
Download
Cell viability ( % ) = mean OD treatment mean OD control × 100 Δ W(%) = I w − F iw I w × 100 Δ E = ( Δ L * ) 2 + ( Δ a * ) 2 + ( Δ b * ) 2


∀ t > t l a g → d N ( t ) d t > 0 , d N ( t ) d t = K 1 ⋅ ( t − t l a g ) 1 + ( t − t l a g ) K 2 ⋅ N ( t ) d ln ( N ( t ) ) d t = K 1 ⋅ ( t − t l a g ) 1 + ( t − t l a g ) K 2 d ( l o g N ( t ) ) d t = K ′ 1 ⋅ ( t − t l a g ) 1 + ( t − t l a g ) K 2




RESEARCH ARTICLE
Improving the shelf life of fresh pasta filata cheese using lactobacilli postbiotics and carbon dots
Elahe Divsalar1, Mehran Moradi1*, Hossein Tajik1*, Rahim Molaei1,2, Amalia Conte3, MAtteo Alessandro Del Nobile4
1Department of Food Hygiene and Quality Control, Faculty of Veterinary Medicine, Urmia University, Urmia, Iran;
2Materials Synthesis Laboratory, Carbon Tech Industrial Group, Carbon Tech, Urmia, Iran;
3Department of Humanistic Studies. Letters, Cultural Heritage, Educational Sciences, University of Foggia, Via Arpi, 176 – 71122 Foggia, Italy;
4Department of Economics, Management and Territory, Via da Zara, 4 – 71121 Foggia, Italy
Abstract
This study investigated the efficacy of aerosolized postbiotics derived from Lactobacillus mesenteroides, zinc-carbon dots (Z-CDs), and their combined application in extending the shelf life of fresh pasta filata cheese (Z-CDs-P). The postbiotics (300 mg/mL), Z-CDs (5 mg/mL), and their combination were used in fresh pasta filata cheese during storage at 4 °C for 18 days by the aerosolization method. A diverse range of microbial and fungal species were monitored in cheese samples. A mathematical model was used to estimate the shelf life of cheese by analyzing both microbiological and sensory data. Based on the results, the control cheese remained acceptable for 5.5 days. Both Z-CDs and Z-CDs420nm treatments slightly extended the shelf life by 1–2 days. The addition of postbiotics further increased the cheese storability, and their combination with Z-CDs and Z-CDs420nm prolonged the shelf life to over 9 and 11 days, respectively. These findings demonstrated the potential of Z-CDs and postbiotics as a promising strategy for extending the shelf life of fresh cheese. Because of its multifunctional properties, the Z-CDs-P solution delivered via aerosolized spray emerges as a promising candidate for applications in dairy industry.
Key words: Active packaging, carbon quantum dots, cell supernatant, photodynamic, whey
*Corresponding Authors: Mehran Moradi, Department of Food Hygiene and Quality Control, Faculty of Veterinary Medicine, Urmia University, Urmia, Iran. Emails: [email protected], [email protected]; Hossein Tajik, Department of Food Hygiene and Quality Control, Faculty of Veterinary Medicine, Urmia University, Urmia, Iran. E-mail: [email protected]; Amalia Conte, Department of Humanistic Studies. Letters, Cultural Heritage, Educational Sciences, University of Foggia, Via Arpi, 176 – 71122 Foggia, Italy. Email: [email protected]
Academic Editor: Yuthana Phimolsiripol, PhD., Division of Product Development Technology, Faculty of Agro-Industry, Chiang Mai University, Thailand
Received: 5 August 2024; Accepted: 2 January 2025; Published: 1 April 2025
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Fresh pasta filata cheese is an Italian cheese made from cow’s or buffalo’s milk. It is usually preserved in tap water, dilute salt solution, or whey solution (Gorrasi et al., 2016). It has a high moisture content (50–60%), high-fat content, high water activity, and slightly acidic pH (5.2 to 5.6) (Torrijos et al., 2022), which reduces its shelf life to 5–7 days at 4 °C (Azhdari and Moradi, 2022). A high population of non-pathogenic lactic acid bacteria (LAB), coliform bacteria, Pseudomonas spp., and other psychrophilic bacteria are the main spoilage microorganisms of cheese. These bacteria release lipases and proteases that affect texture, color, and odor of cheese. Not all microorganisms responsible for the changes in fresh cheese originate from milk; the main source of contamination during cheese processing is the water used in the cooling phase. Coliform and Pseudomonas spp. bacteria frequently contaminate circulating cooling water used in dairies (Hebishy et al., 2024; Segat et al., 2014). Various preservation techniques, including modified atmosphere packaging (Alam and Goyal, 2011; Deshwal et al., 2021), vacuum packaging (Akarca et al., 2015), edible coating (Torrijos et al., 2022), and active systems (Lacivita et al., 2023), are employed to enhance the quality and shelf life of fresh pasta filata cheese.
Different strategies such as film wrapping, dipping, coating, and spraying have been reported for using active agents for antimicrobial packaging of cheese (Costa et al., 2018). Spraying is a common technique to apply a coating solution onto a substrate in laboratory and industrial settings. Aerosolization, a spray method, disperses a liquid phase into tiny droplets or mist (average diameter greater than 5 μm) using conventional spray, electrostatic spray, or commercial nebulizers. Commercial nebulizers are commonly employed for aerosol production, using Bernoulli’s principle-based atomizer nebulizers model (Mukherjee et al., 2017; Sullivan et al., 2020). The characteristics of the formed layer depend on various factors like process parameters, solution properties, and spray dispersion design (Fu and Dudley, 2021). One of the main benefits of spraying is the ability to create a uniform, thin coating layer with precise thickness control. Using spraying devices could minimize secondary contamination in food products (Sharafi et al., 2024). The commercial nebulizer is a widely used aerosol production system that efficiently delivers antimicrobial substances in fine droplets. It is based on Bernoulli’s principle and offers advantages like ease of application and enhanced antimicrobial penetration. Conventional spray methods may have limitations in control and coverage. Both commercial synthetic antimicrobials (Lee et al., 2021) and natural active solutions have been applied using aerosol methods with commercial nebulizers and conventional spray on hake fish fillets (Sullivan et al., 2020) and mozzarella cheese (Torrijos et al., 2022).
Postbiotics (P) are beneficial by-products of probiotics that can be produced in culture media, foods, or digestive tract (Teame et al., 2020). They contain complex combinations of functional and therapeutic substances such as organic acids, exopolysaccharides (EPS), bacteriocins, bioactive peptides, enzymes, and other components that justify the health and bioactive effects of probiotic (Hosseini et al., 2024; Sharafi et al., 2024). Unlike probiotics, postbiotics are nonliving substances that offer similar health benefits, without risk of infection or overgrowth. They are also more convenient to consume, as they do not require refrigeration or special storage (Moradi et al., 2021b). Postbiotics are also more tolerant to pH changes and can be included in a wide range of foods and beverages (Barros et al., 2020). One of the most important groups of postbiotic compounds is that of organic acids, such as lactic acid, benzoic acid, citric acid, etc., that affect membrane integrity by decreasing the intracellular pH. Therefore, postbiotics can remain stable at different pH due to their acidic nature (Hosseini et al., 2022). Peptides with antimicrobial properties in postbiotics are water-soluble and active over a wide pH range, which could allow their application in a wide variety of food products (Aguilar-Toalá et al., 2018). Leuconostoc spp., commonly used in the food industry, is known for its ability to inhibit the growth of spoilage and pathogenic bacteria. Its postbiotics contain various antimicrobial compounds, including organic acids, hydrogen peroxide, and bacteriocins (Raman et al., 2022). Apart from producing antimicrobial compounds, Leuconostoc spp. also produces different types of EPS, such as dextran, levan, and glucan. These EPS have been shown to have immunomodulatory, antioxidant, and anticancer properties (Bisson et al., 2023; Matsuzaki et al., 2021). However, certain production methods can impact the functionality of postbiotics. For instance, spray drying may lead to a loss of volatile metabolites, while lyophilization can remove hydrogen peroxide (Moradi et al., 2021b). Experimental observations suggest that lyophilized postbiotics derived from specific LAB strains may have significantly reduced antioxidant capacity compared to the original solution. Powdered postbiotics exhibit greater antimicrobial activity, but processes like lyophilization can diminish the antibacterial effects of substances like hydrogen peroxide. Consequently, the antimicrobial activity of lyophilized postbiotics appears to be primarily associated with the organic acids, as well as the presence of bacteriocins (Sharafi et al., 2024).
Carbon dots (CDs), with their exceptionally small size of less than 10 nm, emerge as a compelling alternative to traditional antimicrobial chemicals in active packaging (Moradi et al., 2023). Their antimicrobial effect against bacterial cells is because of their ultra-small size, core structure, functional groups on the surface, optical properties, and factors related to the microbial species (Dong et al., 2020). The antimicrobial mechanisms of CDs are caused by physical and mechanical damage to the bacterial membrane/cell, reactive oxygen species (ROS) generation, photocatalytic activity, and cytoplasmic leakage (Divsalar et al., 2023; Esmaeili Koutamehr et al., 2023; Lacivita et al., 2023). Previously, the antimicrobial potential of CDs has been shown on different bacterial species, including Gram-positive and Gram-negative bacteria (Esmaeili Koutamehr et al., 2023; Ghorbani et al., 2022; Varghese and Balachandran, 2021). CDs have been used as photosensitizers (PS) in the photodynamic inactivation of microorganisms (PDI) which results in the termination of bacterial growth by damaging DNA or protein synthesis (Divsalar et al., 2023; Nie et al., 2020). The mechanism of PDI involves a combination of a nontoxic PS and a suitable light source, which selectively destroys microbial cells by producing ROS (Romero et al., 2021). CDs offer effective photodynamic action because of their nontoxic, stable, and high quantum performance (Nie et al., 2020; Romero et al., 2021). Doped CDs are produced by introducing metallic or nonmetallic impurities into the matrix of CDs (Miao et al., 2020) to improve and adjust the intrinsic characteristics of CDs, such as fluorescence efficiency, catalytic activity, and functional properties, also including antimicrobial and antioxidant performances (Li et al., 2019). Zinc can be used as a doping agent in the matrix of CDs to improve optical performance, increase photocatalytic activity, and enhance antibacterial characteristics (Divsalar et al., 2023). CDs are increasingly being explored as active antimicrobial agents in food packaging through aerosolization because of their small size and dispersibility in aqueous solutions (Sullivan et al., 2020), even though applications to dairy sector are very limited.
