsified products (Zhang et al., 2020). Additionally, this conjugation enhances antioxidant activity by forming melanoidin and heterocyclic compounds, as well as reducing ketones, thereby improving the health–promoting properties of dairy products (Wu et al., 2021b). Specifically, when WPI is heated with reducing sugars, the Maillard reaction enhances its reducing power and scavenging capacity, improving its antioxidant properties (Zhang et al., 2020). Additionally, a study has shown that the synergistic effect of ascorbic acid enhances the survival of L. acidophilus in yogurt. At the same time, enzyme–based scavengers, such as glucose oxidase, have been found to increase B. longum counts by up to 40% (Gupta, 2024).
Accordingly, protein conjugation stabilizes the food matrix while introducing antioxidant–active groups, particularly reductones and melanoidins, which scavenge reactive oxygen species (ROS) and chelate pro–oxidant metal ions (Wu et al., 2021b). This dual action mitigates oxidative stress in the gut, protecting encapsulated probiotics while simultaneously delivering systemic antioxidant benefits to the host (Noori et al., 2025). Clinical investigations corroborate these effects, demonstrating that dairy products enriched with protein conjugates improve redox balance, as evidenced by increased total antioxidant capacity and decreased lipid peroxidation in human subjects (Lemaire et al., 2021; Mudgil et al., 2024a).
Moreover, synbiotic yogurts have gained popularity because of their antioxidant, anti–inflammatory, and antihypertensive properties, which have a positive impact on the overall health. For example, synbiotic formulations containing L. brevis KU200019 and FOS have exhibited antioxidant, ACE–inhibitory, and immune–modulating effects in dairy products (Kariyawasam et al., 2021). Yogurt containing L. rhamnosus and B. lactis improves gut motility and relieves irritable bowel syndrome (IBS) (Olson and Aryana, 2022). Similarly, cheese fortified with L. casei and L. plantarum has been linked to reduced blood pressure and body mass index (BMI) (Shah et al., 2024). Additionally, ice cream enriched with L. acidophilus LA–5 and L. casei 01 increased the modulation of SCFAs production and promoted beneficial gut microbiota (Chaikham and Rattanasena, 2017). Another probiotic ice cream containing L. lactis NZ1330 has been shown to reduce Immunoglobulin E (IgE) levels and allergic responses in mice (Vasiee et al., 2020). Therefore, the antioxidant properties of dairy probiotic products suggest that their presence supports the health–promoting features of these products, as probiotics in them have been observed to remain qualitatively intact during GI transit. Beyond direct food applications, protein conjugates have found innovative uses in active packaging systems. For instance, whey protein–polyphenol conjugates incorporated into carboxymethyl cellulose films demonstrate enhanced vapor barrier properties, tensile strength, and intelligent pH–sensing capabilities for monitoring of freshness (Zhao et al., 2023). This innovative approach offers potential for extending the shelf life of dairy products while preserving probiotic viability.
Challenges Associated with Protein Conjugates in Dairy Foods
Hussain et al. (2021) postulated that the use of protein conjugation in dairy probiotics presents both problems and opportunities that must be addressed carefully prior to its application in any domain. Among the discrepancies are the technological challenges that have become most common. The issues of stability and the need for efficient delivery systems add to the complexity of conjugation methods, making it more challenging to incorporate them into protein conjugates containing probiotic dairy products. Another significant barrier to the market’s widespread acceptance of protein–conjugated probiotics is the complex regulations governing the clearance process and safety investigations for novel compounds (Cerk and Aguilera-Gómez, 2022). Consumer acceptance and market perception also predispose the success of protein–conjugated probiotics. Purchase decisions are influenced by a product’s efficacy, safety, and health benefits, as evidenced by a study on the development of the Maillard reaction in UHT milk under varying storage temperatures and thermal cycles (Sunds et al., 2018).
Technological challenges
Probiotics face significant challenges in maintaining viability and functionality throughout the manufacturing, processing, and storage of dairy products. The main factors affecting the survival and stability of probiotics in dairy foods are low temperatures, pH levels, and the composition of the probiotics (Deng et al., 2025; Mazhar et al., 2024). Thermal destabilization and low microbial growth present crucial challenges in dairy product manufacturing, as high–protein dairy matrices, such as evaporated milk, undergo structural changes under heat stress that compromise stability and probiotic viability. Wu et al. (2021a) investigated the modification of milk proteins via glycation via the Maillard reaction to enhance functionality by improving heat stability, eliminating the need for harmful catalysts, and optimizing the performance of dairy proteins during thermal processing.
Furthermore, whey proteins have relatively weak emulsifying properties, leading to instability, turbidity, and phase separation in acidic dairy products, especially during storage and heat exposure (Dıblan et al., 2024). Co–encapsulation with prebiotics serves as a potential strategy to enhance probiotic survival by protecting the cells from environmental stress during processing, storage, and digestion (Rashidinejad et al., 2022). Therefore, addressing these technological challenges through advanced protein conjugation and co–encapsulation strategies is a key to improving the stability and functionality of probiotics in dairy applications.
Regulatory barriers and safety concerns of protein conjugation
The practical applicability of protein–conjugation in dairy probiotics is linked to specific regulatory challenges. The US Food and Drug Administration (FDA) and the European Food Safety Authority (EFSA) are regulatory agencies that require comprehensive safety data, including toxicity and allergen tests, before these products are permitted for human consumption, as reported by Cerk and Aguilera–Gómez (2022) and Zavišić et al. (2023). Strict rules must be backed by solid scientific research when labeling products to make health claims, such as those that boost immunity. Although both EFSA and FDA have established regulations for probiotics and food additives, a gray area remains regarding protein–probiotic conjugates with exception of the study reported by Awaluddin et al. (2025), in which the study confirmed that synbiotic systems exhibit superior biological functionality, compared with probiotic or prebiotic alone, reflecting coordinated regulation of gut structure, nutrient bioavailability, and host performance. Manufacturing these conjugates at a larger scale while maintaining consistent quality and stability can be challenging, and additional steps may be required if the ingredient is considered novel (Spacova et al., 2023).
Moreover, while protein conjugation can enhance probiotic stability and functionality, one critical safety concern is the unintended formation of AGEs, which have been implicated in various metabolic and chronic diseases. The accumulation of AGEs in serum induces oxidative stress and activates the chromogranin A–uncoupling protein 2–glucose transporter 1 pathway, contributing to diabetes–related complications (Sun et al., 2023). Beyond diabetes, dietary AGEs exacerbate inflammation and oxidative stress, representing risk factors for chronic liver diseases. Another study confirmed that higher intake of AGEs, particularly Nε–(carboxymethyl) lysine and methylglyoxal–derived hydroimidazolone–1, were positively correlated with the incidence of fatty liver disease (Xie et al., 2023). To counteract these risks, several strategies are explored to inhibit the formation of AGEs. Approaches include blocking early glycation intermediates, preventing protein cross–linking, chelating metal ions, and suppressing AGE to their receptor (AGE–RAGE) signaling through antioxidant and anti–inflammatory pathways. Potential interventions involve glycation inhibitors (e.g., aminoguanidine and pyridoxamine), AGE breakers (e.g., alagebrium chloride), metal chelators, and natural phytochemical antioxidants (Gu et al., 2024). Excessive heating causes browning, loss of amino acid, and formation of AGE; thus, Bi et al. (2023) recommended 70°C for 15 s to preserve quality and nutrition in dairy systems.
Protein conjugation, while promising for improving stability and functionality, raises critical concerns regarding allergenicity Spacova et al. (2023). Structural modifications can mask epitopes and sometimes reduce IgE–binding, but uncontrolled conjugation reactions may expose new epitopes or generate neoallergens, increasing sensitization risks (Vasiee et al., 2020). Enzymatic methods, such as MTGase, further complicate this issue by altering the conformational epitopes of caseins and whey proteins, which can unpredictably affect immune recognition (Chen et al., 2020). Evidence to date is dominated by in vitro IgE–binding assays and rodent models, both of which have limited predictive value for human allergic responses (Cerk and Aguilera-Gómez, 2022). This lack of translational data creates uncertainty for regulatory assessment, as agencies, such as EFSA and FDA, mandate rigorous evaluation of allergenicity for novel protein modifications, including structural, digestibility, and clinical immunogenicity data. Without systematic human studies, the allergenic safety of conjugated milk proteins remains unresolved, highlighting the urgent need for targeted allergenicity assessments before such technologies can be safely applied in probiotic dairy foods.