This research used for the first time an aerosol delivery system of CDs for a dairy application, also combining the adoption of postbiotics. Notably, the postbiotics solution served as diluent and carrier for Z-CDs during the aerosolization process. The system works as a nebulizer with several advantages, as readily available, simple application, broad surface coverage, production of fine mist droplets, and superior antimicrobial penetration into cheese compared to alternative spray methods. Initially, we comprehensively evaluated the cytotoxic properties of postbiotics derived from L. mesenteroides prepared from ultrafiltered (UF) cheese whey. Quasi-spherical (having some spherical characteristics with a high-density core surrounded by a relatively spherical core) Z-CDs with well-established characteristics and white appearance were synthesized from zinc acetate, based on previous work of Divsalar et al. (2023). Postbiotics (P) and Z-CDs were applied individually and in combination (Z-CDs-P) using the aerosolization technique to fresh pasta filata cheese. While a diverse range of microbes were monitored, only data for the two main spoilage groups, coliforms and Pseudomonas spp., were fitted to a mathematical model, as these groups are the primary determinants of the acceptability of cheese. In addition, changes in sensory quality, pH, weight loss, and color were monitored throughout the storage.
Materials and Methods
Materials
L. mesenteroides (ATCC 10830) was obtained from the Iranian Research Organization for Science and Technology (IROST), Tehran, Iran. UF cheese whey was kindly provided by Pegah dairy factory, Urmia, Iran. All culture media and dilution solutions were obtained from Quelab (Montreal, Canada). Sterile Polytetrafluoroethylene (PTFE) syringe filters were obtained from Millipore Inc (Massachusetts, USA); ethanol from Pars Alcohol, Shiraz, Iran; and other chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA). Light emitting diode (LED420nm) lamps were also purchased from Saba Light (Tehran, Iran).
Preparation of postbiotics
The most used medium for LAB fermentation is De Man, Rogosa, and Sharpe broth (MRS), which has been successfully used in the production of postbiotics for various purposes. Nevertheless, the yellow–brown color of the postbiotics produced in the MRS medium may limit their potential use in food-related applications such as dairy products (Sharafi et al., 2024). So, we selected UF whey solution as a medium for the preparation of white-colored postbiotics. To the aim, L. mesenteroides was grown in MRS broth supplemented with 2% sucrose at 28 °C for 24 h. Bacterial pellets were separated by centrifugation (4,000 × g for 15 min), washed twice with normal saline solution, and resuspended in sterile peptone water (0.1%). After 10-fold serial dilutions, the suspensions were cultured, and the dilution containing 1010 CFU/mL was selected for further analysis (Esmaeilnejad-Moghadam et al., 2019). To prepare postbiotics, the pH of UF cheese whey solution was set to 4.5 with 5N hydrochloric acid, autoclaved (121 °C for 20 min) to remove protein precipitates, and centrifuged to separate the supernatant. Next, yeast extract (1% w/v) was added, and the bacterial suspension (1010 CFU/mL) was added to 99 mL of whey medium. The suspensions were incubated at 28 °C for 24 h, and the postbiotic solution was collected by centrifugation at 4,000 × g for 15 min and sterilized using a syringe filter (0.22 µm). Finally, the postbiotic solutions were freeze-dried and stored at −18 °C for future use (Esmaeilnejad-Moghadam et al., 2019; Sharafi et al., 2022).
Chemical composition of postbiotics
Postbiotic chemical compounds were identified using a gas chromatography-mass spectrometer (GC–MS) GC-7890B/mass detectors MS-5975C model (Agilent Technologies, Santa Clara, CA, USA) connected to an electron ionization mass spectrometer. The samples were mixed with ethyl acetate at a ratio of 1:2, and after separation by the separatory funnel, they were concentrated using a nitrogen pump. A HP-5MS column (30 m in length, 0.25 mm in diameter, and 0.25 μm film thickness) was used for separation, and helium gas with a purity of 99.99% was used as the carrier gas with a linear flow rate of 0.9 mL/min (Mohammadi et al., 2022).
Cytotoxicity of postbiotics
The cytotoxicity of postbiotics was investigated in a mouse fibroblast cell line (L929) using a standard MTT assay (Muktha et al., 2021; Sarajar et al., 2023). Measurement of cell viability and proliferation is crucial for the in vitro assays responding to external factors. The yellow tetrazolium MTT (3-(4, 5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide) is reduced by active cells through dehydrogenase enzymes, producing purple formazan quantifiable by spectrophotometry. The MTT assay reflects cell proliferation rates and responds to apoptosis or necrosis, while minimizing assay steps for efficiency. A linear relationship between cell number and signal enables accurate quantification of proliferation changes. Cell viability was calculated using Equation 1:
where ODtreatment and ODcontrol represent the optical density of cells exposed to postbiotics and optical density of cells alone, respectively.
Z-CDs synthesis
In a previous study (Divsalar et al., 2023), Z-CDs were synthesized hydrothermally using zinc-acetate solution as the doping agent and carbon source. This was accomplished by preparing a zinc acetate solution (1%), pouring it into a hydrothermal reactor (300 mL), and heating it for 6 h at 200 °C. The resulting cooled solution was centrifuged (15,000 rpm for 30 min) and filtered via a sterile PTFE syringe filter (0.45 μm) to remove impurities. Z-CDs solution was freeze-dried and kept in dark and sterile conditions at 4 °C for later use. A white Z-CDs powder that showed no cytotoxicity in the L929 cell line at concentrations below 5 mg/mL was synthesized (Divsalar et al., 2023).
Cheese samples treatments
Fresh pasta filata cheese was purchased from the Calin Dairy Company (Tehran, Iran) and transported to the laboratory under cold conditions. Based on the microbroth method, postbiotics MIC/MBC was 30 mg/mL. Also, postbiotic antimicrobial properties at concentrations of 150, 300, and 450 mg/mL (based on 5 MIC, 10 MIC and 15 MIC) were performed by agar well diffusion. The average zones of inhibition (ZOI) were recorded as 18.34±0.31 mm, 22.32±0.48 mm, and 23.35±0.52 mm, respectively. Since 300 and 450 mg/mL were not significant based on the statistical analysis, due to the nontoxicity of these concentrations and due to the pilot tests conducted in the cheese model, the concentration of 300 mg/mL was used (unpublished data). According to the previous study, the toxicity effects of Z-CDs were investigated at 0.1, 0.5, 1, 5, and 10 mg/mL concentrations, and the results showed that Z-CDs was not toxic in L929 cell line at a concentration of 5 mg/mL. So, this concentration was used for cheese application (Divsalar et al., 2023). The evaluation of the toxicity effects of the combination of postbiotics and CDs were not carried out because requires special investigations. Cheese samples were divided into six groups as follows: (a) cheese sample (control), (b) cheese sample with Z-CDs (5 mg/mL), (c) cheese sample with Z-CDs (5 mg/mL) at a ratio of 2:1 under LED420nm radiation, (d) cheese sample with postbiotics (300 mg/mL), (e) cheese sample with a combination of postbiotics (300 mg/mL) and Z-CDs (5 mg/mL) at a ratio of 2:1, and (f) cheese sample with a combination of postbiotics (300 mg/mL) and Z-CDs (5 mg/mL) under LED420nm radiation at a ratio of 2:1. In a separate experiment, we determined the Z-CDs and postbiotics concentrations and their combination ratios suitable for cheese application. A postbiotic solution was used as a carrier solution for aerosolization. Next, 10 mL of each solution was carefully poured into the tank of an atomizer nebulizer (3A, Lonato del Garda, Italy). The nebulizer was then operated at a speed of 0.35 mL/min for 30 min, ensuring that all parts of the cheese were evenly sprayed. The aerosolization process took place in a chamber with a volume of 7.5 liters, measuring 26.5 cm in length, 21 cm in width, and 17.5 cm in height. The chamber had inlet and outlet with diameter of 2 cm, which were 13 and 2 cm from the chamber bottom, respectively. Fresh pasta filata cheese balls were placed on a sterile perforated grill tray which was 10 cm from the bottom of the chamber and 1 cm from the inlet. Upon completion of the aerosolization process, 50 g of each cheese sample was individually packaged by the overwrapped tray method in a polyethylene box and stored at 4 °C for 18 days. Every 3 days, microbial counts, color, pH, and sensory properties of cheese samples were analyzed (Lacivita et al., 2023; Lucera et al., 2014; Moradi et al., 2021a).
Microbiological analyses
Five grams of each sample were carefully weighed and added to 45 mL of peptone water (0.1%). The mixture was then homogenized using a stomacher for 2 min at 260 rpm to ensure uniform distribution of the microbial cells. Subsequently, 10-fold serial dilutions were prepared from homogenates. Aliquots of each dilution were then plated onto appropriate selective culture media for the following tests: Coliforms, Pseudomonas spp., total mesophilic count (TMC), total psychrotrophic count (TPC), yeast–mold count (YMC), and LAB bacilli count (Lacivita et al., 2023; Sharafi et al., 2022).
pH
Cheese samples were accurately weighed and analyzed using a digital pH meter (Hanna instruments, Lisbon, Portugal) directly in the cheese.
Weight loss
Using a high-precision balance, the weight of the cheese samples was measured at specific storage intervals to estimate weight loss using Equation 2 (Azhdari and Moradi, 2022):
where ΔW, Iw, and Fiw represent the weight loss, primary weight (day 0), and weight at time i (other days), respectively.
Sensory evaluation
Seven trained sensory panelists evaluated the sensory characteristics of cheese samples in terms of color, odor, texture, and overall acceptability using a 10-point hybrid hedonic scale (1 = dislike extremely to 10 = like extremely) (Pimentel et al., 2016). For the analysis, the samples were presented simultaneously. Sensory data were fitted by a modified version of the Gompertz equation (Lacivita et al., 2023) to determine the day within which each sample remained acceptable from a sensory point of view (sensory acceptability limit, SAL value).
Color measurements
The color of the cheese samples (in different levels) was measured using a digital colorimeter (Lovibond, London, UK), and the total color difference (ΔE) of each sample was recorded using the L* (lightness), a* (red–green), and b* (yellow–blue) values, according to the Equation 3 (Sharafi et al., 2022):
Statistical analysis
All the experiments were conducted in triplicates. Statistical analyses were performed using GraphPad Prism version 9.01 (GraphPad Software, USA). Analysis of variance (ANOVA) was performed on all collected data, followed by Duncan’s multiple range test (p < 0.05).
Results and Discussion
Chemical analysis and cytotoxicity of postbiotics
The profile of volatile chemical compounds of the postbiotics was identified using the GC-MS (Table 1). Based on their results, 23 volatile compounds were identified, more than 79% of all compounds. Among the identified compounds, 6-octadecanoic acid methyl ester (Moradi et al., 2021b), 2,3-hydroxypropyl dodecanoic acid ester (Letlole et al., 2021), 4-ethoxy-N-ethyl-N-methyl-Benzenamine (Belhocine et al., 2023; Ottonelli Stopiglia et al., 2012), and benzoic acid (Moradi et al., 2019) with 44.13%, 7.99%, 5.58%, and 3.04% have the highest content, respectively.
Table 1. Metabolic profiling of L. mesenteroides postbiotics using GC-MS.
| No. | Name | Content (%) | Retention time (min) | Expected biological activity | References |
|---|---|---|---|---|---|
| 1 | 4-ethoxy-N-ethyl-N-methyl-Benzenamine | 5.582 | 5.294 | Antibacterial and antifungal | (Belhocine et al., 2023; Ottonelli Stopiglia et al., 2012) |
| 2 | 3-Methoxy-4-[(trimethylsilyl) oxy]-benzaldehyde-o-methyloxime | 1.633 | 11.731 | Antibacterial | (Lingfa and Ankanagari, 2023) |
| 3 | Methyl t-butyl ether | 2.005 | 15.330 | Antimicrobial and antioxidant | (d'Errico et al., 2020; Yu et al., 2013) |
| 4 | Benzoic acid | 3.047 | 17.642 | Antibacterial and antioxidant | (Surowiak et al., 2020) |
| 5 | Benzaldehyde, 3,5-dimethyl | 0.931 | 18.386 | Antibacterial, antifungal, and antioxidant | (Wajda-Hermanowicz et al., 2015) |
| 6 | 6-Octadecenoic acid methyl ester | 44.131 | 22.346 | Antioxidant and antimicrobial | (Moradi et al., 2021b) |
| 7 | 2,4-bis(1,1-dimethylethyl) Phenol | 0.414 | 30.934 | Antioxidant, antibacterial, and antifungal | (Padmavathi et al., 2014) |
| 8 | Dodecanoic acid | 0.658 | 31.183 | Antibacterial, antifungal, and antioxidant | (Kubo et al., 2008) |
| 9 | Nonadecane | 0.326 | 34.224 | Antibacterial, antifungal, and antioxidant | (El-Hawary et al., 2018) |
| 10 | Tetradecanoic acid | 0.408 | 40.307 | Antibacterial, antifungal, and antioxidant | (Sivakumar et al., 2011) |
| 11 | Nonadecene | 0.292 | 41.411 | Antibacterial, antifungal, and antioxidant | (El-Hawary et al., 2018) |
| 12 | 1,4-diaza-2,5-dioxo-3-isobutyl bicyclo[4.3.0]nonane | 0.891 | 45.371 | Antibiofilm | (Nadar Rajivgandhi et al., 2020) |
| 13 | Hexadecanoic acid methyl ester | 0.605 | 45.645 | Antimicrobial, antioxidant, and anti-inflammatory | (Shaaban et al., 2021) |
| 14 | Pyrrolo[1,2-a]pyrazine-1,4-dione hexahydro-3-(2-methylpropyl) | 2.208 | 45.914 | Antioxidant | (Law et al., 2019) |
| 15 | Hexadecanoic acid | 2.575 | 46.967 | Antimicrobial, antioxidant, and anti-inflammatory | (Chen et al., 2010) |
| 16 | Eicosane | 0.302 | 47.946 | Antibacterial and antifungal | (Rhetso et al., 2020) |
| 17 | 2,3-dihydroxypropylester Dodecanoic acid | 7.993 | 50.892 | Antiviral and antibacterial | (Letlole et al., 2021) |
| 18 | Oleic acid | 1.440 | 52.323 | Antibacterial, antitumor, and antifungal | (Verma et al., 2014) |
| 19 | Docosane | 0.668 | 53.937 | Antibacterial, antifungal, and antioxidant | (Perveen et al., 2022) |
| 20 | Hexatriacontane | 0.866 | 56.746 | Antimicrobial and antioxidant | (Aodah et al., 2023) |
| 21 | Octacosane | 0.889 | 59.447 | Antibacterial, antifungal, and antioxidant | (Rhetso et al., 2020) |
| 22 | Bis (2 -ethylhexyl ) phthalate | 0.840 | 63.258 | Antibacterial, antifungal, and antioxidant | (Lotfy et al., 2018) |
| 23 | Pentatriacontane | 0.486 | 64.551 | Antibacterial | (Rehman et al., 2021) |
LAB and their metabolites are known to be safe for long-term use in food fermentation processes (Obis et al., 2019). The use of postbiotics as food additives requires regulatory approval, establishing permissible amounts for safe consumption. In addition, toxicity studies are crucial to fully assess potential risks associated with postbiotics (Mani-López et al., 2022). The cytotoxicity of different concentrations (0.5, 50, 100, 250, and 500 mg/mL) of postbiotics were assessed on L929 cell line after 24 h of exposure. As shown in Figure 1, the safety range of 0.5–500 mg/mL was observed on the L929 cell line compared to control, which indicates the low cytotoxic potential of postbiotics for food systems.
Figure 1. Viability (%) and L929 cells images in the presence of postbiotics during 24 h exposure time. Values with different letters are significantly different (p < 0.05) from the control sample.
Pasta filata cheese shelf life
Various treatments were applied to fresh pasta filata cheese (moisture 47%, fat 20%, and NaCl 1.5%) to evaluate their impact on the microbial quality decay kinetics. To this aim, a diverse range of microbial and fungal species were monitored during storage. However, only the experimental data of two spoilage groups, coliforms and Pseudomonas spp., which are the primary determinants of cheese acceptability, were effectively fitted by a mathematical model. Variations in pH, weight loss, sensory quality and color were also evaluated during the refrigerated storage time.
Effects of treatments on microbial quality decay kinetic
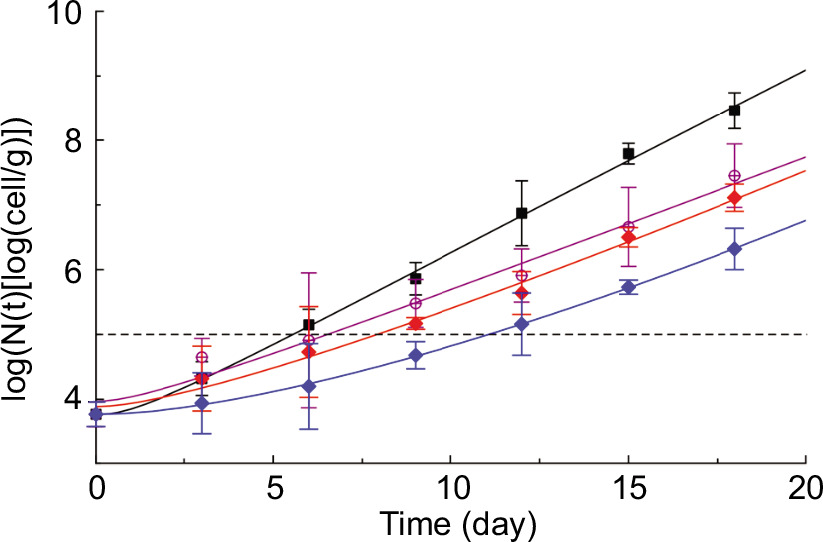
Figure 2 depicts the temporal evolution of viable coliform cell counts during refrigerated storage. For reasons of clarity, the coliform bacteria growth kinetics of only a few of the analyzed samples are presented in Figure 2. The horizontal dashed line represents the coliform threshold (5 log10 CFU/g), signifying the maximum permitted coliform concentration in mozzarella-like fresh cheese (Conte et al., 2007). As anticipated, the coliform bacteria viable cell concentration steadily increased across the entire storage period for all the investigated samples. Figure 2 also highlights a pronounced impact of the applied treatments on coliform bacteria growth kinetics. The time required to surpass the threshold varies between 5 and 11 days. Notably, the control sample attained the threshold considerably faster than the other samples, suggesting that all the treatments investigated effectively retarded coliform bacteria growth kinetics. In addition, it is apparent that the time needed for the other samples to reach the threshold differs from sample to sample. In particular, the order in which microbial growth kinetics slows down is as follows: Z-CDs, Z-CDs420nm, P, Z-CDs-P, Z-CDs420nm-P, with Z-CDs exhibiting the fastest and Z-CDs420nm-P displaying the slowest effect. The observed differences among the samples cannot be attributed solely to their lag phase, as this phase does not appear to be significantly influenced by the type of treatment applied. Data presented in Figure 2 suggest that the observed differences in the growth kinetics are primarily attributable to variations in their specific growth rate (SGR). The growth of coliforms was delayed in samples containing CDs. Alginate-based active-coated fiordilatte cheese with CDs at a concentration of 5,000 mg/L maintained coliform bacteria counts below 5 log10 CFU/g for up to 15 days, while the control cheese exceeded 6 log10 CFU/g after 10 days of storage (Lacivita et al., 2023). Diverse concentrations of lemon extract (500, 1,000, and 1,500 ppm) were used as active agents in conjunction with brine and sodium alginate solution to preserve mozzarella cheese (Conte et al., 2007). Control samples exhibited significantly higher coliform counts compared to treated samples. Notably, the active solution system proved to be more effective in reducing the rate of microbial growth compared to the active gel system. The presence of lemon extract effectively restrained coliform growth during the initial stage of storage, reaching 6.5 log10 CFU/g after 10 days, while the control sample reached 8 log10 CFU/g (Conte et al., 2007).