Future perspectives and research needs in protein conjugation–based dairy products
The advent of protein conjugates in dairy products presents both opportunities and challenges from consumer and technological perspectives, as summarized in Table 4. They enhance probiotic survival, emulsification, and solubility, but also introduce off–flavors and bitterness if the Maillard reaction is not carefully controlled (Sunds et al., 2018). Consumer resistance to genetically modified organisms (GMOs), particularly in regions such as the European Union (EU), supports the use of non–GMO approaches, such as protein conjugation, to enhance acceptance (Börner et al., 2019; Hussain et al., 2021). As gut health remains a primary interest in probiotic consumption, improving formulations to show that clear health benefits can enhance consumer appeal (Gao et al., 2021; Rashidinejad et al., 2022). Therefore, optimization of the Maillard reaction is also crucial to avoid negative impacts on the appearance and flavor of products (Wu et al., 2021b).
Table 4. Constraints and the way forward on applying protein conjugates in probiotic dairy products.
| Category | Description | Possible solutions | References |
|---|---|---|---|
| Technological barriers | |||
| Thermal destabilization | Heat treatments impact the viability of probiotics and the conjugate fatty acid content. Conjugated fatty acid degradation | Microencapsulation or co–encapsulation with prebiotics to protect probiotics. Improving fermentation conditions to preserve fatty acids. Also, use of moderate temperatures (60–70°C), RH: 65–75%, for 24–48h depending on protein–sugar system; monitor browning index or absorbance at 420nm; for lactose–free milk, 70°C for 15s preserves nutrition conjugates. |
Bi et al., 2023; Mao et al., 2022; Wu et al., 2021a |
| Low microbial growth in dairy matrices | Environmental stresses in high–protein dairy products can inhibit the growth of probiotics. | Selecting probiotic strains that can withstand stress with oxygen scavengers, such as tocopherols. Use strains like L. rhamnosusGG, L. plantarumWCFS1, B. longumBB536; choose strains with EPS production, thick cell walls, and robust membrane fatty acids to enhance thermal and acid tolerance. |
Guo et al., 2022; He et al., 2025; Liao et al., 2021 |
| Phase separation, poor solubility, and emulsification | Because whey protein concentrates are poorly soluble, they become turbid and unstable in acidic drinks. | Glycation enhances the solubility and functional qualities of whey proteins. Conjugated polysaccharides improve stability and emulsifying qualities. | Dıblan et al., 2024 |
| Encapsulation efficiency | Probiotic survival can be enhanced by utilizing optimized co–encapsulation methods that incorporate prebiotics. | By enhancing probiotic protection with the use of multi–layer microencapsulation. | Rashidinejad et al., 2022 |
| Raw milk availability (input procurement) | Environmental disruptions (e.g., volcanic eruption) reduce dairy cattle populations and milk yield, threatening stable raw material supply for functional dairy production. | Diversifying milk sourcing, integration of milk powder or alternative protein sources, and developing shelf–stable protein conjugates to buffer supply volatility. | Guntoro et al., 2025 |
| Technology adoption | Modern technologies (financial, communication, and product) can enhance dairy cooperative efficiency and financial performance. | Capacity building, tax incentives, supportive policies, and training programs. | Nyakio Ireri et al., 2025 |
| Regulatory hurdles | |||
| Novel food approval and GRAS certification | Protein conjugation in probiotics is a novel approach, thus requiring extensive approval and meeting GRAS criteria for market acceptance. | Communicate safety studies, including toxicity and allergenicity assessments, to regulatory agencies for approval and submission. | Cerk and Aguilera- Gómez, 2022; Spacova et al., 2023 |
| Labeling and health claims | Strict regulations on health claims related to probiotics and protein conjugates. | By ensuring clinical trial–backed claims that are scientifically validated. Moreover, uniform standards for labeling protein–conjugated probiotics should be established. EU: EFSA Novel Food Regulation 2015/2283; US: FDA GRAS new dietary ingredient notifications provide compositional, toxicological, and allergenicity data; document safe processing and residual cross–linkers. |
Aduah et al., 2024 |
| Consumer expectations and market perception | |||
| Consumer expectations | Protein conjugates can enhance the survival of probiotics but may also lead to off–flavors and bitterness. | The Maillard reaction is carefully controlled to prevent the development of unwanted flavors. Protein conjugates enhance emulsification and solubility while reducing bitterness. Use approved food–grade cross–linkers (e.g., genipin) at the minimal effective concentrations; remove any unreacted chemicals; and evaluate sensory properties in pilot formulations. |
Laurujisawat et al., 2025; Sunds et al., 2018; Wei and Huang, 2019 |
| Consumer perception of GMO | GMO products face consumer resistance in regions such as the EU. | Non–GMO methods (e.g., protein conjugation) may improve consumer acceptance of dairy probiotics. | Börner et al., 2019; Hussain et al., 2021 |
Notes: GRAS: generally recognized as safe; EPS: extracellular polymeric substances; GMO: Genetically modified organisms.
Protein conjugates offer functional benefits, such as enhanced probiotic delivery; however, they may negatively affect color and flavor (Deng et al., 2025; Wu et al., 2021b). Incorporating postbiotics, including those derived from L. acidophilus, further enhances immune responses. Using strains such as L. reuteri and other fermented probiotic derivatives can strengthen these effects (Garnier et al., 2019). Additionally, probiotic–based exopolysaccharides enhance the rheological properties of dairy products, improving their texture, flavor, and consumer appeal while contributing to health benefits (Ali et al., 2023; Zhao and Liang, 2023). Thus, future studies are expected to provide information on how these upcoming industrial processes may affect probiotic dairy foods in a practical setting.
Future research should combine detailed mechanistic investigations, such as molecular signaling, microbial adhesion, and bioactive peptide release, with well–designed human clinical trials to validate the effects of protein conjugates on probiotic functionality, gut microbiota modulation, and immune responses within real–world dairy matrices. Further cost–benefit analysis of protein conjugation is required to assess its economic feasibility and guide industrial–scale implementation while balancing functional efficacy and product quality.
Study Limitations
Although this review provides an integrated overview of protein conjugation strategies and their impact on probiotic functionality in dairy systems, several limitations must be considered. First, most of the available evidence is derived from in vitro experiments and animal models, which do not fully capture the complexity of human digestive and immune responses. For example, allergenicity assessments rely heavily on rodent models and IgE–binding assays, both of which exhibit limited predictive value for human outcomes (Cerk and Aguilera–Gómez, 2022; Vasiee et al., 2020). Similarly, most studies evaluating thermal stability, GI tolerance, or encapsulation performance examine individual probiotic strains, despite clear strain–dependent variability in conjugation efficiency and functional response (Li et al., 2025; Liao et al., 2021; Mudgil et al., 2024b). Second, variations in processing conditions, such as temperature, RH, pH, and ionic strength, that affect reproducibility and hinder cross–study comparison. Lack of standardized protocols for the Maillard reaction, enzymatic, or chemical conjugation leads to difficulties in identifying optimal conditions and determining scalability for industrial applications (Gao et al., 2021; Wu et al., 2021b). Additionally, most current reports focus on small–scale laboratory processes, with limited evaluation of pilot– or industrial–scale production, limiting insights into cost, energy demand, and process robustness (Parhi et al., 2024). Moreover, the potential formation of AGEs during the Maillard conjugation presents unresolved safety concerns. Although the existing research highlights correlations between dietary AGEs and inflammatory or metabolic disorders (Sun et al., 2023; Xie et al., 2023), there is still limited research that measures AGE formation in conjugated dairy systems or examines the potential long–term exposure risks in humans. Finally, the regulatory landscape for protein–probiotic conjugates remains underdeveloped. Limited human clinical trials, insufficient allergenicity data, and unclear classification frameworks restrict the translation of these technologies into commercial dairy products (Cerk and Aguilera–Gómez, 2022; Spacova et al., 2023). Addressing these limitations requires coordinated efforts to develop standardized methodologies, conduct well–designed human studies, and establish regulatory guidance specific to conjugated protein systems.
Conclusions
Protein conjugation, through processes such as the Maillard–type and enzymatic cross–linking, has demonstrated considerable potential in enhancing the functionality and viability of probiotics in dairy systems. These conjugates act as dual–function systems, providing a protective barrier for probiotic cells under adverse conditions, such as heat, acidity, and bile salts, while simultaneously improving texture, emulsifying properties, and the overall stability within dairy matrices. Consistent findings across various applications continue to show improvements in encapsulation efficiency, thermal tolerance, and water–holding capacity, which together support the broader integration of conjugated systems in both fermented and nonfermented dairy products.
Despite these advantages, broader application remains limited by several factors, including the need for tighter control of the Maillard reaction conditions to prevent excessive browning and the formation of undesirable compounds, along with the continued challenge of managing potential sensory changes and navigating complex regulatory frameworks related to food safety and functional claims. Consequently, future research on protein conjugation should prioritize the refinement of reaction conditions, particularly through optimization of the Maillard reaction and enzymatic pathways to improve reproducibility, minimize undesirable end products, and enhance safety. In addition, exploring novel protein and carbohydrate sources with promising functional and nutritional attributes may further expand the applicability of conjugates across diverse food systems.