Figure 2. Time course during refrigerated storage (4 °C) of the concentration of viable coliform counts. The curves shown in the figure have been obtained by fitting Equation 7 to the experimental data. Control cheese (black); cheese treated by Z-CDs (violet); P (red), and Z-CDs420nm-P (blue).
Figure 3 shows the Pseudomonas spp. viable cell counts plotted as a function of storage time. As was done previously, to enhance the readability of the figure, only a subset of the samples evaluated in this study are displayed. The dashed horizontal line shown represents the threshold for Pseudomonas spp. bacteria, set at 6 log10 CFU/g, as reported in a previous paper also dealing with mozzarella cheese (Conte et al., 2007); when the Pseudomonas spp. viable cell counts exceeds this threshold, the product is no longer acceptable because of the noticeable sensory defects. As observed for coliforms, the Pseudomonas spp. viable cell counts progressively increased during the entire period of observation for all the samples. The treatments used greatly affected the Pseudomonas spp. growth kinetics, as they did with coliforms. Consequently, the time at which the cheese become unacceptable (i.e., the time at which the Pseudomonas spp. threshold is reached) is strongly affected by the treatments. It has been observed that all the preservation strategies used in this study slowed down the growth kinetics of this spoilage group. Moreover, important differences among them have been also demonstrated. Specifically, it was observed that the microbial growth kinetics differ from each other both in terms of lag phase and SGR. In fact, unlike what was observed in the case of coliforms, for Pseudomonas spp., in addition to a slowdown in growth kinetics, a variation in the shape of the microbial growth curve is also observed. Similar trends to those observed for coliforms were observed regarding the time required to reach the Pseudomonas spp. threshold. In particular, the order in which the treated samples reached the threshold is as follows: Z-CDs, Z-CDs420nm, P, Z-CDs-P, Z-CDs420nm-P; Z-CDs being the fastest and Z-CDs420nm-P being the slowest.
Figure 3. Time course during refrigerated storage (4 °C) of the concentration of viable Pseudomonas spp. counts. The curves shown in the figure have been obtained by fitting Equation 7 to the experimental data. Control cheese (black); cheese treated by Z-CDs420nm (green); Z-CDs-P (sky-blue); and Z-CDs420nm-P (blue).
The population of Pseudomonas spp. bacteria in the control showed a concentration beyond the permissible limit (6 log10 CFU/g after the third day). It took slightly more than 6 days for samples to become unacceptable in treatments containing CDs 2,500 mg/L, while the other treatments needed more time to reach unacceptable levels (more than a week) and had slower growth. When the CDs were directly dissolved in brine at a concentration of 5,000 mg/L, the microbial inhibition was more obvious, and the samples reached the spoilage threshold within 10 days. It is worth noting that the worst sample was the control, as it was microbiologically unacceptable in the first few days (Lacivita et al., 2023).
In terms of the preservability of fresh pasta filata cheese, the primary concern is the rapidity with which the microbial load reaches its threshold. Keeping this in mind, a simple model was used to quantitatively describe the microbial growth kinetics obtained in this study and to determine the time at which the viable cell concentration reached its threshold (i.e., the microbial acceptability limit—MAL).
The model is based on two hypotheses: (a) the growth kinetic follows a first-order kinetic type equation with a time-dependent kinetic constant and (b)
where N(t) is the viable cell concentration at time t, and tlag is the lag time.
The following equation is proposed to describe the time course of the viable cell concentration ∀t > tlag:
Which can be rearranged as:
where Ki are the model parameters, which are assumed to be constant. It is worth noting that the term
where .
The curves shown in Figures 2 and 3 have been obtained by fitting Equation 7 to the experimental data. As can be inferred from the above-mentioned Figures, despite its simplicity, the proposed model adequately interpolates the experimental data. The model parameters obtained from the fitting procedure were employed to estimate the time required for each microbial species to reach the threshold (MAL). The obtained results are listed in Table 2. The same table also shows the values of the percentage increase in MAL compared to the control (Δ%MALColi and Δ%MALPse).
Table 2. Values of microbial acceptability limit (MAL) and 0%MAL for coliforms and Pseudomonas spp. in fresh cheese treated with Zinc carbon dots (Z-CDs) and postbiotics (P) and untreated fresh cheese (control).
| Treatment | MALPse (day) | VKMALPse | MALColi (day) | MiMAl™ |
|---|---|---|---|---|
| Control | 9.43 | - | 5.55 | - |
| Z-CDs | 10.27 | 9 | 6.52 | 17 |
| Z-CDs420nm | 10.93 | 16 | 7.05 | 27 |
| P | 12.30 | 30 | 7.91 | 42 |
| Z-CDs-P | 13.40 | 42 | 9.40 | 69 |
| Z-CDs420nm-P | 14.96 | 59 | 11.07 | 99 |
MAL values represent fitting parameters of Equation 7 to the experimental data . N%MALColi and N%MALPse are the percentage increase in MAL compared to the control, to better highlight the difference between control and active samples.
Based on the MAL values obtained for coliforms (MALColi), it is evident that the investigated treatments have a significant impact. For instance, the MALColi of sample Z-CDs420nm-P is twice that of the control sample. All the treated samples show a MALColi greater than that of the control sample. The MALColi increases according to the following order: Z-CDs, Z-CDs420nm, P, Z-CDs-P, and Z-CDs420nm-P. However, samples Z-CDs and Z-CDs420nm show a modest increase in MALColi, and Δ%MALColi equal to 17% and 27% were observed, respectively. On the contrary, the use of postbiotics seems to cause a nonnegligible increase in (Δ%MALColi = 42%). Moreover, postbiotics seems to have a catalytic effect on both Z-CDs and Z-CDs420nm treatments. In fact, when used in combination with postbiotics, the MALColi of Z-CDs and Z-CDs420nm increases noticeably; Δ%MALColi of 69% and 99% were observed for Z-CDs-P and Z-CDs420nm-P, respectively. As previously reported, the differences observed in MALColi values do not appear to be because of differences in the lag time; rather they can be ascribed to differences in the SGR.
Regarding Pseudomonas spp., the MAL values (MALPse) presented in Table 2 show a trend similar to that observed for coliforms, reinforcing the idea that the addition of postbiotics further enhanced the effectiveness of both Z-CDs and Z-CDs420nm.However, in the case of Pseudomonas spp., the Δ%MALPse are not as large as those found for coliforms, suggesting that the studied treatments were not as effective on Pseudomonas spp. as they were on coliforms. This is not surprising, as the effectiveness of the given antimicrobial treatment strongly depends on the target microorganism (Ezati et al., 2022; Zhao et al., 2022).
Figure 4A shows the time course during storage of the coliform-SGR (SGRColi). Equation 5 was used to the calculate the curves shown in Figure 4 (a); the values of K1, K2, and tlag are those obtained by fitting Equation 7 to the coliform growth kinetic data shown in Figure 2. It is useful to note that to understand how the SGRColi affects the MALColi of a specific sample, it is necessary to limit oneself to seeing the SGRColi data until the MALColi is reached. As one would expect, the lag time of all the samples is close to zero; therefore, it cannot be considered responsible for the observed difference in the MALColi values. On the other hand, Figure 4 (a) shows that the SGRColi greatly changed from one sample to another, thus supporting the idea that the observed differences in the MALColi must be attributed to differences in the SGR.
Figure 4. Time course during storage at 4 °C of the coliform (A) and Pseudomonas spp. (B) specific growth rate. Control cheese (black); cheese treated by Z-CDs (violet); Z-CDs420nm (green); P (red); Z-CDs-P (sky-blue); and Z-CDs420nm-P (blue).
Figure 4B shows the time course during storage of the Pseudomonas spp. SGR (SGRPse). Also, for Pseudomonas spp., Equation 6 was used to calculate the curves shown in figure 4, and the values of K1, K1, and tlag were those obtained by fitting Equation 7 to the Pseudomonas spp. growth kinetic data shown in Figure 3. Contrary to what was previously observed for coliforms, Pseudomonas spp. show a marked effect of treatments on both the lag time and on the SGRPse. As can be inferred for Figure 4, the tlag of the control sample is zero, whereas that of the treated samples is close to 5 days. Moreover, from what is reported in Figure 4, it is evident that the treatments alter the entire SGRPse curve, if compared to the control sample. While the cheese without any treatment did not reach its maximum SGR, all treated samples reached their maximum SGR at about 6 days. From what is reported in figure 4, it is not possible to establish between tlag and SGRPse as to which one is the most important in determining the increase in the MALPse. Both tlag and SGRPse contributed to increase the MALPse of treated samples, if compared to the control.
Regarding mesophilic and psychrotrophic bacteria, very similar trends were recorded. Figure 5 reports the evolutions of these spoilage groups for control and treated cheese samples. In both cases, while the bacteria in the control product rapidly increased and reached high concentrations around 8 log10 CFU/g, in the treated cheese an important growth delay and less proliferations were detected. Specifically, regardless the treatment, the mesophilic bacteria started to grow after about 5 days of lag phase and reached concentrations less than 7 log10 CFU/g in the 2 weeks of monitoring (Figure 5A). Similarly, microbial counts of psychrotrophic bacteria remained around 4 log10 CFU/g for the first week and then increased around to 6 log10 CFU/g during the storage period. The treatment Z-CDs420nm-P was found to be the most effective against both mesophilic and psychrotrophic bacteria (Figure 5B). Whey-based CDs were used as an antimicrobial agent in fiordillate cheese packaging (Lacivita et al., 2023). CDs (2,500 and 5,000 mg/L) were added through an alginate-based coating or directly to the cheese. The best results were observed for CDs dissolved in brine at a concentration of 5,000 mg/L, which delayed the total mesophilic bacteria population which reached 6 log10 CFU/g on day 15 of storage (Lacivita et al., 2023). Figure 5C reports trends of yeasts and molds in treated and untreated cheese. Also, in this case, it is possible to observe that fungal growth was rapid in the control sample, whereas it was delayed in all the treated products. Except for the control, during the initial days of storage, yeasts and molds remained at very low levels (around 3 log CFU/g) and began to grow only after 3–4 days. Even though the treatments were not effective to completely inhibit fungal proliferation, all the treatments were found to be useful to delay the growth of yeasts and molds. The results of the study by Lacivita et al. (2023) showed that CDs also had interesting effects on yeast populations. Fiordillate cheese with active coating and brine showed a gradual increase in yeast population, which was less than the control groups. After 10 days, 7 log10 CFU/g remained for control, while 6 log10 CFU/g remained at the end of the 15th day in treated samples (Lacivita et al., 2023). As can be observed, LAB in all the samples were more inhibited in the treated cheese (Figure 5D) with Z-CDs-P and Z-CDs420nm-P. In a similar study, LAB counts in control mozzarella samples showed an increasing trend during the storage period, while in postbiotic-treated cheese, they increased between day 8 and 16. Compared to the control, they inhibited LAB by approximately 1 to 1.3 log10 CFU/g, because of the organic acid content of the Lactobacillus acidophilus LA-5 and Bifidobacterium animalis BB-12 postbiotics (Sharafi et al., 2022).
Figure 5. Time course during refrigerated storage (4 °C) of counts of viable mesophilic bacteria (A), psychrotrophic bacteria (B), yeast and mold (C), and LAB (D). Control cheese (black); cheese treated by Z-CDs (violet); cheese treated by Z-CDs420nm (green); P (red); Z-CDs-P (sky-blue); and Z-CDs420nm-P (blue).
Effects of treatments on pH and weight loss
During the storage period, all the samples were monitored for pH and weight loss (Figure 6). Figure 6A demonstrates that the pH of the control samples slightly increased after the initial week of storage, moving from approximately 5.3 to about 6.4 at the end of the storage period. By contrast, all treated products exhibited a decline in pH. Specifically, the values shifted from approximately 5.2 to values within the range of 4.5–4.7. This phenomenon can be attributed to the acidic nature of the cheese, the activity of LAB, and the presence of postbiotic compounds that have a low pH. In addition, the abundance of LAB in cheese is high, and with the extended storage time, these bacteria proliferate in the control sample. However, pH does not decrease and even slightly increases, which might be attributed to the activity of other microorganisms such as Pseudomonas spp. Their proteolytic enzymes produce amino acids, which raise the pH of the samples (Luengo and Olivera, 2020).
Figure 6. Time course during refrigerated storage (4 °C) of pH (A) and weight loss (B). Control cheese (black); cheese treated by Z-CDs (violet); Z-CDs420nm (green); P (red); Z-CDs-P (sky-blue); and Z-CDs420nm-P (blue).
Regarding weight loss, it is evident that all the treatments had better weight maintenance (Figure 6B). The control cheese experienced a weight loss of approximately 21%, while the weight loss in all the treated samples at the end of storage ranged from 17.8% to 14.3%. Azhdari and Moradi (2022) examined the effects of carboxymethylcellulose coating with and without natamycin on the weight loss of mozzarella cheese during 8 days of storage. In all the samples, weight loss increased linearly over time. It appears that the coating significantly influenced the weight loss of cheese. As a result, on day 8, the uncoated cheese sample exhibited a weight loss of approximately 11.9%, while the reduction in coated cheese was 8.6% (Azhdari and Moradi, 2022).
Effects of treatments on sensory quality
As expected, sensory acceptability of cheese products declined during storage, but significant differences among treated and untreated cheese were recorded (p<0.05). While control became unacceptable after 2 weeks of storage for undesirable changes in odor, color, and texture, all the other treated cheese remained suitable for consumption for the entire monitoring period (data not shown). This means that the sole data of the control cheese were fitted to calculate the SAL. From the fitting procedure (Lacivita et al., 2023), a SAL value equal to 14.88 days was calculated and for the other samples, a SAL value higher than the observation period can be indicated (SAL > 18 days), without difference among the various treatments. The findings recorded in terms of sensory properties confirmed the effects recorded on microbial quality because they demonstrate that the preservation strategies were able to control microbial proliferation and consequently allowed for promoting a better control of the sensory changes. It has been reported that during the storage of cheese, there are some off-flavor volatile compounds, which originate from the activities of cryogenic bacteria such as Pseudomonas spp., and whose content leads to off-flavors in the product (Faccia et al., 2021). One of the characteristics of CDs is their smoky aroma, which is caused by the carboxyl group (COOH) linked to an aromatic ring. The most important effects of smoky odor play a role in increasing sensory acceptance. Using the characteristics of mild pasteurization of radio frequency and the excellent antibacterial and antioxidant properties of CDs, their combination can be used as a new technique to suppress the production of fish-like odor (Zhao et al., 2022). Overall, the evaluators did not find any statistically significant differences in the overall acceptance of cheese stored under different treatments.
Effects of treatments on cheese color
The effect of different treatments on fresh pasta filata cheese color parameters is shown in Table 3. Three parameters including L* (lightness), a* (red–green), and b* (yellow–blue) were used to investigate the color change; finally, by calculating ΔE, the extent of color change developed in different samples compared to the control was calculated (Figure 7). If the difference in ΔE value between two samples is more than 5, the color change is detectable with the naked eye (Sharafi et al., 2022). The difference in ΔE value in the control and in the other treatments from the 12th day onward is more than 5, and it was detectable with the naked eye, and it was significant between the control and the other treatments from the 9th day onward. Also, there was no significant difference among the treatments from day 9 onward. Since the solutions used were colorless, no significant difference was observed between the control and the treatments on days 3 and 6, which is considered as a strong point for the use of these colorless antimicrobial compounds in dairy products. After the sixth day, because of the increase in the microbial growth of the control, a significant difference was observed compared to the other treated cheese. The color change in the samples may be related to the accumulation of postbiotic solutions and Z-CDs on the product surface. The color of postbiotics is related to the whey used to prepare these compounds (Gómez-Narváez et al., 2017). Application of postbiotics prepared in whey produced similar color changes in cheese (Jutinico-Shubach et al., 2020) and fresh beef (Arrioja-Bretón et al., 2020). However, the control sample exhibited more pronounced color changes during storage, which can be likely associated with combined proteolytic and lipolytic activities of psychrophilic bacteria (Marino et al., 2019).
Figure 7. The color difference in fresh pasta filata cheese samples. Values with different letters are significantly different (p ≤ 0.05) from the control sample.
Table 3. Color changes in fresh pasta filata cheese treated with Zinc carbon dots (Z-CDs) and postbiotics (P).