Personalized functional dairy foods represent a promising direction, emerging from the integration of gut microbiome profiles, metabolic and immune biomarkers, and indicators of gut–barrier function. When combined with predictive modeling approaches, including machine–learning (ML)–based diet–microbiome response tools, these developments open the possibility for designing protein–probiotic conjugates that align more closely with individual health objectives, such as metabolic regulation, GI tolerance, and immune modulation. Within this framework, protein conjugation offers opportunities to improve nutrient interactions, modulate allergenicity, and selectively influence host–microbiota dynamics.
However, translating these advances into practical applications require comprehensive cost–benefit evaluations and well–designed human clinical studies to verify efficacy and safety beyond laboratory and animal evidence. At the same time, successful commercialization depends on achieving industrial scalability through environmentally responsible and economically viable processing strategies, supported by artificial intelligence (AI)–driven and omics–based formulation tools that strengthen process robustness and accelerate innovation. Ultimately, long–term progress depends on adherence to international regulatory frameworks to ensure that functional claims are scientifically substantiated and that consumer safety is maintained consistently.
Data Availability
The data are available upon request.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Author Contributions
Conceptualization, writing–original draft preparation, and writing–review and editing: Nareekan Chaiwong, Auengploy Chailangka, Idris Kaida Zubairu, Tri Indrarini Wirjantoro, Pavalee Chompoorat Tridtitanakia, and Mohsen Gavahian; writing–review and editing: Juan Manuel Castagnini and Mahsa Majzoobi; supervision, conceptualization, funding acquisition, and writing–review and editing: Yuthana Phimolsiripol. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declared no conflict of interest.
Funding
This research was funded by the “Reinventing University Program” and “International Research Fellowship at Chiang Mai University (Visiting Professor),” Chiang Mai University, Thailand. In addition, thanks to the Center of Excellence at Agro Bio–Circular–Green Industry, Chiang Mai University.
REFERENCES
Abdeen, E.S.M., Hamed, A.M., and Ismail, H.A. 2024. Production and evaluation of novel functional cream cottage cheese fortified with bovine colostrum and probiotic bacteria. Journal of Food Science and Technology 61(8): 1457–1469. 10.1007/s13197-023-05910-0
Abd El–Salam, M.H., and El–Shibiny, S. 2015. Preparation and properties of milk proteins–based encapsulated probiotics: a review. Dairy Science & Technology 95(4): 393–412. 10.1007/s13594-015-0223-8
Aduah, M., Adzitey, F., Yusuf, K.A., and Huda, N. 2024. Market, regulatory and consumer challenges of functional meat products. In: Rather, S.A., Masoodi, F.A. (Eds.), Handbook of Processed Functional Meat Products. Springer, Cham, Switzerland, pp. 545–564. 10.1007/978-3-031-69868-2_20
Akalan, M., Karakuş, M.Ş., Özaslan, Z.T., Başyiğit, B., Karaaslan, A., and Karaaslan, M. 2024. Brown rice protein–dietary fibre conjugate improves bioaccessibility of minerals via modifying gelling structure of yoghurt. International Journal of Food Science & Technology 59(11): 8851–8863. 10.1111/ijfs.17085
Akbal, S., Uğur Geçer, E., and Ertürkmen, P. 2025. Probiotic viability and bioactive properties of buffalo yoghurt produced using high cholesterol-assimilating probiotic strains. Veterinary Medicine and Science 11(2): e70233. 10.1002/vms3.70233
Ali, A.H., Bamigbade, G., Tarique, M., Esposito, G., Obaid, R., AbuJdayil, B., and Ayyash, M. 2023. Physicochemical, rheological, and bioactive properties of exopolysaccharide produced by a potential probiotic Enterococcus faecalis 84B. International Journal of Biological Macromolecules 240: 124425. 10.1016/j.ijbiomac.2023.124425
Al–Sulbi, O.S., and Shori, A.B. 2022. Viability of selected strains of probiotic Lactobacillus spp., and sensory evaluation of concentrated yogurt (labneh) made from cow, camel, and cashew milk. Food Science and Technology 42: e113321. 10.1590/fst.113321
Awaluddin, A., Rahardja, D.P., and Mujnisa, A. 2025. The effects of prebiotic, probiotic and synbiotic supplementation on the performance, small intestinal morphometry, and carcass traits of broiler chicken. Journal of Global Innovations in Agricultural Sciences 13(2): 485–493. 10.22194/JGIAS/25.1553
A’yun, Q., Demicheli, P., De Neve, L., Wu, J., Balcaen, M., Setiowati, A.D., Martins, J.C., Van Troys, M., and Van Der Meeren, P. 2020. Dry heat induced whey protein–lactose conjugates largely improve the heat stability of O/W emulsions. International Dairy Journal 108: 104736. 10.1016/j.idairyj.2020.104736
Benedetti, L., Cattaneo, L., Vercesi, A., Trevisi, E., and Piccioli–Cappelli, F. 2024. Effects of live Saccharomyces cerevisiae yeast administration in periparturient dairy cows. Animals 14(3): 472–472. 10.3390/ani14030472
Bi, H., Wang, Y., Guo, Y., Liao, Z., and Na, Z. 2023. Influence of pasteurization on Maillard reaction in lactose–free milk. Molecules 28(20): 7105. 10.3390/molecules28207105
Bönisch, E., Oh, Y.J., Anzengruber, J., Hager, F.F., López–Guzmán, A., Zayni, S., Hinterdorfer, P., Kosma, P., Messner, P., Duda, K.A., and Schäffer, C. 2018. Lipoteichoic acid mediates binding of a Lactobacillus S–layer protein. Glycobiology 28(3): 148–158. 10.1093/glycob/cwx102.
Börner, R.A., Kandasamy, V., Axelsen, A.M., Nielsen, A.T., and Bosma, E.F. 2019. Genome editing of lactic acid bacteria: opportunities for food, feed, pharma and biotech. FEMS Microbiology Letters 366(1): fny291. 10.1093/femsle/fny291
Cerk, K., and Aguilera-Gómez, M. 2022. Microbiota analysis for risk assessment: evaluation of hazardous dietary substances and its potential role on the gut microbiome variability and dysbiosis. EFSA Journal 20: e200404. 10.2903/j.efsa.2022.e200404
Chaikham, P., and Rattanasena, P. 2017. Combined effects of low–fat ice cream supplemented with probiotics on colon microfloral communities and their metabolites during fermentation in a human gut reactor. Food Bioscience 17: 35–41. 10.1016/j.fbio.2016.12.005
Chailangka, A., Leksawasdi, N., Seesuriyachan, P., Ruksiriwanich, W., Sommano, S.R., Jantanasakulwong, K., Rachtanapun, P., Castagnini, J.M., Barba, F.J., and Phimolsiripol, Y. 2023. Improving vitamin D stability and antioxidant activity in imitation mozzarella cheese by conjugated cricket protein with fructooligosaccharide. Food Science and Technology (LWT) 183: 114898. 10.1016/j.lwt.2023.114898
Chaiwong, N., Seesuriyachan, P., Rachtanapun, P., Gavahian, M., Bangar, S.P., Khaneghah, A.M., Wangtueai, S., Leksawasdi, N., Jantanasakulwong, K., Chailangka, A., Sommano, S.R., Castagnini, J.M., and Phimolsiripol, Y. 2025. Enhancing solubility, emulsion properties, and antioxidant activity of whey protein powder via wet−heating conjugated with galactooligosaccharides. Journal of Agriculture and Food Research 19: 101666. 10.1016/j.jafr.2025.101666
Chen, C.C., Chen, L.Y., Chan, D.S., Chen, B.Y., Tseng, H.W., and Hsieh, J.F. 2020. Influence of microbial transglutaminase on physicochemical and cross–linking characteristics of individual caseins. Molecules 25(17): 3992. 10.3390/molecules25173992