| Treatment | Days | L* | a* | b* |
|---|---|---|---|---|
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 87.46 ± 1.09a | 2.46 ± 1.20a | 11.33 ± 1.62a | |
| 6 | 85.96 ± 2.44a | 1.90 ± 2.26b | 10.86 ± 1.21a | |
| Control | 9 | 82.86 ± 1.21b | -2.96 ± 0.45b | 20.90 ± 2.45a |
| 12 | 80.51 ± 3.81a | -1.30 ± 1.90b | 21.93 ± 1.12a | |
| 15 | 87.86 ± 1.09a | -4.83 ± 0.40a | 20.20 ± 2.44a | |
| 18 | 75.60 ± 2.98d | -4.83 ± 0.35b | 24.26 ± 2.06a | |
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 86.86 ± 1.30a | 1.83 ± 0.20b | 11.06 ± 0.41a | |
| 6 | 85.76 ± 2.10a | -3.30 ± 1.93c | 10.96 ± 0.40a | |
| Z-CDs | 9 | 83.10 ± 2.39b | -2.53 ± 0.55b | 13.43 ± 2.55c |
| 12 | 84.36 ± 2.31b | -1.40 ± 1.83b | 15.30 ± 1.08c | |
| 15 | 80.33 ± 0.85c | -4.73 ± 1.04a | 18.23 ± 0.49b | |
| 18 | 80.20 ± 0.1ac | -4.30 ± 0.65b | 21 ± 1.58b | |
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 86.83 ± 1.19a | 1.50 ± 1.11b | 10.80 ± 0.33a | |
| Z-CDs420nm | 6 | 86.63 ± 0.80a | -4.30 ± 0.34c | 10.86 ± 1.10a |
| 9 | 83.13 ± 1.71b | -3.83 ± 0.92a | 14.40 ± 0.87c | |
| 12 | 82.80 ± 1.66c | -2.53 ± 1.06a | 16.53 ± 1.01c | |
| 15 | 79.43 ± 0.85c | -2.86 ± 1.10b | 18.30 ± 1.17b | |
| 18 | 77.93 ± 2.08b | -4.10 ± 0.98b | 20.93 ± 1.16b | |
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 87.20 ± 1.21a | 1.03 ± 0.45b | 11.10 ± 0.95a | |
| 6 | 86.76 ± 1.96a | 2.40 ± 0.81a | 11.23 ± 0.87a | |
| P | 9 | 84 ± 2.02b | -3.97 ± 0.20a | 13.66 ± 2.76c |
| 12 | 82.70 ± 1.65c | 1.30 ± 0.98b | 18.06 ± 1.26b | |
| 15 | 81.50 ± 2.06b | -5.73 ± 0.50a | 20.30 ± 1.91a | |
| 18 | 77.33 ± 2.55b | -5 ± 0.96b | 21.63 ± 1.30b | |
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 87.33 ± 0.57a | 1.83 ± 0.57b | 10.86 ± 1.41a | |
| 6 | 86.16 ± 2.49a | 1.73 ± 2.05b | 11.23 ± 1.51a | |
| Z-CDs-P | 9 | 84.66 ± 2.08a | -2.93 ± 1.33b | 14.73 ± 1.33b |
| 12 | 83.40 ± 1.57a | -1.50 ± 0.55b | 16.93 ± 2.65c | |
| 15 | 82.83 ± 1.25b | -5.03 ± 0.57a | 18.86 ± 3b | |
| 18 | 81.86 ± 1.21a | -6.18 ± 1.21a | 22.86 ± 1.21b | |
| 0 | 88.56 ± 0.80a | 0.66 ± 0.33a | 12.98 ± 1.10a | |
| 3 | 86.86 ± 1.15a | 1.74 ± 0.68b | 10.60 ± 0.40a | |
| 6 | 85.83 ± 1.01a | -3.63 ± 0.64c | 11.83 ± 0.20a | |
| Z-CDs420nm-P | 9 | 84.63 ± 0.75b | -3.10 ± 0.91a | 15.20 ± 0.45b |
| 12 | 82.83 ± 0.90c | -2.26 ± 0.25a | 15.96 ± 0.85 | |
| 15 | 82.26 ± 1.21b | -2.26 ± 1.62b | 19.76 ± 1.42a | |
| 18 | 81.36 ± 1.70a | -3 ± 0.87c | 20.46 ± 1.29c |
*Within each row, values with different letters are significantly different (p < 0.05).
Shelf life calculation
To calculate the shelf life of the product, both microbial and sensory quality need to be considered. Therefore, according to what is reported in other studies dealing with the shelf life of fresh cheese (Lacivita et al., 2023), comparing the day on which microbial and sensory quality remained acceptable (MAL and SAL). From the microbiological point of view, data of MAL values of coliforms and Pseudomonas spp. (Table 2) need to be compared. From the sensory point of view, the SAL of the control was 14.88 days, whereas the SAL for all the other samples is higher than 18 days, regardless of the treatment applied. Comparing the MAL and SAL values, it was possible to infer that the shelf life of our fresh pasta filata cheese was limited by the coliform proliferation and therefore it coincided with MAL of coliforms. According to these data, the control cheese remained acceptable for about 5.5 days; the treatment with Z-CDs and Z-CDs420nm promoted a slight increase by 1–2 days of shelf life. The addition of postbiotics further increased the storability of cheese and their combination with Z-CDs and with Z-CDs irradiated at 420 nm, further prolonged the shelf life up to more than 9 and 11 days, respectively. Therefore, the results demonstrate that the best combination of Z-CDs420nm-P was able to double the shelf life of the fresh pasta filata cheese, compared to the untreated sample. The combination of postbiotics and Z-CDs had a volume ratio of 2:1 at concentration of 300 mg/mL and 5 mg/mL, respectively. Postbiotics are considered as a Z-CDs solvent. The cell wall of Gram-negative bacteria is made up of a thin layer of peptidoglycan between two cell membranes, phospholipid, and lipopolysaccharide (Ghorbani et al., 2022). Postbiotics from L. mesenteroides, with an acidic nature (pH: 3.5), use organic acids to damage cell membranes and inhibit bacterial growth. Protonated organic acid molecules disrupt the membrane. These molecules also reduce the pH around the cell and exert osmotic pressure on cells (Zhang et al., 2021). Because of the existence of the lipopolysaccharide layer, the Z-CDs cannot directly interact with the peptidoglycan layer but can only interact with certain places in the cell membrane of bacteria. Therefore, postbiotics’ organic acids initially lead to membrane disruption, then Z-CDs exert their antimicrobial effects through the effect of disrupting cellular macromolecules and cellular metabolism, leading to the cell death (Zhao et al., 2019). The synergy between postbiotics and Z-CDs, targeting the cell wall to increase permeability, enhances their effectiveness in destroying bacteria. This dual approach can be promising in combating bacterial pathogens efficiently. The shelf life of fiordilatte cheese containing green CDs was estimated by Lacivita et al. (2023) and according to the results, Pseudomonas spp. primarily affected the shelf life of the product. While the control became unacceptable within 3.5 days, coated samples remained acceptable for twice as long (6 to 7 days). Direct addition of CDs (5000 mg/L) to brine increased shelf life up to 10 days. Compared to other preservation strategies, the use of CDs in brine represents a very promising technique.
Conclusions
Designing food packaging with specific features such as antimicrobial effects without using artificial and chemical preservatives has always been one of the priorities of researchers in the field of food industry. The use of antimicrobial compounds such as postbiotics prepared in whey and white Z-CDs is an innovation for use in dairy products and a challenge for limiting the use of postbiotics based on conventional culture media and black CDs. This study investigated the efficacy of aerosolized Z-CDs, postbiotics derived from L. mesenteroides, and their combined application in extending the shelf life of fresh pasta filata cheese. Postbiotics and Z-CDs were not toxic in L929 fibroblasts at < 500 mg/mL and 5 mg/mL, respectively. The results revealed that individual and combined applications of postbiotics and Z-CD solutions significantly improved the shelf life of fresh pasta filata cheese, with the combined approach proving most effective. All the treatments demonstrated positive impacts on shelf life, with Z-CDs420nm-P exhibiting the most pronounced effect. This treatment achieved reductions in coliforms (2.14 log10 CFU/g), Pseudomonas spp. (2.97 log10 CFU/g), mesophilic bacteria (1.69 log10 CFU/g), psychrotrophic bacteria (2.16 log10 CFU/g), molds–yeasts (1.90 log10 CFU/g), and LAB (log10 CFU/g) during 18 days of storage at 4 °C, compared to the control. Notably, the combination of Z-CDs420nm-P doubled the shelf life of the fresh pasta filata cheese compared to the untreated sample. It is hypothesized that the aromatic and smoky compounds present in Z-CDs penetrated the cheese, imparting a pleasant smoky aroma. Based on the antimicrobial, chemical, and sensory evaluations, these two preservative compounds appear to be suitable additives for fresh cheese. Postbiotics seems to have a catalytic effect on both Z-CDs and Z-CDs420nm treatments. However, challenges such as the scalability of the aerosolization process, cost-effectiveness, and regulatory compliance for the use of CDs in food systems is a complex phenomenon. Future studies should focus on testing these compounds in other food matrices, evaluating long-term storage effects and exploring potential hybrid treatment methods to enhance applicability.
Data Availability Statement
During the preparation of this work, the authors used Paperpal to improve readability and language. After using this service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication. The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank the Faculty of Veterinary Medicine, Urmia University, for funding and technical support.
Author Contributions
Elahe Divsalar was responsible for investigation, formal analysis, and writing—original draft. Mehran Moradi was involved in conceptualization, validation, and writing—review & editing. Hossein Tajik did supervision and visualization. Rahim Molaei performed conceptualization, validation, and writing—review & editing. Amalia Conte was responsible for conceptualization, formal analysis, and writing—review & editing. Alessandro Del Nobile did data curation, supervision and writing–original draft.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
The Authors acknowledge the financial support of Urmia University.
REFERENCES
Aguilar-Toalá, J.E., Garcia-Varela, R., Garcia, H.S., et al., 2018. Postbiotics: An evolving term within the functional foods field. Trends in Food Science & Technology 75: 105–114. 10.1016/j.tifs.2018.03.009
Akarca, G., Tomar, O., and Gök, V., 2015. Effect of different packaging methods on the quality of stuffed and sliced Mozzarella cheese during storage. Journal of Food Processing and Preservation 39(6): 2912–2918. 10.1111/jfpp.12542
Alam, T., and Goyal, G.K., 2011. Effect of MAP on microbiological quality of Mozzarella cheese stored in different packages at 7 ± 1°C. Journal of Food Science and Technology 48(1): 120–123. 10.1007/s13197-010-0141-y
Aodah, A.H., Hashmi, S., Akhtar, N., et al., 2023. Formulation development, optimization by box-behnken design, and in vitro and ex vivo characterization of hexatriacontane-loaded transethosomal gel for antimicrobial treatment for skin infections. Gels 9(4): 322. 10.3390/gels9040322
Arrioja-Bretón, D., Mani-López, E., Palou, E., et al., 2020. Antimicrobial activity and storage stability of cell-free supernatants from lactic acid bacteria and their applications with fresh beef. Food Control 115: 107286. 10.1016/j.foodcont.2020.107286
Azhdari, S., and Moradi, M., 2022. Application of antimicrobial coating based on carboxymethyl cellulose and natamycin in active packaging of cheese. International Journal of Biological Macromolecules 209: 2042–2049. 10.1016/j.ijbiomac.2022.04.185
Barros, C.P., Guimarães, J.T., Esmerino, E.A., et al., 2020. Paraprobiotics and postbiotics: Concepts and potential applications in dairy products. Current Opinion in Food Science 32: 1–8. 10.1016/j.cofs.2019.12.003
Belhocine, M., Bourzami, R., Dergal, F., et al., 2023. Physical, chemical and antibacterial properties of benzethonium chloride: Experimental and ab-initio analysis. Journal of Molecular Structure 1293: 136299. 10.1016/j.molstruc.2023.136299
Bisson, G., Comuzzi, C., Giordani, E., et al., 2023. An exopolysaccharide from Leuconostoc mesenteroides showing interesting bioactivities versus foodborne microbial targets. Carbohydrate Polymers 301 : 120363. 10.1016/j.carbpol.2022.120363
Chen, D.F., Li, X., Xu, Z., et al., 2010. Hexadecanoic acid from Buzhong Yiqi decoction induced proliferation of bone marrow mesenchymal stem cells. Journal of Medicinal Food 13(4): 967–975. 10.1089/jmf.2009.1293
Conte, A., Scrocco, C., Sinigaglia, M., et al., 2007. Innovative active packaging systems to prolong the shelf life of Mozzarella cheese. Journal of Dairy Science 90(5): 2126–2131.
Costa, M.J., Maciel, L.C., Teixeira, J.A., et al. (2018). Use of edible films and coatings in cheese preservation: Opportunities and challenges. Food Research International 107: 84–92. 10.1016/j.foodres.2018.02.013
d’Errico, G., Aloj, V., Ventorino, V., et al., 2020. Methyl t-butyl ether-degrading bacteria for bioremediation and biocontrol purposes. PLOS ONE 15(2): e0228936. 10.1371/journal.pone.0228936
Deshwal, G.K., Panjagari, N.R., Singh, A.K., et al., 2021. Performance evaluation of a biopolymer-based in-package UV activated colorimetric oxygen indicator with modified atmosphere packaged Mozzarella cheese. Journal of Packaging Technology and Research 5(2): 51–57. 10.1007/s41783-021-00108-1
Divsalar, E., Tajik, H., Moradi, M., et al., 2023. Carbon dot-based antimicrobial photosensitizer: Synthesis, characterization and antimicrobial performance against food borne pathogens. Food Bioscience 56: 103220. 10.1016/j.fbio.2023.103220
Dong, X., Liang, W., Meziani, M.J., et al., 2020. Carbon dots as potent antimicrobial agents. Theranostics 10(2): 671–686. 10.7150/thno.39863
El-Hawary, S.S., El-Tantawi, M.E., Kirollos, F.N., et al., 2018. Chemical composition, in vitro cytotoxic and antimicrobial activities of volatile constituents from Pyrus communis L., and Malus domestica Borkh. fruits cultivated in Egypt. Journal of Essential Oil Bearing Plants 21(6): 1642–1651. 10.1080/0972060X.2018.1553637
Esmaeili Koutamehr, M., Moradi, M., Tajik, H., et al., 2023. Sour whey-derived carbon dots; synthesis, characterization, antioxidant activity and antimicrobial performance on foodborne pathogens. LWT 184: 114978. 10.1016/j.lwt.2023.114978
Esmaeilnejad-Moghadam, B., Rezaei Mokarram, R., Hejazi, M.A., et al., 2019. Low molecular weight dextran production by Leuconostoc mesenteroides strains: Optimization of a new culture medium and the rheological assessments. Bioactive Carbohydrates and Dietary Fibre 18: 10081.
Ezati, P., Priyadarshi, R., and Rhim, J.W., 2022. Prospects of sustainable and renewable source-based carbon quantum dots for food packaging applications. Sustainable Materials and Technologies 33: e00494. 10.1016/j.susmat.2022.e00494
Faccia, M., Gambacorta, G., Pasqualone, A., et al., 2021. Quality characteristics and consumer acceptance of high-moisture Mozzarella obtained from heat-treated goat milk. Foods 10(4): 833. 10.3390/foods10040833
Fu, Y., and Dudley, E.G., 2021. Antimicrobial-coated films as food packaging: A review. Comprehensive Reviews in Food Science and Food Safety 20(4): 3404–3437.
Ghorbani, M., Tajik, H., Moradi, M., et al., 2022. One-pot microbial approach to synthesize carbon dots from baker’s yeast-derived compounds for the preparation of antimicrobial membrane. Journal of Environmental Chemical Engineering 10(3): 107525. 10.1016/j.jece.2022.107525
Gómez-Narváez, F., Medina-Pineda, Y., and Contreras-Calderón, J., 2017. Evaluation of the heat damage of whey and whey proteins using multivariate analysis. Food Research International 102: 768–775. 10.1016/j.foodres.2017.09.074
Gorrasi, G., Bugatti, V., Tammaro, L., et al., 2016. Active coating for storage of Mozzarella cheese packaged under thermal abuse. Food Control 64, 10–16. 10.1016/j.foodcont.2015.12.002
Hebishy, E., Yerlikaya, O., Reen, F.J., et al., 2024. Microbiological aspects and challenges of dairy powders–II: Biofilm/biofouling. International Journal of Dairy Technology 77(3): 691–712. 10.1111/1471-0307.13076
Hosseini, S.A., Abbasi, A., Sabahi, S., et al. 2022. Application of postbiotics produced by lactic acid bacteria in the development of active food packaging. Biointerface Research in Applied Chemistry 12(5): 6164–6183. 10.33263/BRIAC125.61646183
Hosseini, S.H., Farhangfar, A., Moradi, M., et al., 2024. Beyond probiotics: Exploring the potential of postbiotics and parabiotics in veterinary medicine. Research in Veterinary Science 167: 105133. 10.1016/j.rvsc.2023.105133
Jutinico-Shubach, A., Gutiérrez-Cortés, C., and Suarez, H., 2020. Antilisterial activity of chitosan-based edible coating incorporating cell-free supernatant from Pediococcus pentosaceus 147 on the preservation of fresh cheese. Journal of Food Processing and Preservation 44(9): e14715. 10.1111/jfpp.14715
Kubo, I., Fujita, K., and Nihei, K., 2008. Antimicrobial activity of anethole and related compounds from aniseed. Journal of the Science of Food and Agriculture 88(2): 242–247. 10.1002/jsfa.3079
Lacivita, V., Tarantino, F., Molaei, R., et al., 2023. Carbon dots from sour whey to develop a novel antimicrobial packaging for fiordilatte cheese. Food Research International 172: 113159. 10.1016/j.foodres.2023.113159
Law, J.W.F., Ser, H.L., Ab Mutalib, N.S., et al., 2019. Streptomyces monashensis sp. nov., a novel mangrove soil actinobacterium from east Malaysia with antioxidative potential. Scientific Reports 9(1): 3056. 10.1038/s41598-019-39592-6
Lee, C.L., Kim, G.H., and Yoon, K.S., 2021. Effects of combined aerosolization with ultraviolet C light-emitting diode on enterohemorrhagic Escherichia coli and staphylococcus aureus attached to soft fresh produce. Foods 10(8): 1843. 10.3390/foods10081834
Letlole, B.R., Damen, E.P.C.W., and Jansen van Rensburg, C., 2021. The Effect of α-Monolaurin and butyrate supplementation on broiler performance and Gut health in the absence and presence of the antibiotic growth promoter Zinc bacitracin. Antibiotics 10(6): 651. 10.3390/antibiotics10060651
Li, F., Yang, D., and Xu, H. (2019). Non-metal-heteroatom-doped carbon dots: Synthesis and properties. Chemistry—A European Journal 25(5): 1165–1176. 10.1002/chem.201802793
Lingfa, L., and Ankanagari, S., 2023. GC-MS profiling of reproductive stage withania somnifera for antimicrobial and anticancer phytochemicals. Biomedical and Pharmacology Journal 16(1): 197–211. 10.13005/bpj/2601
Lotfy, M.M., Hassan, H.M., Hetta, M.H., et al., 2018. Di-(2-ethylhexyl) Phthalate, a major bioactive metabolite with antimicrobial and cytotoxic activity isolated from River Nile derived fungus Aspergillus awamori. Beni-Suef University Journal of Basic and Applied Sciences 7(3): 263–269. 10.1016/j.bjbas.2018.02.002
Lucera, A., Mastromatteo, M., Conte, A., et al., 2014. Effect of active coating on microbiological and sensory properties of fresh mozzarella cheese. Food Packaging and Shelf Life 1(1): 25–29. 10.1016/j.fpsl.2013.10.002
Luengo, J.M., and Olivera, E.R., 2020. Catabolism of biogenic amines in Pseudomonas species. Environmental Microbiology 22(4): 1174–1192. 10.1111/1462-2920.14912
Mani-López, E., Arrioja-Bretón, D., and López-Malo, A., 2022. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Comprehensive Reviews in Food Science and Food Safety 21(1): 604–641. 10.1111/1541-4337.12872
Marino, M., Dubsky de Wittenau, G., Saccà, E., et al., 2019. Metagenomic profiles of different types of Italian high-moisture Mozzarella cheese. Food Microbiology 79: 123–131. 10.1016/j.fm.2018.12.007
Matsuzaki, C., Nakashima, Y., Endo, I., et al., 2021. Enzymatically synthesized exopolysaccharide of a probiotic strain Leuconostoc mesenteroides NTM048 shows adjuvant activity to promote IgA antibody responses. Gut Microbes 13(1) : 1949097. 10.1080/19490976.2021.1949097
Miao, S., Liang, K., Zhu, J., et al., 2020. Hetero-atom-doped carbon dots: Doping strategies, properties and applications. Nano Today 33: 100879. 10.1016/j.nantod.2020.100879
Mohammadi, R., Moradi, M., Tajik, H., et al., 2022. Potential application of postbiotics metabolites from bioprotective culture to fabricate bacterial nanocellulose based antimicrobial packaging material. International Journal of Biological Macromolecules 220: 528–536. 10.1016/j.ijbiomac.2022.08.108
Moradi, M., Kousheh, S.A., Razavi, R., et al., 2021a. Review of microbiological methods for testing protein and carbohydrate-based antimicrobial food packaging. Trends in Food Science and Technology 111: 595–609. 10.1016/j.tifs.2021.03.007
Moradi, M., Mardani, K., and Tajik, H., 2019. Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. LWT 111: 457–464. 10.1016/j.lwt.2019.05.072
Moradi, M., Molaei, R., and Guimarães, J.T., 2021b. A review on preparation and chemical analysis of postbiotics from lactic acid bacteria. Enzyme and Microbial Technology 143: 109722. 10.1016/j.enzmictec.2020.109722
Moradi, M., Molaei, R., Kousheh, S.A., et al., 2023. Carbon dots synthesized from microorganisms and food by-products: Active and smart food packaging applications. Critical Reviews in Food Science and Nutrition, 63(14): 1943–1959. 10.1080/10408398.2021.2015283
Mukherjee, B., Paul, P., Dutta, L., et al., 2017. Pulmonary administration of biodegradable drug nanocarriers for more efficacious treatment of fungal infections in lungs: Insights based on recent findings. Multifunctional Systems for Combined Delivery, Biosensing and Diagnostics, 1778. Elsevier Inc. 10.1016/b978-0-323-52725-5.00014-9
Muktha, H., Sharath, R., Kottam, N., et al. 2021. Carbon dots—A study of its cytotoxic activity against HepG2 and MCF-7 cell lines. Materials Today: Proceedings 49: 608–612. 10.1016/j.matpr.2021.04.515
Nadar Rajivgandhi, G., Ramachandran, G., Chenthis Kanisha, C., et al., 2020. Anti-biofilm compound of 1, 4-diaza-2, 5-dioxo-3-isobutyl bicyclo[4.3.0]nonane from marine Nocardiopsis sp. DMS 2 (MH900226) against biofilm forming K. pneumoniae. Journal of King Saud University—Science 32(8): 3495–3502. 10.1016/j.jksus.2020.10.012
Nie, X., Jiang, C., Wu, S., et al., 2020. Carbon quantum dots: A bright future as photosensitizers for in vitro antibacterial photodynamic inactivation. Journal of Photochemistry and Photobiology B: Biology 206: 111864. 10.1016/j.jphotobiol.2020.111864
Obis, D., Paris, M., and Ouwehand, A.C., 2019. The safety of lactic acid bacteria for use in foods. Lactic Acid Bacteria Microbiological and Functional Aspects.