Choi, H.W., Kim, J.H., Ham, S.H., Park, C., Kim, J.W., Hahn, J., and Choi, Y.J. 2025. Effect of heating time and drying method on the functional properties of soy protein isolate–maltodextrin conjugates for plant–based meringue cookies. Future Foods 11: 100587. 10.1016/j.fufo.2025.100587
D’Amico, V., Cavaliere, M., Ivone, M., Lacassia, C., Celano, G., Vacca, M., Maria, F., Fontana, S., Angelis, M.D., Denora, N., and Lopedota, A.A. 2025. Microencapsulation of probiotics for enhanced stability and health benefits in dairy functional foods: a focus on pasta filata cheese. Pharmaceutics 17(2): 185. 10.3390/pharmaceutics17020185
da Silva, T.M., de Deus, C., de Souza Fonseca, B., Lopes, E.J., Cichoski, A.J., Esmerino, E.A., Cristiane, S., Muller, E.I., Flores, E.M.M., and de Menezes, C.R. 2019. The effect of enzymatic crosslinking on the viability of probiotic bacteria (Lactobacillus acidophilus) encapsulated by complex coacervation. Food Research International 125: 108577. 10.1016/j.foodres.2019.108577
Deng, Y., Kolodjski, S., Lewis, G., Onan, G., and Kim, Y. 2025. Physical and functional characterization of whey protein–lignin biocomposite films for food packaging applications. Future Foods 11: 100554. 10.1016/j.fufo.2025.100554
Dıblan, S., Salum, P., Ulusal, F., and Erbay, Z. 2024. Impact of conjugation of whey protein concentrate with different carbohydrates: monitoring structural and technofunctional variations. International Dairy Journal 158: 106036. 10.1016/j.idairyj.2024.106036
Dimitrellou, D., Sakadani, E., and Kandylis, P. 2025. Enhancing probiotic viability in yogurt: the role of apple fibers in supporting Lacticaseibacillus casei ATCC 393 during storage and gastrointestinal transit. Foods 14(3): 376. 10.3390/foods14030376
Doğan, M., and Ay, M. 2021. Evaluation of the probiotic potential of Pediococcus strains from fermented dairy product kefir. Czech Journal of Food Sciences 39(5): 376–383. 10.17221/71/2021-cjfs
Dong, L., Li, Y., Chen, Q., Liu, Y., Qiao, Z., Sang, S., Zhang, J., Zhan, S., Wu, Z., and Liu, L. 2023. Research advances of advanced glycation endproducts in milk and dairy products: formation, determination, control strategy and immunometabolism via gut microbiota. Food Chemistry 417: 135861. 10.1016/j.foodchem.2023.135861
Du, T., Liu, Z., Guan, Q., Xiong, T., and Peng, F. 2023. Application of soy protein isolate–xylose conjugates for improving the viability and stability of probiotics microencapsulated by spray drying. Journal of the Science of Food and Agriculture 103(13): 6500–6509. 10.1002/jsfa.12728
El Jeni, R., Villot, C., Koyun, O., Osorio–Doblado, A., Baloyi, J., Lourenco, J., Steele, M., and Callaway, T. 2024. Invited review: “probiotic” approaches to improving dairy production: reassessing “magic foo–foo dust.” Journal of Dairy Science 107(4): 1832–1856. 10.3168/jds.2023-23831
Fucarino, A., Burgio, S., Paladino, L., Caruso Bavisotto, C., Pitruzzella, A., Bucchieri, F., and Cappello, F. 2022. The microbiota is not an organ: introducing the muco–microbiotic layer as a novel morphofunctional structure. Anatomia 1(2): 186–203. 10.3390/anatomia1020019
Ganji-Arjenaki, M., and Rafieian-Kopaei, M. 2018. Probiotics are a good choice in remission of inflammatory bowel diseases: a meta–analysis and systematic review. Journal of Cellular Physiology 233(3): 2091–2103. 10.1002/jcp.25911
Gao, J., Li, X., Zhang, G., Sadiq, F.A., Simal–Gandara, J., Xiao, J., and Sang, Y. 2021. Probiotics in the dairy industry–advances and opportunities. Comprehensive Reviews in Food Science and Food Safety 20(4): 3937–3982. 10.1111/1541-4337.12755
Gao, X., Zhao, J., Zhang, H., Chen, W., and Zhai, Q. 2022. Modulation of gut health using probiotics: the role of probiotic effector molecules. Journal of Future Foods 2(1): 1–12. 10.1016/j.jfutfo.2022.03.011
Ganji–Arjenaki, M., & Rafieian–Kopaei, M. 2018. Probiotics are a good choice in remission of inflammatory bowel diseases: a meta analysis and systematic review. Journal of cellular physiology, 233(3), 2091–2103. 10.1002/jcp.25911
Garnier, L., Mounier, J., Lê, S., Pawtowski, A., Pinon, N., Camier, B., Chatel, M., Garric, G., Thierry, A., Coton, E., and Valence, F. 2019. Development of antifungal ingredients for dairy products: from in vitro screening to pilot scale application. Food Microbiology 81: 97–107. 10.1016/j.fm.2018.11.003
Gavahian, M., Mathad, G.N., Oliveira, C.A., and Khaneghah, A.M. 2021. Combinations of emerging technologies with fermentation: interaction effects for detoxification of mycotoxins? Food Research International 141: 110104. 10.1016/j.foodres.2021.110104
Gharibzahedi, S.M.T., and Altintas, Z. 2024. Transglutaminase–crosslinked lesser mealworm protein isolate: a new milk fat substitute for high–quality probiotic set yogurts. Food Hydrocolloids 146: 109172. 10.1016/j.foodhyd.2023.109172
Goh, K.K.T., Sarkar, A., and Singh, H. 2020. Milk protein–polysaccharide interactions. In: Boland, M., Singh, H. (Eds.), Milk proteins (Third Edition). Academic Press, Elsevier, Amsterdam, The Netherlands, pp. 387–419. 10.1016/B978-0-12-815251-5.00013-X
Gouseti, O., Larsen, M.E., Amin, A., Bakalis, S., Petersen, I.L., Lametsch, R., and Jensen, P.E. 2023. Applications of enzyme technology to enhance transition to plant proteins: a review. Foods 12(13): 2518. 10.3390/foods12132518
Groeger, D., O’Mahony, L., Murphy, E.F., Bourke, J.F., Dinan, T.G., Kiely, B., Shanahan, F., and Quigley, E.M.M. 2013. Bifidobacterium Infantis 35624 modulates host inflammatory processes beyond the gut. Gut Microbes 4(4): 325–339. 10.4161/gmic.25487
Gu, M.J., Lee, Y.R., Kim, D., Kim, Y., and Ha, S.K. 2024. Comprehensive research on the properties of advanced glycation end products in food and biological samples and their harmful role in inducing metabolic diseases. Comprehensive Reviews in Food Science and Food Safety 23(5): e13412. 10.1111/1541-4337.13412
Guntoro, B., Guntoro, T.M., and Hoang Qui, N. 2025. Production characteristics and risk analysis of dairy farming enterprise in Sleman District, Yogyakarta, Indonesia. Journal of Global Innovations in Agricultural Sciences 13(4): 1457–1464. 10.22194/JGIAS/25.1696
Guo, Q., Tang, J., Li, S., Qiang, L., Chang, S., Du, G., Yue, T., and Yuan, Y. 2022. Lactobacillus plantarum 21805 encapsulated by whey protein isolate and dextran conjugate for enhanced viability. International Journal of Biological Macromolecules 216: 124–131. 10.1016/j.ijbiomac.2022.06.207
Gupta, P. 2024. Role of oxygen absorbers in food as packaging material, their characterization and applications. Journal of Food Science and Technology 61(2): 242–252. 10.1007/s13197-023-05681-8
Hadjimbei, E., Botsaris, G., and Chrysostomou, S. 2022. Beneficial effects of yoghurts and probiotic fermented milks and their functional food potential. Foods 11(17): 2691. 10.3390/foods11172691
Halavach, T.M. 2024. Efficient enzymatic hydrolysis of the whey protein fraction from bovine milk. Journal of Global Innovations in Agricultural Sciences 12(3): 743–749. 10.22194/JGIAS/24.1362
Han, M., Liao, W., Wu, S., Gong, X., and Bai, C. 2020. Use of Streptococcus thermophilus for the in situ production of γ–aminobutyric acid–enriched fermented milk. Journal of Dairy Science 103(1): 98–105. 10.3168/jds.2019-16856
Hao, L., Liu, Y., Szeto, I.M.Y., Hao, H., Zhang, T., Liu, T., and Yi, H. 2025. Milk exosome–based delivery system for probiotic encapsulation that enhances the gastrointestinal resistance and adhesion of probiotics. Nutrients 17(5): 923–923. 10.3390/nu17050923
Hasnain, M.A., Kang, D.K., and Moon, G.S. 2024. Research trends of next generation probiotics. Food Science and Biotechnology 33(9): 2111–2121. 10.1007/s10068-024-01626-9
He, X., Cui, Y., Jia, Q., Zhuang, Y., Gu, Y., Fan, X., and Ding, Y. 2025. Response mechanisms of lactic acid bacteria under environmental stress and their application in the food industry. Food Bioscience 64: 105938. 10.1016/j.fbio.2025.105938
He, X., Yang, W., and Qin, X. 2023. Ultrasound–assisted multilayer Pickering emulsion fabricated by WPI–EGCG covalent conjugates for encapsulating probiotics in colon–targeted release. Ultrasonics Sonochemistry 97: 106450. 10.1016/j.ultsonch.2023.106450
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Morelli, L., Canani, R.B., Flint, H.J., Salminen, S., Calder, P.C., and Sanders, M.E. 2014. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews. Gastroenterology & Hepatology 11(8): 506–514. 10.1038/nrgastro.2014.66
Hou, C., Wu, S., Xia, Y., Phillips, G.O., and Cui, S.W. 2017. A novel emulsifier prepared from Acacia seyal polysaccharide through Maillard reaction with casein peptides. Food Hydrocolloids 69: 236–241. 10.1016/j.foodhyd.2017.01.038
Huang, Y., Lu, Y., Liu, X., Wu, W., Li, W., Lai, S., Aadil, R.M., Rajoka, M.S.R., Wang, L., and Zeng, X. 2024. Metabolic properties, functional characteristics and practical application of Streptococcus thermophilus. Food Reviews International 40(2): 792–813. 10.1080/87559129.2023.2202406
Hussain, N., Li, R., Takala, T.M., Tariq, M., Zaidi, A.H., and Saris, P.E.J. 2021. Generation of lactose–and protease–positive probiotic Lacticaseibacillus rhamnosus GG by conjugation with Lactococcus lactis NCDO 712. Applied and Environmental Microbiology 87(6): e02957-20. 10.1128/aem.02957-20
Ibrahim, S.A., Ayivi, R.D., Zimmerman, T., Siddiqui, S.A., Altemimi, A.B., Fidan, H., Esatbeyoglu, T., and Bakhshayesh, R.V. 2021. Lactic acid bacteria as antimicrobial agents: food safety and microbial food spoilage prevention. Foods 10(12): 3131. 10.3390/foods10123131
Jang, H.J., Lee, N.K., and Paik, H.D. 2024. Overview of dairy–based products with probiotics: fermented or non–fermented milk drink. Food Science of Animal Resources 44(2): 255–268. 10.5851/kosfa.2023.e83
Kariyawasam, K.M.G.M., Lee, N.K., and Paik, H.D. 2021. Fermented dairy products as delivery vehicles of novel probiotic strains isolated from traditional fermented Asian foods. Journal of Food Science and Technology. 58: 2467–2478. 10.1007/s13197-020-04857-w
Kathuria, D., Gautam, S., and Thakur, A. 2023. Maillard reaction in different food products: effect on product quality, human health and mitigation strategies. Food Control 153: 109911. 10.1016/j.foodcont.2023.109911
Kaur, H., Kaur, G., and Ali, S.A. 2022. Dairy–based probiotic–fermented functional foods: an update on their health–promoting properties. Fermentation 8(9): 425. 10.3390/fermentation8090425
Khan, H., Mudgil, P., Alkaabi, S.A.S., Al Rashdi, Y.H.S., and Maqsood, S. 2024. Maillard reaction–based conjugation of pea protein and prebiotic polydextrose: optimization, characterization and functional properties enhancement. Frontiers in Sustainable Food Systems 8: 1463058. 10.3389/fsufs.2024.1463058
Kondrotiene, K., Zavistanaviciute, P., Aksomaitiene, J., Novoslavskij, A., and Malakauskas, M. 2023. Lactococcus lactis in dairy fermentation–health–promoting and probiotic properties. Fermentation 10(1): 16. 10.3390/fermentation10010016
Kowalczyk, M., Znamirowska–Piotrowska, A., Buniowska–Olejnik, M., and Pawlos, M. 2022. Sheep milk symbiotic ice cream: effect of inulin and apple fiber on the survival of five probiotic bacterial strains during simulated in vitro digestion conditions. Nutrients 14(21): 4454. 10.3390/nu14214454
Latif, A., Shehzad, A., Niazi, S., Zahid, A., Ashraf, W., Iqbal, M.W., Rehman, A., Riaz, T., Aadil, R.M., Khan, I., Özogul, F., Rocha, J.M., Esatbeyoglu, T., and Korma, S.A. 2023. Probiotics: mechanism of action, health benefits and their application in food industries. Frontiers in Microbiology 14(21): 4454. 10.3389/fmicb.2023.1216674
Laurujisawat, P., Dumrongchai, T., Rodklongtan, A., and Chitprasert, P. 2025. Genipin–cross–linked whey protein isolate for enhanced probiotic viability: balancing protein structure and cell interactions during spray–dried encapsulation, storage, and digestion. Food Science and Technology (LWT) 216: 117347. 10.1016/j.lwt.2025.117347
Lee, A.H., Rodriguez Jimenez, D.M., and Meisel, M. 2025. Limosilactobacillus reuteri–a probiotic gut commensal with contextual impact on immunity. Gut Microbes 17(1) : article 2451088. 10.1080/19490976.2025.2451088
Lemaire, M., Ménard, O., Cahu, A., Nogret, I., BriardBion, V., Cudennec, B., Cuinet, I., Ruyet, L., Baudry, C., Dupont, D., Blat, S., Deglaire, A., and HuërouLuron, L. 2021. Addition of dairy lipids and probiotic lactobacillus fermentum in infant formulas modulates proteolysis and lipolysis with moderate consequences on gut physiology and metabolism in Yucatan piglets. Frontiers in Nutrition 8: 615248. 10.3389/fnut.2021.615248
Li, H., Peng, F., Peng, Z., Liu, Z., Huang, T., and Xiong, T. 2023. Protection effect of gelatin–xylooligosaccharides Maillard reaction products on spray–dried Limosilactobacillus fermentum and possible action mechanism. Food Bioscience 56: 103251. 10.1016/j.fbio.2023.103251
Li, W., Zhao, Y., Li, S., Yun, L., Wu, T., and Zhang, M. 2025. Improving the physical stability of Lactobacillus plantarum LP90 during storage by mixing carboxymethylated dextran–whey protein conjugates and small–molecule sugars. Food Research International 203: 115834. 10.1016/j.foodres.2025.115834
Liao, Y., Hu, Y., Fu, N., Hu, J., Xiong, H., Chen, X.D., and Zhao, Q. 2021. Maillard conjugates of whey protein isolate–xylooligosaccharides for the microencapsulation of Lactobacillus rhamnosus: protective effects and stability during spray drying, storage and gastrointestinal digestion. Food & Function 12(9): 4034–4045. 10.1039/d0fo03439h
Lim, T.W., Lim, R.L.H., Pui, L.P., Tan, C.P., and Ho, C.W. 2024. Evaluating the potential of stabilised betacyanins from fermented red dragon fruit (Hylocereus polyrhizus) drink: sustainable colouration and antioxidant enhancement of stirred yoghurt. Future Foods 10: 100452. 10.1016/j.fufo.2024.100452
Liu, H., Cui, S.W., Chen, M., li, Y., Liang, R., Xu, F., and Zhong, F. 2019. Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: a review. Critical Reviews in Food Science and Nutrition 59(17): 2863–2878. 10.1080/10408398.2017.1377684
Liu, H., Gong, J., Chabot, D., Miller, S.S., Cui, S.W., Ma, J., Zhong, F., and Wang, Q. 2016. Incorporation of polysaccharides into sodium caseinate–low melting point fat microparticles improves probiotic bacterial survival during simulated gastrointestinal digestion and storage. Food Hydrocolloids 54: 328–337. 10.1016/j.foodhyd.2015.10.016
Liu, Y., Liu, Y., Zhong, M., Tuly, J.A., Dong, W., Ren, X., and Ma, H. 2025. Protein–polysaccharide complexes for O/W emulsions: structural engineering and their relevance to food stability and safety. Comprehensive Reviews in Food Science and Food Safety 24(5): e70289. 10.1111/1541-4337.70289
Liu, X., Xia, B., Hu, L., Ni, Z., Thakur, K., and Wei, Z. 2020. Maillard conjugates and their potential in food and nutritional industries: a review. Food Frontiers 1(4): 382–397. 10.1002/fft2.43
Liu, X., Xue, F., and Adhikari, B. 2023. Production of hemp protein isolate–polyphenol conjugates through ultrasound and alkali treatment methods and their characterization. Future Foods 7: 100210. 10.1016/j.fufo.2022.100210