Ottonelli Stopiglia, C.D., Mezzomo Collares, F., Aulo Ogliari, F., et al., 2012. Antimicrobial activity of [2-(methacryloyloxy)ethyl]trimethylammonium chloride against Candida spp. Revista Iberoamericana de Micología 29(1): 20–23. 10.1016/j.riam.2011.03.003
Padmavathi, A.R., Abinaya, B., and Pandian, S.K., 2014. Phenol, 2,4-bis(1,1-dimethylethyl) of marine bacterial origin inhibits quorum sensing mediated biofilm formation in the uropathogen Serratia marcescens. Biofouling 30(9): 1111–1122. 10.1080/08927014.2014.972386
Perveen, K., Bukhari, N.A., Al Masoudi, L.M., et al., 2022. Antifungal potential, chemical composition of Chlorella vulgaris and SEM analysis of morphological changes in Fusarium oxysporum. Saudi Journal of Biological Sciences 29(4): 2501–2505. 10.1016/j.sjbs.2021.12.033
Pimentel, T.C., da Cruz, A.G., and Deliza, R., 2016. Sensory evaluation: Sensory rating and scoring methods. Encyclopedia of Food and Health 744–749. 10.1016/B978-0-12-384947-2.00617-6
Raman, J., Kim, J.S., Choi, K.R., et al., 2022. Application of lactic acid bacteria (LAB) in sustainable agriculture: Advantages and limitations. International Journal of Molecular Sciences 23(14): 7784. 10.3390/ijms23147784
Rehman, N.U., Alsabahi, J.N., Alam, T., et al., 2021. Chemical composition and biological activities of essential oil from aerial parts of Frankenia pulverulenta L., and Boerhavia elegans Choisy from northern Oman. Journal of Essential Oil Bearing Plants 24(5): 1180–1191. 10.1080/0972060X.2021.1978870
Rhetso, T., Shubharani, R., Roopa, M.S., et al., 2020. Chemical constituents, antioxidant, and antimicrobial activity of Allium chinense G. Don. Future Journal of Pharmaceutical Sciences 6(1): 1–9. 10.1186/s43094-020-00100-7
Romero, M.P., Alves, F., Stringasci, M.D., et al., 2021. One-pot microwave-assisted synthesis of carbon dots and in vivo and in vitro antimicrobial photodynamic applications. Frontiers in Microbiology 12: 662149. 10.3389/fmicb.2021.662149
Sarajar, B., Alizadeh, A., Moradi, M., et al., 2023. Effects of postbiotics obtained from food probiotic and protective cultures on proliferation and apoptosis in HCT-116 colorectal cancer cells. Applied Food Biotechnology 10(2): 85–101. 10.22037/afb.v10i2.39745
Segat, A., Biasutti, M., Iacumin, L., et al., 2014. Use of ozone in production chain of high moisture Mozzarella cheese. LWT—Food Science and Technology 55(2): 513–520.
Shaaban, M.T., Ghaly, M.F., and Fahmi, S.M., 2021. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. Journal of Basic Microbiology 61(6): 557–568. 10.1002/jobm.202100061
Sharafi, H., Divsalar, E., Rezaei, Z., et al., 2024. The potential of postbiotics as a novel approach in food packaging and biopreservation: A systematic review of the latest developments. Critical Reviews in Food Science and Nutrition 64(33): 2524–12554. 10.1080/10408398.2023.2253909
Sharafi, H., Moradi, M., and Amiri, S., 2022. Application of cheese whey containing postbiotics of Lactobacillus acidophilus LA5 and Bifidobacterium animalis BB12 as a preserving liquid in high-moisture mozzarella. Foods 11(21): 3387. 10.3390/foods11213387
Sivakumar, R., Jebanesan, A., Govindarajan, M., et al., 2011. Larvicidal and repellent activity of tetradecanoic acid against Aedes aegypti (Linn.) and Culex quinquefasciatus (Say.) (Diptera:Culicidae). Asian Pacific Journal of Tropical Medicine 4(9): 706–710. 10.1016/S1995-7645(11)60178-8
Sullivan, D.J., Cruz-Romero, M.C., Hernandez, A.B., et al., 2020. A novel method to deliver natural antimicrobial coating materials to extend the shelf-life of European hake (Merluccius merluccius) fillets. Food Packaging and Shelf Life 25(5): 100522. 10.1016/j.fpsl.2020.100522
Surowiak, A.K., Lochyński, S., and Strub, D.J., 2020. Unsubstituted oximes as potential therapeutic agents. Symmetry 12(4): 575. 10.3390/sym12040575
Teame, T., Wang, A., Xie, M., et al., 2020. Paraprobiotics and postbiotics of probiotic Lactobacilli, their positive effects on the host and action mechanisms: A review. Frontiers in Nutrition 7: 570344. 10.3389/fnut.2020.570344
Torrijos, R., Nazareth, T.M., Calpe, J., et al., 2022. Antifungal activity of natamycin and development of an edible film based on hydroxyethylcellulose to avoid Penicillium spp. growth on low-moisture Mozzarella cheese. LWT 154: 112795. 10.1016/j.lwt.2021.112795
Varghese, M., and Balachandran, M., 2021. Antibacterial efficiency of carbon dots against Gram-positive and Gram-negative bacteria: A review. Journal of Environmental Chemical Engineering 9(6): 106821. 10.1016/j.jece.2021.106821
Verma, S., Bhardwaj, A., Vij, M., et al., 2014. Oleic acid vesicles: A new approach for topical delivery of antifungal agent. Artificial Cells, Nanomedicine, and Biotechnology, 42(2): 95–101. 10.3109/21691401.2013.794351
Wajda-Hermanowicz, K., Pieniążczak, D., Zatajska, A., et al., 2015. A study on the condensation reaction of 4-amino-3,5-dimethyl-1,2,4-triazole with benzaldehydes: Structure and spectroscopic properties of some new stable hemiaminals. Molecules 20(5): 17109–17131. 10.3390/molecules200917109
Yu, X., Zhao, M., Liu, F., et al., 2013. Identification of 2,3-dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-one as a strong antioxidant in glucose–histidine maillard reaction products. Food Research International 51(1): 397–403. 10.1016/j.foodres.2012.12.044
Zhang, J., Liu, S., Wang, X., et al., 2021. Highly efficient Ti3+ self-doped TiO2 co-modified with carbon dots and palladium nanocomposites for disinfection of bacterial and fungi. Journal of Hazardous Materials 413. 10.1016/j.jhazmat.2021.125318
Zhao, C., Wang, X., Wu, L., et al., 2019. Nitrogen-doped carbon quantum dots as an antimicrobial agent against Staphylococcus for the treatment of infected wounds. Colloids and Surfaces B: Biointerfaces 179: 17–27. 10.1016/j.colsurfb.2019.03.042
Zhao, L., Zhang, M., and Wang, H., 2022. Inhibition of the fishy odor from boiled crab meatballs during storage via novel combination of radio frequency and carbon dots. Food Control 136: 108843. 10.1016/j.foodcont.2022.108843
Zhao, L., Zhang, M., Mujumdar, A.S., et al., 2022. Application of carbon dots in food preservation: A critical review for packaging enhancers and food preservatives. Critical Reviews in Food Science and Nutrition 63(24): 6738–6756. 10.1080/10408398.2022.2039896