Loyeau, P.A., Spotti, M.J., Vinderola, G., and Carrara, C.R. 2021. Encapsulation of potential probiotic and canola oil through emulsification and ionotropic gelation, using protein/polysaccharides Maillard conjugates as emulsifiers. Food Science and Technology (LWT) 150: 111980. 10.1016/j.lwt.2021.111980
Mai, T.T., Thi Thu, P., Thi Hang, H., Trang, T.T.T., Yui, S., Shigehisa, A., Tien, V.T., Dung, T.V., Nga, P.B., Hung, N.T., and Tuyen, L.D. 2021. Efficacy of probiotics on digestive disorders and acute respiratory infections: a controlled clinical trial in young Vietnamese children. European Journal of Clinical Nutrition 75(3): 513–520. 10.1038/s41430-020-00754-9
Maity, C., Bagkar, P., Dixit, Y., Tiwari, A., and Gupta, A.K. 2020. Process and storage stability of Bacillus coagulans LBSC in food matrices and appraisal of calorific restriction: B. coagulans LBSC stability and calorie restriction. Applied Food Biotechnology 8(1): 57–69. 10.22037/afb.v8i1.31212
Manyatsi, T.S., Mousavi Khaneghah, A., and Gavahian, M. 2024. The effects of ultrasound on probiotic functionality: an updated review. Critical Reviews in Food Science and Nutrition 64(31): 11643–11660. 10.1080/10408398.2023.2242490
Mao, B., Guo, W., Huang, Z., Tang, X., Zhang, Q., Yang, B., Zhao, J., Cui, S., and Zhang, H. 2022. Production of conjugated fatty acids in probiotic–fermented walnut milk with the addition of lipase. Food Science and Technology (LWT) 172: 114204. 10.1016/j.lwt.2022.114204
Massounga Bora, A.F., Li, X., Liu, L., and Zhang, X. 2021. Enhanced in vitro functionality and food application of Lactobacillus acidophilus encapsulated in a whey protein isolate and (−)–Epigallocatechin–3–Gallate conjugate. Journal of Agricultural and Food Chemistry 69(37): 11074–11084. 10.1021/acs.jafc.1c02158
Mazhar, S., Simon, A., Khokhlova, E., Colom, J., Leeuwendaal, N., Deaton, J., and Rea, K. 2024. In vitro safety and functional characterization of the novel Bacillus coagulans strain CGI314. Frontiers in Microbiology 14: 1302480. 10.3389/fmicb.2023.1302480
Milczek, E.M. 2018. Commercial applications for enzyme–mediated protein conjugation: new developments in enzymatic processes to deliver functionalized proteins on the commercial scale. Chemical Reviews 118(1): 119–141. 10.1021/acs.chemrev.6b00832
Minj, S., and Anand, S. 2022. Development of a spray–dried conjugated whey protein hydrolysate powder with entrapped probiotics. Journal of Dairy Science 105(3): 2038–2048. 10.3168/jds.2021-20978
Moiseenko, K.V., Glazunova, O.A., Savinova, O.S., Shabaev, A.V., and Fedorova, T.V. 2023. Changes in composition of some bioactive molecules upon inclusion of Lacticaseibacillus paracasei probiotic strains into a standard yogurt starter culture. Foods 12(23): 4238. 10.3390/foods12234238
Mokhtari, S., Jafari, S.M., and Khomeiri, M. 2019. Survival of encapsulated probiotics in pasteurized grape juice and evaluation of their properties during storage. Food Science and Technology International 25(2): 120–129. 10.1177/1082013218801113
Mudgil, P., Ajayi, F.F., Alyafei, A.S., Yap, P.G., Gan, C.Y., and Maqsood, S. 2024a. Unlocking the hypolipidemic potential of bioactive peptides derived from probiotic fermented cattle, camel, goat, and sheep milk: a comprehensive investigation through in vitro, in silico, and molecular docking studies. Frontiers in Sustainable Food Systems 8: 1443708. 10.3389/fsufs.2024.1443708
Mudgil, P., Alalawi, M., Alghaithi, M., Mustapha, A.T., and Maqsood, S. 2025. Modification of camel milk proteins through complexation with green solvent extracted date seed polyphenols: effects on physiochemical, technofunctional properties and bioactive properties. International Dairy Journal 164: 106188. 10.1016/j.idairyj.2025.106188
Mudgil, P., Alkaabi, F., Khan, H., Javed, M., Hajamohideen, A.R., Hamed, F., and Maqsood, S. 2024b. Enhanced viability and stability of the Lactobacillus reuteri DSM 17938 probiotic strain following microencapsulation in pea and rice protein–inulin conjugates. Frontiers in Sustainable Food Systems 8: 1284069. 10.3389/fsufs.2024.1284069
Muhammad, S., Li, M., Jia, Q., Ijaz, M., Liang, S., Zeng, W., Chen, D., Zhang, Y., Du, X., Song, W., Guo, B. 2026. Advances in the engineering of living probiotics for cancer immunotherapy. Theranostics 16(3): 1164–1226. 10.7150/thno.125301
Nghiep, H.X. 2024. Effects of energy feed combinations and non–protein nitrogen on feed intake and nutrient digestibility in Saanen crossbred goats. Journal of Global Innovations in Agricultural Sciences 12(4): 1171–1177. 10.22194/JGIAS/24.1535
Noori, M., Shateri, Z., Babajafari, S., Eskandari, M.H., Parastouei, K., Ghasemi, M., Afshari, H., and Samadi, M. 2025. The effect of probiotic–fortified kefir on depression, appetite, oxidative stress, and inflammatory parameters in Iranian overweight and obese elderly: a randomized, double–blind, placebo–controlled clinical trial. Journal of Health, Population and Nutrition 44(1): 30. 10.1186/s41043-025-00773-x
Nyakio Ireri, S.S., and Gatimbu, K.K. 2025. The nexus between technology integration and financial performance of dairy cooperatives in Mt. Kenya Region, Kenya. Journal of Global Innovations in Agricultural Sciences 13(4): 1481–1491. 10.22194/JGIAS/25.1710
Oana, K., Shimizu, K., Takada, T., Makino, H., Yamazaki, M., Katto, M., Ando, M., Kurakawa, T., and Oishi, K. 2023. Manipulating the growth environment through co–culture to enhance stress tolerance and viability of probiotic strains in the gastrointestinal tract. Applied and Environmental Microbiology 89(12): e0150223. 10.1128/aem.01502-23
Olson, D.W., and Aryana, K.J. 2022. Probiotic incorporation into yogurt and various novel yogurt–based products. Applied Sciences 12(24): 12607. 10.3390/app122412607
Pan, Y., Wu, Z., Xie, Q.T., Li, X.M., Meng, R., Zhang, B., and Jin, Z.Y. 2020. Insight into the stabilization mechanism of emulsions stabilized by Maillard conjugates: protein hydrolysates–dextrin with different degree of polymerization. Food Hydrocolloids 99: 105347. 10.1016/j.foodhyd.2019.105347
Parhi, P., Liu, S.Q., and Choo, W.S. 2024. Synbiotics: effects of prebiotics on the growth and viability of probiotics in food matrices. Bioactive Carbohydrates and Dietary Fibre 32: 100462. 10.1016/j.bcdf.2024.100462
Peled, S., Freilich, S., Hanani, H., Kashi, Y., and Livney, Y.D. 2024. Next–generation prebiotics: Maillard–conjugates of 2'–fucosyllactose and lactoferrin hydrolysates beneficially modulate gut microbiome composition and health promoting activity in a murine model. Food Research International 177: 113830. 10.1016/j.foodres.2023.113830
Peled, S., and Livney, Y.D. 2021. Oligosaccharide–lactoferrin shell–crosslinked particles for selective targeting of proteins to probiotic bacteria in the colon. Food Hydrocolloids 120: 106973. 10.1016/j.foodhyd.2021.106973
Puangmanee, S., Hayakawa, S., Sun, Y., and Ogawa, M. 2008. Application of whey protein isolate glycated with rare sugars to ice cream. Food Science and Technology Research 14(5): 457–466. 10.3136/fstr.14.457
Rabah, H., Rosa do Carmo, F., and Jan, G. 2017. Dairy propionibacteria: versatile probiotics. Microorganisms 5(2): 24. 10.3390/microorganisms5020024
Rashidi, K., Razi, B., Darand, M., Dehghani, A., Janmohammadi, P., and Alizadeh, S. 2021. Effect of probiotic fermented dairy products on incidence of respiratory tract infections: a systematic review and meta–analysis of randomized clinical trials. Nutrition Journal 20(1): 1–20. 10.1186/s12937-021-00718-0
Rashidinejad, A., Bahrami, A., Rehman, A., Rezaei, A., Babazadeh, A., Singh, H., and Jafari, S.M. 2022. Co–encapsulation of probiotics with prebiotics and their application in functional/synbiotic dairy products. Critical Reviews in Food Science and Nutrition 62(9): 2470–2494. 10.1080/10408398.2020.1854169
Rodrigues, P.D., Fernandes, I., de A.A., de Marins, A.R., Feihrmann, A.C., and Gomes, R.G. 2024. Use of mucilage from Opuntia ficus–indica in the manufacture of probiotic cream cheese. Processes 12(10): 2289. 10.3390/pr12102289
Rolim, F.R.L., Oliveira, C.J.B., de Freitas Neto, O.C., Dos Santos, K.M.O., Guerra, G.C.B., Rodrigues, R.V., de Assis, P.O.A., Araújo, D.F., de S., de Carvalho, V.A.G., Lemos, M.L.P., da Silva, N.M.V., Soares, J.K.B., Garcia, H.E.M., de Souza, E.L., Souza, F., de A.L., de Barros, M.E.G., de Oliveira, M.E.G., and Queiroga, R.C.R.E. 2021. Microbiological, immunological, and histological changes in the gut of Salmonella Enteritidis–challenged rats fed goat cheese containing Lactobacillus rhamnosus EM1107. Journal of Dairy Science 104(1): 179–197. 10.3168/jds.2020-18820
Samarathunga, J., Phuong, T., Gabard, M., Strazdins, K., Rens, J., and Adhikari, B., 2025. Microalgal proteins as ingredients for crating dairy mimetic products: prospects for substituting bovine milk proteins. Future Foods 11: 100556. 10.1016/j.fufo.2025.100556
Santiago–López, L., Hernández–Mendoza, A., Mata–Haro, V., Vallejo–Cordoba, B., and González–Córdova, A.F. 2018. Immune response induced by fermented milk with potential probiotic strains isolated from artisanal Cocido cheese. Food and Agricultural Immunology 29(1): 911–929. 10.1080/09540105.2018.1485632
Santoso, T., Ho, T.M., Vinothsankar, G., Jouppila, K., Chen, T., Owens, A., Lazarjani, M.P., Farouk, M.M., Colgrave, M.L., Otter, D., Kam, R., and Le, T.T. 2024. Effects of laccase and transglutaminase on the physicochemical and functional properties of hybrid lupin and whey protein powder. Foods 13(13): 2090. 10.3390/foods13132090
Sasaki, D., Sasaki, K., Ikuta, N., Yasuda, T., Fukuda, I., Kondo, A., and Osawa, R. 2018. Low amounts of dietary fibre increase in vitro production of short–chain fatty acids without changing human colonic microbiota structure. Scientific Reports 8(1): 435. 10.1038/s41598-017-18877-8
Sbehat, M., Mauriello, G., and Altamimi, M. 2022. Microencapsulation of probiotics for food functionalization: an update on literature reviews. Microorganisms 10(10): 1948. 10.3390/microorganisms10101948
Seo, C.W., and Oh, N.S. 2022a. Functional application of Maillard conjugate derived from a κ–carrageenan/milk protein isolate mixture as a stabilizer in ice cream. Food Science and Technology (LWT) 161: 113406. 10.1016/j.lwt.2022.113406
Seo, C.W., and Yoo, B. 2022b. Effect of milk protein isolate/κ–carrageenan conjugates on rheological and physical properties of whipping cream: a comparative study of Maillard conjugates and electrostatic complexes. Food Science of Animal Resources 42(5): 889–902. 10.5851/kosfa.2022.e42
Shah, A.B., Baiseitova, A., Zahoor, M., Ahmad, I., Ikram, M., Bakhsh, A., Shah, M.A., Ali, I., Idress, M., Ullah, R., Nasr, F.A., and Al–Zharani, M. 2024. Probiotic significance of Lactobacillus strains: a comprehensive review on health impacts, research gaps, and future prospects. Gut Microbes 16(1): 2431643. 10.1080/19490976.2024.2431643
Shen, Y., Miao, C., Ma, M., Zhen, Z., He, J., Pei, X., Zhang, Y., Man, C., Zhao, Q., and Jiang, Y. 2024. Mechanistic insights into the changes of biological activity and physicochemical characteristics in Lacticaseibacillus paracasei fortified milk powder during storage. Food Chemistry 452: 139501. 10.1016/j.foodchem.2024.139501
Sibanda, T., Marole, T.A., Thomashoff, U.L., Thantsha, M.S., and Buys, E.M. 2024. Bifidobacterium species viability in dairy–based probiotic foods: challenges and innovative approaches for accurate viability determination and monitoring of probiotic functionality. Frontiers in Microbiology 15: 1327010. 10.3389/fmicb.2024.1327010
Sim, S.Y.J., SRV, A., Chiang, J.H., and Henry, C.J. 2021. Plant proteins for future foods: a roadmap. Foods 10(8): 1967. 10.3390/foods10081967
Song, H., Li, R., Yang, L., Wang, S., Zhang, Y., Li, J., Zhu, D., and Liu, H. 2024. Encapsulation of probiotic Lacticaseibacillus casei with whey protein isolate/soybean hull polysaccharide enhances cell viability in harsh conditions. Food Hydrocolloids 154: 110126. 10.1016/j.foodhyd.2024.110126
Song, W.F., Yao, W.Q., Chen, Q.W., Zheng, D., Han, Z.Y., and Zhang, X.Z. 2022. In Situ bioorthogonal conjugation of delivered bacteria with gut inhabitants for enhancing probiotics colonization. ACS Central Science 8(9): 1306–1317. 10.1021/acscentsci.2c00533
Spacova, I., Binda, S., Ter Haar, J.A., Henoud, S., Legrain–Raspaud, S., Dekker, J., Espadaler–Mazo, J., Langella, P., Martín, R., Pane, M., and Ouwehand, A.C., 2023. Comparing technology and regulatory landscape of probiotics as food, dietary supplements and live biotherapeutics. Frontiers in Microbiology 14: 1272754. 10.3389/fmicb.2023.1272754
Stuivenberg, G.A., Daisley, B.A., Akouris, P.P., and Reid, G. 2022. In vitro assessment of histamine and lactate production by a multi–strain synbiotic. Journal of Food Science and Technology 59(9): 3419–3427. 10.1007/s13197-021-05327-7
Sun, K.X., Chen, Y.Y., Li, Z., Zheng, S.J., Wan, W.J., Ji, Y., and Hu, K. 2023. Genipin relieves diabetic retinopathy by down–regulation of advanced glycation end products via the mitochondrial metabolism–related signaling pathway. World Journal of Diabetes 14(9): 1349–1368. 10.4239/wjd.v14.i9.1349
Sun, X., Wang, H., Li, S., Song, C., Zhang, S., Ren, J., and Udenigwe, C.C. 2022. Maillard–type protein–polysaccharide conjugates and electrostatic protein–polysaccharide complexes as delivery vehicles for food bioactive ingredients: formation, types, and applications. Gels 8(2):135. 10.3390/gels8020135
Sunds, A.V., Rauh, V.M., Sørensen, J., and Larsen, L.B. 2018. Maillard reaction progress in UHT milk during storage at different temperature levels and cycles. International Dairy Journal 77: 56–64. 10.1016/j.idairyj.2017.08.008
Talearngkul, R., Sae-tan, S., and Sirivarasai, J. 2023. Effect of yogurt ice cream on the viability and antidiabetic potential of the probiotics Lactobacillus acidophilus, Lacticaseibacillus rhamnosus, and Bifidobacterium animalis subsp. lactis after in vitro digestion. Foods 12(23): 4373. 10.3390/foods12234373
Urango, A.C.M., Meireles, M.A.A., and Silva, E.K. 2024. Maillard conjugates produced from proteins and prebiotic dietary fibers: technological properties, health benefits and challenges. Trends in Food Science & Technology 147: 104438. 10.1016/j.tifs.2024.104438
Vasiee, A., Falah, F., Sankian, M., Tabatabaei–Yazdi, F., and Mortazavi, S.A. 2020. Oral immunotherapy using probiotic ice cream containing recombinant food–grade Lactococcus lactis which inhibited allergic responses in a BALB/c mouse model. Journal of Immunology Research 2020: 2635230. 10.1155/2020/2635230
Vivek, K., Mishra, S., Pradhan, R.C., Nagarajan, M., Kumar, P.K., Singh, S.S., Manvi, D., and Gowda, N.N. 2023. A comprehensive review on microencapsulation of probiotics: technology, carriers and current trends. Applied Food Research 3(1): 100248. 10.1016/j.afres.2022.100248
Wang, S., Xiao, Y., Tian, F., Zhao, J., Zhang, H., Zhai, Q., and Chen, W. 2020. Rational use of prebiotics for gut microbiota alterations: specific bacterial phylotypes and related mechanisms. Journal of Functional Foods 66: 103838. 10.1016/j.jff.2020.103838
Wei, Z., and Huang, Q. 2019. Assembly of protein–polysaccharide complexes for delivery of bioactive ingredients: a perspective paper. Journal of Agricultural and Food Chemistry 67(5): 1344–1352. 10.1021/acs.jafc.8b06063
Werasakulchai, A., Ngamwonglumlert, L., Chiewchan, N., Yoovidhya, T., and Devahastin, S. 2025. Pumpkin–based carotenoids complexed with dairy and plant proteins: stability of complexes and characteristics of their spray–dried powders. Future Foods 11: 100629. 10.1016/j.fufo.2025.100629
Wu, J., Chen, S., Sedaghat Doost, A., A’yun, Q., and Van Der Meeren, P. 2021a. Dry heat treatment of skim milk powder greatly improves the heat stability of recombined evaporated milk emulsions. Food Hydrocolloids 112: 106342. 10.1016/j.foodhyd.2020.106342
Wu, Q., Kan, J., Cui, Z., Ma, Y., Liu, X., Dong, R., Huang, D., Chen, L., Du, J., and Fu, C. 2024. Understanding the nutritional benefits through plant proteins–probiotics interactions: mechanisms, challenges, and perspectives. Critical Reviews in Food Science and Nutrition 65(18): 3569–3587. 10.1080/10408398.2024.2369694
Wu, J., Li, H., A’yun, Q., Sedaghat Doost, A., De Meulenaer, B., and Van der Meeren, P. 2021b. Conjugation of milk proteins and reducing sugars and its potential application in the improvement of the heat stability of (recombined) evaporated milk. Trends in Food Science & Technology 108: 287–296. 10.1016/j.tifs.2021.01.019
Xiang, J., Liu, F., Wang, B., Chen, L., Liu, W., & Tan, S. 2021. A literature review on Maillard reaction based on milk proteins and carbohydrates in food and pharmaceutical products: advantages, disadvantages, and avoidance strategies. Foods, 10(9), 1998. 10.3390/foods10091998
Xie, F., Zhao, J., Liu, D., Wan, Z., Sun, K., and Wang, Y. 2023. Associations of dietary advanced glycation end products with liver steatosis via vibration controlled transient elastography in the United States: a nationwide cross–sectional study. European Journal of Nutrition 63: 173–183. 10.1007/s00394-023-03253-2
Xu, Z.Z., Huang, G.Q., Xu, T.C., Liu, L.N., and Xiao, J.X. 2019. Comparative study on the Maillard reaction of chitosan oligosaccharide and glucose with soybean protein isolate. International Journal of Biological Macromolecules 131: 601–607. 10.1016/j.ijbiomac.2019.03.101
Yao, Y., Cai, X., Ye, Y., Wang, F., Chen, F., and Zheng, C. 2021. The role of microbiota in infant health: from early life to adulthood. Frontiers in Immunology 12: 708472. 10.3389/fimmu.2021.708472
Yi, J., Chen, X., Wen, Z., and Fan, Y. 2024. Improving the functionality of pea protein with laccase–catalyzed crosslinking mediated by chlorogenic acid. Food Chemistry 433: 137344. 10.1016/j.foodchem.2023.137344
Yin, L., Han, S., Xiaowen, J., Liu, J., Chen, Z., Yang, X., He, Z., Sun, M., Sun, J., and Gao, H. 2025. Probiotic–drug conjugates achieve synchronized site–specific probiotic colonization and on–demand drug release against ulcerative colitis and its complication. Journal of the American Chemical Society 147(34): 30924–30942. 10.1021/jacs.5c08094
Yoha, K.S., Anukiruthika, T., Anila, W., Moses, J.A., and Anandharamakrishnan, C. 2021. 3D printing of encapsulated probiotics: effect of different post–processing methods on the stability of Lactiplantibacillus plantarum (NCIM 2083) under static in vitro digestion conditions and during storage. Food Science and Technology (LWT) 146: 111461. 10.1016/j.lwt.2021.111461
Yousefvand, A., Huang, X., Zarei, M., and Saris, P.E.J. 2022. Lacticaseibacillus rhamnosus GG survival and quality parameters in kefir produced from kefir grains and natural kefir starter culture. Foods 11(4): 523. 10.3390/foods11040523
Yu, L., Zhao, D., Nian, Y., and Li, C. 2021. Casein–fed mice showed faster recovery from DSS–induced colitis than chicken–protein–fed mice. Food & Function 12(13): 5806–5820. 10.1039/d1fo00659b
Yuan, M., Cao, Y., Zheng, H., Chen, K., Lu, Y., Wang, J., Zhu, L., Chen, M., Cai, Z., and Shen, Y. 2024. Structural and functional properties of Maillard–reacted casein phosphopeptides with different carbohydrates. Food Science and Biotechnology 33(7): 1603–1614. 10.1007/s10068-023-01463-2
Zang, J., Lin, T., Shi, Y., Lin, Y., Xu, C., Ma, K., Zhang, C., Rui, X., Gan, D., and Li, W. 2025. Recent advances in exopolysaccharide–protein interactions in fermented dairy-and plant-based yogurts: mechanisms, influencing factors, health benefits, analytical techniques, and future directions. Comprehensive Reviews in Food Science and Food Safety 24(4): e70219. 10.1111/1541-4337.70219
Zanjani, M.A.K., Ehsani, M.R., Tarzi, B.G., and Sharifan, A. 2018. Promoting probiotics survival by microencapsualtion with Hylon starch and genipin cross–linked coatings in simulated gastro–intestinal condition and heat treatment. Iranian Journal of Pharmaceutical Research (IJPR) 17(2): 753. 10.22037/ijpr.2018.2199
Zareie, Z., Moayedi, A., Tabar–Heydar, K., Khomeiri, M., Maghsoudlou, Y., and Garavand, F. 2025. Enhancing the microbial dynamics, volatile profile, and ripening efficiency of white brined cheese using Lactiplantibacillus plantarum L33 as a probiotic co–culture. Food Research International 203: 115912. 10.1016/j.foodres.2025.115912
Zavišić, G., Popović, M., Stojkov, S., Medić, D., Gusman, V., Lješković, N.J., and Galović, A.J. 2023. Antibiotic resistance and probiotics: knowledge gaps, market overview and preliminary screening. Antibiotics 12(8): 1281–1281. 10.3390/antibiotics12081281
Żbikowska, A., Szymańska, I., and Kowalska, M. 2020. Impact of inulin addition on properties of natural yogurt. Applied Sciences 10(12): 4317. 10.3390/app10124317
Zhang, X., Li, X., Lü, L., Wang, L., Massounga, F., and Du, L. 2020. Covalent conjugation of whey protein isolate hydrolysates and galactose through Maillard reaction to improve the functional properties and antioxidant activity. International Dairy Journal 102: 104584. 10.1016/j.idairyj.2019.104584
Zhang, Z., Wang, B., Holden, G., Chen, J., and Adhikari, B. 2023a. Improving functional properties of Spirulina protein by covalent conjugation followed by complex coacervation processes. Future Foods 7: 100239. 10.1016/j.fufo.2023.100239
Zhang, S., Wang, K., Qin, Y., Zhu, S., Gao, Q., and Liu, D. 2023b. The synthesis, biological activities and applications of protein–polysaccharide conjugates in food system: a review. Food Quality and Safety 7: fyad006. 10.1093/fqsafe/fyad006
Zhao, X., Li, C., and Xue, F. 2023. Effects of whey protein–polyphenol conjugates incorporation on physicochemical and intelligent pH–sensing properties of carboxymethyl cellulose based films. Future Foods 7: 100211. 10.1016/j.fufo.2022.100211
Zhao, X., and Liang, Q. 2023. Optimization, probiotic characteristics, and rheological properties of exopolysaccharides from Lactiplantibacillus plantarum MC5. Molecules 28(6): 2463. 10.3390/molecules28062463
Zheng, B.D., Gan, L., Tian, L.Y., and Chen, G.H. 2023. Protein/polysaccharide–based hydrogels loaded probiotic–mediated therapeutic systems: a review. International Journal of Biological Macromolecules 253: 126841. 10.1016/j.ijbiomac.2023.126841
Zhong, S.R., Li, M.F., Zhang, Z.H., Zong, M.H., Wu, X.L., and Lou, W.Y. 2021. Novel antioxidative wall materials for Lactobacillus casei microencapsulation via the Maillard reaction between soy protein isolate and prebiotic oligosaccharides. Journal of Agricultural and Food Chemistry 69(46): 13744–13753. 10.1021/acs.jafc.1c02907
Zommiti, M., Feuilloley, M.G.J., and Connil, N. 2020. Update of probiotics in human world: a nonstop source of benefactions till the end of time. Microorganisms 8(12): 1907. 10.3390/microorganisms8121907